
U. O. George-Okafor
Department of Applied Microbiology and Brewing, Enugu State, University of Science and Technology, Agbani, Nigeria
F. J.C. Odibo
Department of Applied Microbiology and Brewing, Nnamdi Azikiwe University, Awka, Nigeria
Journal of Biological Sciences
Year: 2011 | Volume: 11 | Issue: 4 | Page No.: 299-306







ABSTRACT
There are limited reliable information on the commercial production and utilization of proteases for detergent and other industrial uses in Nigeria. Hence, the purification and characterization of Bacillus sp. Gs-3 protease for its potential industrial uses were investigated. The dialyzed crude enzyme was purified 17-fold in a two-step procedure involving carboxymethyl sepharose ionic-exchange chromatography and phenyl sepharose 6-fast flow hydrophobic interaction chromatography. The purified enzyme had its optimal activity at pH 9.0 and 90°C and was stable over a pH range 8.0-11.0. It readily hydrolyzed all the tested protein substrates but exhibited highest affinity for gelatin (Km 0.15 mg mL-1). It retained at least 66.2±0.02% of its original activity in the presence of the tested local commercial detergents and removed bloodstains completely. Its activity was significantly (p>0.05) enhanced by Cu2+ ion but strongly inhibited (75.6±0.07%) by Phenyl-methyl Sulfonyl Fluoride (PMSF). Thus, the enzyme demonstrated desirable properties suitable for its biotechnological applications especially in detergent industry.
PDF Abstract XML References Citation
Received: May 21, 2011;
Accepted: September 08, 2011;
Published: September 28, 2011
How to cite this article
U. O. George-Okafor and F. J.C. Odibo, 2011. Purification and Some Properties of Thermo-stable Alkaline Serine Protease from Thermophilic Bacillus sp. Gs-3. Journal of Biological Sciences, 11: 299-306.
DOI: 10.3923/jbs.2011.299.306
URL: https://scialert.net/abstract/?doi=jbs.2011.299.306
DOI: 10.3923/jbs.2011.299.306
URL: https://scialert.net/abstract/?doi=jbs.2011.299.306
INTRODUCTION
Proteases are very important in both physiological and commercial fields. Proteases from microbial sources are preferred to those from plant and animal sources as they possess desirable characteristics required for their biotechnological applications. For instance, protein deposits in contact lenses can easily be removed with protease from Bacillus sp. 158 (Pawar et al., 2009).
Among the various groups of proteases, alkaline proteases are of utmost industrial importance (Gupta et al., 2002). Their utilization for bating, hair removal and leather processing has been reported (Tork et al., 2010; Hameed et al., 1999; Varela et al., 1997). The best known application of alkaline proteases is their use in household detergents (Ito et al., 1998). They have been successfully incorporated as additives in detergent formulations for removal of protein substances (Kumar et al., 2008). Most are known to have long term stability in detergent products and are energy cost efficient as they lower the washing temperature and thus obtain quicker and more reliable products (Gupta et al., 2002). Their stability in a wide pH range not-with-standing, they may be found to be thermo-labile especially when produced from mesophilic organisms (Griffin et al., 1992). When thermo-stable, they tend to offer additional advantage as they are more rigid than thermo-liable enzymes and more resistant to thermal and chemical denature. Their applications also result in faster reaction rates, increase in solubility of non gaseous reactants and reduced incidence of microbial contamination (Zeikus et al., 1998).On a commercial scale, alkaline proteases from Bacillus sp. have been highly utilized (Gupta et al., 2002); for instance, Alcalase and Esperase from Bacillus licheniformis and Savinase from alkaliphilic strain of Bacillus amyloliquefaciens are produced by Novo industry in Denmark (Outtrup and Boyce, 1990). Unfortunately, the commercial production of proteases for detergent use is not widespread in developing countries like Nigeria that requires such for easy washings of difficult household and hospital clothes that are stained by blood and other protein materials. Thus, based on good properties of thermo-stable alkaline proteases, the study was aimed at purifying and characterizing a thermo-stable alkaline serine protease from a newly isolated Bacillus sp. GS-3 with the view of ascertaining its suitability for biotechnological exploitation especially in detergent industry.
MATERIALS AND METHODS
Collection of materials: The resins and surfactants used for the study were kindly provided by Prof F.J.C. Odibo while the rest of the chemicals and reagents which were of analytical grades were purchased from the open market. The organism (Bacillus sp. Gs-3) utilized for the protease production was obtained from the local stock culture collection center of the Biotechnology unit of Nnamdi Azikiwe University (NAU), Awka, Nigeria.
Study period: The study was carried out from June, 2004 to January, 2007 at the Departmental laboratory and Prof. Odibo’s personal laboratory linked to that of the Department.
Preparation of the organism: The stock organism was activated by culturing in Nutrient broth at 50°C and 180 rev min-1 for 18-24 h using shake-incubator. Then, 5 mL of the growth medium representing 5% of the microbial growth served as the inoculum.
Protease production: The 72 h-fermentation at 50°C was carried out with 5% inoculum in a medium containing (w/v): sucrose (1%), peptone (1%), yeast extract (0.5%), K2HPO4 (0.5%), MgSO4.7H2O (0.02%) and Na2CO3 (2%). The fermentation broth was centrifuged (4°C; 8000 revmin-1) for 10 min and the recovered clear supernatant was assayed for protease activity.
Protease assay: The protease activity was determined by incubating equal volumes of the enzyme and 1% buffered casein (pH 8.0) at 50°C for 30 min. The buffered casein was prepared with 0.2 M phosphate buffer. The reaction was terminated by the addition of 2 mL of 5% Trichloroacetic Acid (TCA). The reaction mixture was centrifuged and the supernatant measured for protease activity by the method of Lowry et al. (1951) as stipulated by Upton and Forgarty (1977). One unit (U) of the protease activity was defined as the amount of protease that released 1 mg of tyrosine from casein per min under the applied condition.
Protein estimation: The protein concentration of the enzyme was measured by the method of Bradford (1976) using Bradford reagent (Coomassie Brilliant Blue G-250) and Bovine Serum Albumin (BSA) as the standard at a wavelength of 595 nm.
Purification of the protease: The crude enzyme (500 mL) was first centrifuged 10-folds by dialyzing overnight against 5 M sucrose solution. The two purification steps involving Carboxymethyl (CM) Sepharose and Phenyl sepharose-6-fast flow high-sub (Pharmacia products, USA) were performed at 4°C.
The dialyzed enzyme (50 mL) was applied to CM Sepharose (2.0x14.5 cm) previously equilibrated with 0.2 M phosphate buffer (pH 8.0). The protein elution was performed by a linear gradient of NaCl concentration of 0-3 in 0.2 M phosphate buffer (pH 8.0) at a flow rate of 60 mL h-1. Each 10 mL fraction collected was assayed for both protease activity and protein content. Thereafter, active fractions with protease activity were pooled and dialyzed as earlier described.
The resulting concentrated enzyme (20 mL) was made hydrophobic by the addition of 3.5 g of 3 M NaCl and the pre-washed phenyl sepharose-6-fast flow high-sub was also made hydrophobic by washing with 3 M NaCl solution in 0.2 M phosphate buffer (pH 8.0). Then, the enzyme was eluted with 150 mL of varying concentrations (from 3 to 1 M) of NaCl solution, followed by 300 mL of 0.2 M phosphate buffer (pH 8.0) at a flow rate of 86 mL h-1. Protein content and protease activity of each 10 mL-fraction were determined as earlier stipulated. The active fractions eluted with 3 M NaCl solution were pooled and first dialyzed with 0.2 M phosphate buffer (pH 8.0) to remove the effect of NaCl before dialyzing with 5 M sucrose solution.
Determination of pH activity and stability: The effect of pH on the enzyme activity was measured according to the standard method except that the reaction was carried out over a pH range of 3.0-11.0. Three buffer systems used were 0.1 M citrate buffer for pH 3.0-6.0, 0.2 M phosphate buffer for pH 7.0-8.0 and 0.2 M phosphate-NaOH buffer for pH 9.0-11.0. The pH stability of the enzyme was estimated by pre-incubating with the same buffer systems (pH 3.0-11.0) for 31/4 h at 30°C and the residual activity measured as earlier described.
Temperature activity and stability profile: This was determined by incubating the reaction mixture at varying temperatures of 40-100°C for 30 min and the stability measured by pre-incubating the enzyme (1 mL) for 30 min at 40-100°C with prompt cooling on ice before the addition of equal volumes of 1% casein (pH 9.0). The thermal stability (half-life) was also examined at varying temperatures of 50-100°C for 60 min at every 10 min interval (Odibo and Ulbrich-Hofmann, 2001). Their residual activities were measured as earlier described.
Effect of inhibitors and metal ions: The effects were determined by pre-incubating equal volumes of the enzyme with each buffered inhibitor or ion (5 mM; pH 9.0) for 30 min at 30°C. The divalent cations used were Cu2+, CO2+, Pb2+, Mg2+, Zn2+, Fe2+, Ca2+, Al2+, Hg2+ and St2+ while the modulators were Phenylmethyl Sulphonylfluoride (PMSF), Iodoacetate (IAA), N-bromosuccinamide (N-BS) and Ethylene bis tetra-acetic acid (Oxyethylene Nitrilo) (sigma, USA). The residual activities were assayed at 90°C and the results obtained were compared with the enzyme-free inhibitor and metal-ion controls. The stimulatory effect of the Cu2+ ion on the thermal stability of the enzyme was evaluated at 50-100°C by the standard method as earlier described and the result compared with that of enzyme-free Cu2+ which served as the control.
Substrate specificity and enzyme kinetics: The enzyme activity was tested against 1% substrate (casein, egg albumin, BSA, gelatin) in 0.2 M phosphate buffer (pH 9.0). Their various concentrations (0-1.0 mg mL-1), were incubated with equal amounts of the enzyme and the activity assayed. The kinetic constants (Km and Vmax) of the enzyme were estimated by double reciprocal plots of the data as described by Lineweaver and Burk (1934).
Detergent stability and cleansing effect: Sodium Dodecyl Sulphate (SDS), Sodium deoxycholate and the local enzyme-free detergents (Klin, Key, Jumbo and GSM) purchased from Nigerian markets were diluted with distilled water to a concentration of 10 mg mL-1. Then, the method of Adinarayana et al. (2003) was applied for the assay at 90°C for 30 min. The residual activity was measured by the standard method and then compared with the control (without the detergent). The cleansing potential of the enzyme as a detergent additive was examined on 48 h-blood stained pieces of white cloth as described by Adinarayana et al. (2003). The treated blood-stained clothes were, respectively incubated at 90°C for 10 min. Thereafter, the clothes were rinsed with clean water and air-dried. Visual examination of the dried clothes served as the measure of the enzyme cleansing ability (Durham et al., 1987; Showell, 1999; Wolff et al., 1996). The untreated pieces of cloth served as the control.
Statistical analysis: The relative activity of each variable was expressed in relation to the control which was taken as 100%. Then, the software application (Statistical Package for Social Sciences (SPSS), Version 16.0, was utilized to analyze the data using paired t-test. A calculated p-value >0.05 was considered to be statistically significant.
RESULTS AND DISCUSSION
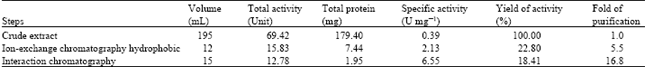
Enzyme purification: The results of the enzyme purification are summarized in Table 1. A two-step process resulted in a 17-fold purification with a yield of 18.4% and a specific activity of 6.55 U mg-1 proteins. On phenyl sepharose 6-fast flow (high sub), there was a single sharp peak coinciding with protease activity of 30.1 Um L-1 from fraction 7 (Fig. 1). The single sharp peak of protease activity by phenyl sepharose 6-fast, suggests that the protease may be the major casein-degrading enzyme produced by the Bacillus sp.
pH activity and stability profile: The activity and stability optima of the enzyme were at pH 9.0 (Fig. 2). The pH activity profile is the same with the protease of Bacillus licheniformis which also demonstrated maximum activity at pH 9.0 (Al-Shehri and Yasser, 2004).
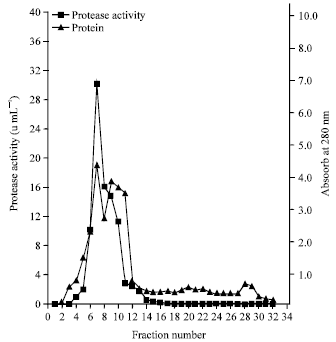 | |
| Fig. 1: | Elution profile of alkaline protease of Bacillus sp. GS-3 on phenyl sepharose 6-fastflow High-sub |
| Table 1: | Purification summary of alkaline protease of Bacillus sp. GS-3 |
 | |
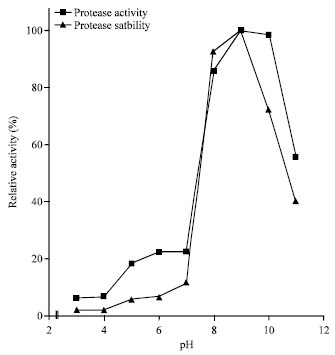 | |
| Fig. 2: | Effect of pH on the activity and stability of alkaline protease from Bacillus sp. GS-3 |
The results closely resemble thermo-stable serine alkaline proteases from Bacillus subtilis PE-11; Thermoactinomyces sp. strain H5682 and Streptomyces sp. YSA-130 that had their pH optima between 10.0 and 11.5 (Adinarayana et al., 2003; Tsuchiya et al., 1991; Yum et al., 1994). Stability over a broad pH (8.0-11.0) and low activity (<28%) at pH 3.0-7.0 are indications that the enzyme is alkaline.
Temperature activity and stability profile: The protease demonstrated optimal activity at 90°C (Fig. 3). The optimal activity at 90°C was higher than the thermo-stable proteases from Bacillus brevis SSAI, Natrialba magadii, Bacillus subtilis PE-11 and Thermoactinomyces thalpophilus which had their optimal activities between 60-70°C (Aftab et al., 2006; Adinarayana et al., 2003; Gimenez et al., 2000; Odibo and Obi, 1988).
The thermal stability profile (Fig. 3) showed that the enzyme was optimally stable at 70°C but the activity gradually decreased with time (Fig. 4). However, the protease still had a half-life (t1/2) of a 50 min at 100°C and this result is higher than t1/2 <50 and t1/2 of 3.4 min at 50°C demonstrated, respectively by Bacillus subtilis PE-11 protease and Subtilisin Carlsberg (Adinarayana et al., 2003; Durham, 1987). The observed stability profile indicates the thermo-stability property of the enzyme which is an advantage for use in industrial fermentation processes.
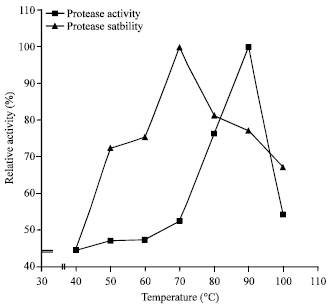 | |
| Fig. 3: | Effect of temperature on the activity and stability of alkaline protease from Bacillus sp. GS-3 |
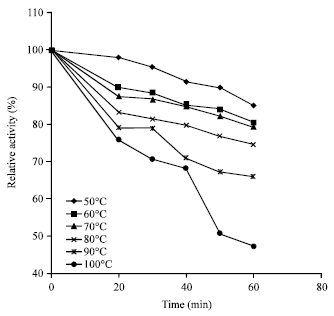 | |
| Fig. 4: | Thermal stability of alkaline protease from Bacillus sp. GS-3 at temperature range of 50-100°C |
Effect of metal ions and modulators: Among all the tested metal ions, only Cu2+ enhanced (226.7%) the activity of the enzyme (Table 2). Thermal stabilization of the protease by Cu2+ was achieved most at 50°C but up to 85% activity was retained at 100°C (Fig. 5). The demonstration of Cu2+ as a good stimulant to the study enzyme activity is in line with the report from Ogbonna et al. (2003) but at variance with the findings of Rahman et al. (1994) where Cu2+ reduced the activity of Bacillus stearothermophilus F1 protease.
| Table 2: | The influence of metal ion on the activity of alkaline protease of Bacillus sp. GS-3 |
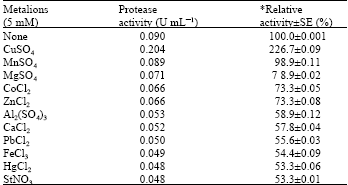 | |
| *Relative activity was expressed in comparison with the activity of the enzyme free-metal ion which was taken as 100%. SE: Standard error | |
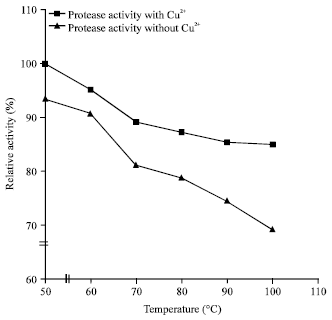 | |
| Fig. 5: | Temperature stability profiles of the protease of Bacillus sp. GS-3 in the presence and absence of Cu2+ |
This result suggests that Cu2+ apparently protected the enzyme against thermal denature and played a vital role in maintaining the active conformation of the enzyme at high temperature (Donaghy and Mckay, 1993).
All the tested modulators had some inhibitory effects on the enzyme activity but PMSF mostly inhibited (24.4%) the activity (Table 3). This gives the evidence that the enzyme is a serine protease since PMSF sulphonates the essential serine residue in the active site of the protease and this has been reported to result in extensive loss of enzyme activity (Gold and Fahrney, 1964). The inhibition of the enzyme by PMSF is comparable to earlier reports on the serine protease inhibition by PMSF (Aftab et al., 2006; Gimenez et al., 2000; Tsuchida et al., 1986; Yamagata and Ichishima, 1989). The inhibitory effect (41%) by the EDTA also suggests the enzyme to be a metallo-protease but not a cysteine (thiol) protease.
Enzyme hydrolytic activity: The protease hydrolyzed both plant and animal protein substrates.
| Table 3: | Effect of modulators on alkaline protease of Bacillus sp. GS-3 |
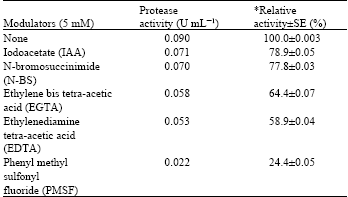 | |
| *Relative activity was expressed in comparison with the activity of the enzyme free-modulators and was taken as 100% | |
| Table 4: | Relative rates of substrate hydrolysis by alkaline protease of Bacillus sp. GS-3 |
 | |
The hydrolytic activity was more on gelatin (100%) than on casein (93.8%), BSA (71.8%) and egg albumin (58.3%) (Table 4). The greater affinity of the enzyme for gelatin is comparable to protease from Halobacterium sp. AFI which exhibited highest proteolytic activity on gelatin than casein and bovine serum albumin (Habib et al., 2011). The hydrolytic activity of this enzyme is encouraging because some enzymes such as trypsin and neutrase exhibited limited hydrolysis on certain protein products (Hou and Zhao, 2011). The enzyme kinetic studies showed low Km values, ranging from Km 0.15 to 0.33 mg mL-1 (Fig. 6) and this is a confirmation that the enzyme had broad substrate specificity (Plummer, 1978); hence, its potential as a hydrolytic agent in industrial fermentations. Judging from the Km values obtained, this enzyme is characteristically very similar to the protease from sorghum malt variety KSV-11 (Ogbonna et al., 2003) but at variance with some other reports (Khan et al., 2000, 2008; Ogbonna et al., 2004).
Detergent stability and cleansing effect: The result on the enzyme stability with the local laundry detergents (GSM, JUMBO, KEY and KLIN), shows that the protease retained 90.8% of its initial activity with GSM detergent, followed by Jumbo (78.5%), the least stability (66.2%) was with Klin (Table 5). This result implies that the enzyme exhibited good stability and compatibility in the presence of local laundry detergents as over 65% of its initial activity at pH 9.0 for 1h was retained. However, the 65% retention was lower than the 90% obtained from Spilosoma obliqua protease (Anwar and Saleemuddin, 2000).
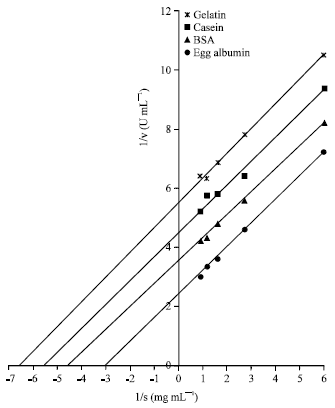 | |
| Fig. 6: | Lineweaver-burk plot for the determination of km of alkaline protease from Bacillus sp. GS-3 |
| Table 5: | Alkaline protease stability against different local laundry detergents |
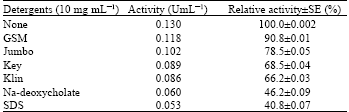 | |
The difference could be attributed to the detergent formulations used since the test detergents for Spilosoma obliqua protease were not enzyme free as those utilized in the study.
The enzyme was labile in the presence of Sodium deoxycholate and SDS as 53.8 to 59.2% activity was lost. The lost activity could be explained from the fact that the bound SDS/Sodium deoxycholate contributes to a large net negative charge to the enzyme, thereby rendering its intrinsic charge insignificant and consequently altering the enzyme conformation. The negative effects of SDS and Sodium deoxycholate on protease activity have previously been reported by Anwar and Saleemuddin (2000) and Gimenez et al. (2000).
The blood cleansing ability of the test detergent improved more with the addition of the purified protease than without (Fig. 7). This suggests strongly that the enzyme has all the potentials to be used as laundry detergent additive which will improve the performance of heavy-duty laundry detergents.
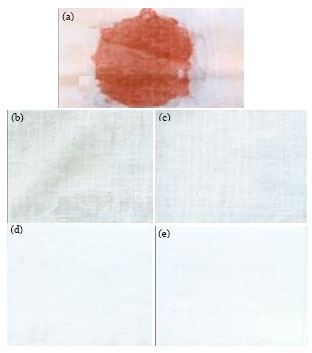 | |
| Fig. 7(a-e): | (a) 48 h-blood stained white cloth (unwashed). (b) Washed blood stained white cloth without detergent and protease. (c) Washed blood stained white cloth with only detergent. (d) Washed blood stained white cloth with detergent and enzyme. (e) Unstained white cloth (control) |
CONCLUSION
The purified protease from thermophilic Bacillus sp. GS-3 demonstrated useful properties such as high optimum temperature, increased thermal stability at alkaline pH, broad substrate specificity and excellent compatibility with detergents. These properties call for its utilization in industries especially in the detergent industry where it can be fully exploited commercially like Alcalase and Esperase from Bacillus licheniformis produced by Novo industry, Denmark under the trade name Biotex.
The immobilization and characterization of the enzyme will be determined since immobilization will facilitate its re-use at higher concentration in bioreactors.
ACKNOWLEDGMENTS
The authors wish to thank the laboratory workers of Biotechnology center of the university who assisted in providing and washing of the used glass-wares. The Director and the Head of the Department who provided stand-by generator in times of power failure.
REFERENCES
- Adinarayana, K., P. Ellaiah and D.S. Prasad, 2003. Purification and partial characterization of thermostable serine alkaline protease from a newly isolated Bacillus subtilis PE-11. AAPS Pharm. Sci. Technol., 4: 1-9.
Direct Link - Aftab, S., S. Ahmed, S. Saeed and S.A. Rasool, 2006. Screening, isolation and characterization of alkaline protease producing bacteria from soil. Pak. J. Biol. Sci., 9: 2122-2126.
CrossRefDirect Link - Habib, S.M.A., A.N.M. Fakhruddin, S. Begum and M.M. Ahmed, 2011. Production and characterization of thermo-alkaline extracellular protease from Halobacterium sp. AF1. Asian J. Biotechnol., 3: 345-356.
CrossRefDirect Link - Al-Shehri, M.A. and M.S. Yasser, 2004. Production and some properties of protease produced by Bacillus licheniformis isolated from Tihamet Aseer, Saudi Arabia. Pak. J. Biol. Sci., 7: 1631-1635.
CrossRefDirect Link - Anwar, A. and M. Saleemuddin, 2000. Alkaline protease from Spilosoma obliqua: Potential applications in bioformulation. Biotechnol. Appied Biochem., 31: 85-89.
PubMed - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Donaghy, J.A. and A.M. McKay, 1993. Production and properties of an alkaline protease by Aureobasidium pullulans. J. Applied Bacteriol., 74: 662-666.
CrossRefDirect Link - Durham, D.R., 1987. Utility of subtilisin GX as a detergent additive. J. Appllied Bacteriol., 63: 381-386.
CrossRefDirect Link - Durham, D.R., D.B. Stewart and E.J. Stellwag, 1987. Novel alkaline and heat stable proteases from alkalophilic Bacillus species strain GX 6638. J. Bacteriol., 169: 2762-2768.
Direct Link - Gimenez, M.I., C.A. Studdert, J.J. Sanchez and R.E. De Castro, 2000. Extracellular protease of Natrialba Magadii purification and biochemical characterization. Extremophiles, 4: 181-188.
CrossRefDirect Link - Griffin, H.L., R.V. Greene and M.A. Cotta, 1992. Isolation and characterization of an alkaline protease from the marine shipworm bacterium. Curr. Microbiol., 24: 111-117.
CrossRefDirect Link - Hameed, A., T. Keshavarz and C.S. Evans, 1999. Effect of dissolved oxygen tension and pH on the production of extracellular protease from a new isolate of Bacillus subtilis K2, for use in leather processing. J. Chem. Technol. Biotechnol., 74: 5-8.
Direct Link - Ito, S., T. Kobayashi, K. Ara, K. Ozaki, S. Kawai and Y. Hatada, 1998. Alkaline detergent enzymes from alkaliphiles: Enzymatic properties, genetics and structures. Extremophiles, 2: 185-190.
PubMed - Khan, H., M. Subhan, S. Mehmood, M.F. Durrani, S. Abbas and S. Khan, 2008. Purification and characterization of serine protease from seeds of Holarrhena antidysenterica. Biotechnology, 7: 94-99.
CrossRefDirect Link - Khan, A.R., S. Nirasawa, S. Kaneko, T. Shimonishi and K. Hayashi, 2000. Characterization of a solvent resistant and thermo-stable amino-peptidase from the hyperthermophilic bacterium, Aquifex aeolicus. Enzy. Microb. Technol., 27: 83-88.
CrossRefDirect Link - Kumar, D., Savitri, N. Thakur, R. Verma and T.C. Bhalla, 2008. Microbial proteases and application as laundry detergent additive. Res. J. Microbiol., 3: 661-672.
CrossRefDirect Link - Lineweaver, H. and D. Burk, 1934. The determination of enzyme dissociation constants. J. Am. Chem. Soc., 56: 658-666.
CrossRefDirect Link - Odibo, F.J.C. and S.K.C. Obi, 1988. Purification and some properties of a thermostable protease of Thermoactinomyces thalpophilus. World J. Microbiol. Biotechnol., 4: 327-332.
CrossRefDirect Link - Odibo, F.J.C. and R. Ulbrich-Hofmann, 2001. Thermostable &alpha-amylase and glucoamylase from Thermomyces lanuginosus F1. Acta Biotechnol., 21: 141-153.
CrossRef - Ogbonna, A.C., S.K.C. Obi, B.N. Okolo and F.J.C. Odibo, 2003. Purification and some properties of a protease from sorghum malt variety KSV8-11. J. Inst. Brew., 109: 179-186.
Direct Link - Ogbonna, A.C., S.K.C. Obi, B.N. Okolo and F.J.C. Odibo, 2004. Purification and some properties of a cysteine proteinase from sorghum malt variety SK5912. J. Sci. Food Agric., 84: 113-120.
CrossRefDirect Link - Pawar, R., V. Zambare, S. Barve and G. Paratkar, 2009. Application of protease isolated from Bacillus sp. 158 in enzymatic cleansing of contact lenses. Biotechnol., 8: 276-280.
CrossRefDirect Link - Rahman, R.N.Z.A., C.N. Razak, K. Ampon, M. Basri, W.M. Zin, W. Yunus and A.B. Salleh, 1994. Purification and characterization of a heat-stable alkaline protease from Bacillus stearothermophilus F1. Applied Microbiol. Biotechnol., 40: 822-827.
CrossRefDirect Link - Tork, S., M.M. Aly and L. Nawar, 2010. Biochemical and molecular characterization of a new local keratinase producing Pseudomomanas sp., MS21. Asian J. Biotechnol., 2: 1-13.
CrossRefDirect Link - Tsuchida, O., Y. Yamagata, T. Ishizuka, T. Arai, J.I. Yamada, M. Takeuchi and E. Ichishima, 1986. An alkaline proteinase of an alkalophilic Bacillus sp. Curr. Microbiol., 14: 7-12.
CrossRefDirect Link - Upton, M.E. and W.M. Fogarty, 1977. Production and purification of thermostable amylase and protease of Thermomonospora viridis. Applied Microbiol., 33: 59-64.
Direct Link - Varela, H., M.D. Ferrari, L. Belobradjie, A. Vazquez and M.L. Loperena, 1997. Skin unhairing proteases of Bacillus subtilis: Production and partial characterization. Biotechnol. Lett., 19: 755-758.
CrossRefDirect Link - Hou, Y. and X.H. Zhao, 2011. Limited hydrolysis of two soybean protein products with trypsin or neutrase and the impacts on their solubility, gelation and fat absorption capacity. Biotechnology, 10: 190-196.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Gold, A.M. and D. Fahrney, 1964. Sulfonyl fluorides as inhibitors of esterases. II. Formation and reactions of phenylmethane sulfonyl α-chymotrypsin. Biochemistry, 3: 783-791.
PubMed - Gupta, R., Q. Beg and P. Lorenz, 2002. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol., 59: 15-32.
CrossRefPubMedDirect Link