
S.M. Ahsan Habib
Department of Environmental Sciences, Jahangirnagar University, Dhaka-1342, Bangladesh
LiveDNA: 880.22510
A.N.M. Fakhruddin
Department of Environmental Sciences, Jahangirnagar University, Dhaka-1342, Bangladesh
LiveDNA: 880.3906
ORCID: 0000-0002-6058-6950
Shamima Begum
Department of Microbiology and Biotechnology, Jagannath University, Dhaka-1100, Bangladesh
Monzur Morshed Ahmed
Institute of Food Science and Technology, Bangladesh Council of Scientific and Industrial Research, Dhaka-1205, Bangladesh
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 4 | Page No.: 345-356







ABSTRACT
Optimum incubation period, temperature and pH for production of extracellular alkaline protease production by novel bacterium Halobacterium sp. AF1 were investigated. The protease were also characterized for its stability on various environmental conditions. Extracellular thermo-alkaline activity was determined by measuring proteolytic activity of the culture supernatants and stability of the protease was determined by incubating protease under different environmental conditions. The optimum production of AF1 protease was achieved within 72 h of culture at 40°C with the pH 8. The activity of the protease produced by this organism was stable up to 91% at 70°C and 83% at pH 9.5. AF1 protease was not only stable in the presence of organic solvent (25% v/v) but also exhibit a higher activity for acetone and n-hexane than in the absence of solvents. These results suggested that the protease produced by the organism was thermo-stable organic solvent tolerant extracellular alkaline protease. The remarkable stability was found for 5 mM Ca+2, Na+, Hg+2 and Fe+3 as they had been showed marginal effect on proteolytic activity of isolated protease. The AF1 protease was retained their activity up to 57-67% for 1.5% sodium dodecyl sulfate and H2O2. Higher proteolytic activity was found for gelatin (1% w/v) than that of casein or bovine serum albumin. The AF1 protease exhibited excellent stability towards temperature, pH, organic solvents and metal ions. These properties make it suitable to be used in tannery industrial applications as well as detergent additive which reduce the industrial use of chemicals wastes in the environment.
PDF Abstract XML References Citation
Received: February 24, 2011;
Accepted: April 25, 2011;
Published: June 15, 2011
How to cite this article
S.M. Ahsan Habib, A.N.M. Fakhruddin, Shamima Begum and Monzur Morshed Ahmed, 2011. Production and Characterization of Thermo-alkaline Extracellular Protease from Halobacterium sp. AF1. Asian Journal of Biotechnology, 3: 345-356.
DOI: 10.3923/ajbkr.2011.345.356
URL: https://scialert.net/abstract/?doi=ajbkr.2011.345.356
DOI: 10.3923/ajbkr.2011.345.356
URL: https://scialert.net/abstract/?doi=ajbkr.2011.345.356
INTRODUCTION
There are nearly 300 tanneries in Hazaribagh, Dhaka. Daily discharge of waste from these tanneries is about 18,000 litres of liquid waste, 115 tones solid wastes during off-peak time. According to a recent estimate, about 60,000 tons of raw hides and skins are processed in these tanneries every year, a process which releases nearly 95,000 litres of untreated effluents into the open environment daily, in addition to contamination of the water of the river Buriganga with high chemical and biological oxygen demand (Rusal et al., 2006).
Tannery is a red category industry (Farooque and Hasan, 2004). Leather industry contributes to one of the major industrial pollution problems facing the country and the pollution causing chemicals, viz. lime, sodium sulphide, salt, solvents, etc. arise mainly from the pre-tanning processes of leather processing (UNEP, 1991). An important enzyme used in pre-tanning processes belongs to the group of proteolytic enzymes and proteases.
Proteases constitute one of the most important groups of industrial enzymes accounting for more than 60% of the total enzyme sales (Gupta et al., 2002, 2005). The applications of proteases include in the leather, detergent, food, meat tenderization industries. Proteases are also important tools in studying the structure of proteins and peptides. Besides that, they are also used in pharmaceuticals, medical diagnosis, lens cleansing, decomposition of gelatin on X-ray films as well as in textiles as a non-hazardous and bioalternative (Joo et al., 2002; Patel et al., 2005; Tari et al., 2006; Pawar et al., 2009). Proteolysis enzymes are derivative enzymes which catalyze the cleavage of peptide bonds in other proteins. For environmental reasons, the mistreatment of leather using an enzymatic approach is preferable as it offers several advantages, e.g., easy control, speed and waste reduction, thus being eco-friendly (Andersen, 1998). As protease represents one of the major groups of industrial enzymes, it is worthwhile to screen microbes from new habitats for proteases with novel properties to meet the needs of rapidly growing industries (Kumar et al., 2008). Aims of the present study were:
| • | Optimization of environmental conditions for production of extracellular alkaline protease by a novel bacterium Halobacterium sp. AF1 |
| • | Characterization of the protease produced by the organism |
MATERIALS AND METHODS
Materials: Bacterial isolate was collected from Industrial Microbiology section, Institute of Food Science and Technology, BCSIR, Dhaka on 10th January, 2010 (Habib., 2010). Chemicals were obtained from different sources MERCK, BDH, INVITROGEN and OXOID.
Cultural condition for the Production of alkaline protease: A loopful of culture from agar plate was inoculated into 100 mL-glass tube containing 10 mL of alkaline protease production medium and incubated overnight at 180 rpm and 50°C. Five millilitre of this culture was then inoculated into 250 mL capacity Erlenmeyer flask containing 95 mL of the same medium and incubated at 50°C in an orbital shaker for 72 h. Cells and insoluble materials were removed by centrifugation at 10000 g for 10 min at 4°C and the cell free supernatant was filtered through a 0.2 μm pore-size membrane filter and was used as the source of crude enzyme.
Alkaline protease assay: Protease activity was determined by a slightly modified method of Yang and Huang (1994). One protease unit was defined as the amount of enzyme that releases 1 μg of tyrosine per mL per minute under the defined assay conditions.
Determination of bacterial growth: For the determination of bacterial growth sample was centrifuged at 10000 g for 10 min at 4°C and the pellet was washed with sterilized normal saline three times to remove the suspended particles. Finally, absorbance was taken at 660 nm.
Total protein determination: Protein content of the enzyme solution was measured by the method of Lowry et al. (1951) using Bovine Serum Albumin (BSA) as a standard.
Effect of temperature on the activity of crude alkaline protease: The influence of temperature on the catalytic activity of the crude alkaline protease was determined by incubating enzyme at 30, 40, 50, 60 and 70°C. Thereafter protease activity was measured for control and incubated samples.
Effect of pH on the activity of the crude alkaline protease: The influence of pH on the alkaline protease activity was determined by measuring the enzyme activity after incubating enzyme sample at 40°C for 30 min at varying pH values ranging from 6.5 to 9.5 using different suitable buffers, 50 mM sodium acetate (pH 6.5), 50 mM sodium phosphate (pH 7.5), 50 mM Tris-HCl (pH 8.5), 50 mM glycine-NaOH (pH 9.5). Thereafter protease activity was measured for control and incubated samples.
Organic solvent-stability of crude alkaline protease: Three milliliter of crude protease was incubated with 1.0 mL of organic solvent at 40°C with a constant shaking at 150 rpm for 30 min. For control, the solvent was replaced by distilled water. The relative activity which remained after 30 min of incubation in 25% (v/v) of organic solvent was measured. The organic solvents chosen in this study were toluene, acetone, butanol and n-hexane. Thereafter protease activity was measured for control and incubated samples.
Effect of various metal ions on proteolytic activity: The effects of different metal ions (e.g., Ca2+, Na+, Hg+2, Fe+3) at 5 mM concentration on protease activity was determined by incubating the enzyme with different metal salts at 40°C for 30 min and thereafter the relative activities were determined for control and incubated samples.
Effect of surfactants and oxidants on protease activity: The compatibility of protease with surfactants and oxidants was studied in the presence of 0.5, 1.0 and 1.5% SDS (Sodium dodecyl sulphate) as surfactants and 0.5, 1.0 and 1.5% of H2O2 as oxidants. The enzyme sample was incubated at 40°C with surfactants and oxidants for 30 min and thereafter the proteolytic activity was measured as standard assay condition. The enzyme activity of a control sample (without surfactants and oxidants) was considered as 100%.
Effect of protease inhibitors and chelators on enzyme activity: The effect of 5 mM various protease inhibitors such as cysteine-inhibitor β-mercaptoethanol [β-ME] and a chelator of divalent cations Ethylene Diamine Tetra Acetic Acid (EDTA) (metalloprotease inhibitor) were determined by preincubation with the enzyme solution for 30 min at 40°C. The enzyme activity of a without inhibitors was considered as 100%.
Hydrolysis of protein substrates: Protease activity with various protein substrates including bovine serum albumin, casein and gelatin was assayed. 1% of protein substrate was incubated with protease for 30 min at 40°C. The reaction was stopped by adding 10% of trichloroacetic acid and allowed to stand at room temperature for 10 min. The undigested protein was removed by centrifugation and peptides released were assayed. The specific protease activity towards casein was taken as a control.
RESULTS
Production of protease
Optimization of incubation period for protease production: Halobacterium sp. AF1 was incubated in protease producing broth, proteolytic activity, biomass (OD at 660 nm) and total protein concentration in media was estimated at different time intervals and results are presented in Fig. 1. Protease activity increase with incubation period up to 72 h and highest protease activity was found in 72 h, corresponding growth was also higher than that of other times. Media proteins decreased with increase in biomass. After 72 h protease activity and biomass reduced with time.
The effect of temperature on the production of thermo-stable extracellular alkaline protease by Halobacterium sp. AF1: The optimum temperature for the production of protease was investigated for 30, 40 and 50°C. From Fig. 2, it can be concluded that incubation period of 72 h was optimum for the production of alkaline protease by Halobacterium sp. AF1. In shaken culture, 40°C was found to be optimum temperature for both protease production and bacterial growth. Incubation at 30 and 50°C was found to the lower production of protease. Bacterial growth was highest at 72 h and then decreased. The total media protein was decreased with incubation time.
The effect of pH on the production of thermo-stable extracellular alkaline protease: Extra-cellular protease was detected over a pH range (pH 6 to pH 9) with the optimum production of protease and bacterial growth exhibited at pH 8. From the analysis of Fig. 3, it can be concluded that 72 h was the optimum incubation time for the growth and protease production for Halobacterium sp. AF1. The bacterial growth and production of protease in an alkaline medium at pH 8 were higher as compared to that in an acidic medium at pH 6.
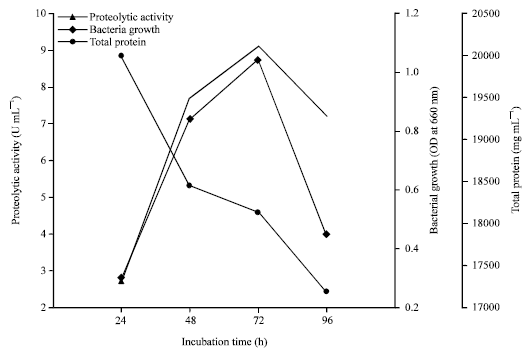 | |
| Fig. 1: | Proteolytic activity, bacterial growth and total protein after different incubation period at pH 8 for 30°C |
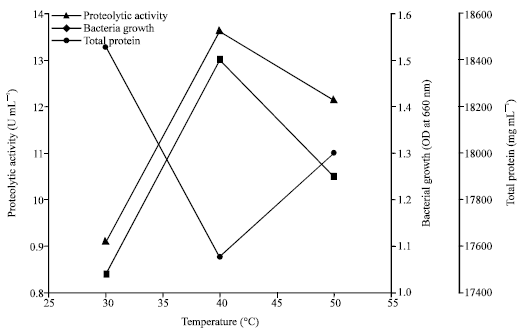 | |
| Fig. 2: | Proteolytic activity, bacterial growth and total protein after incubation period of 72 h pH 8 for different temperature |
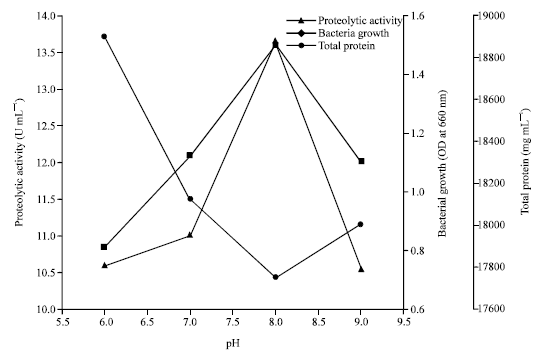 | |
| Fig. 3: | Proteolytic activity, bacterial growth and total protein after incubation period of 72 h at 40°C for different pH |
Characterization of the AF1 protease
Effects of temperature and pH on the stability of AF1 protease: The stability of produced protease was examined by incubating it in 30 to 70°C for 30 min. After incubation the proteolytic activity was measured. The enzyme activity at 40°C was highest and considered at 100%. By decreasing temperature to 30°C relative proteolytic activity decreased to 95%. With the increase of temperature relative proteolytic activity was slightly decreased (Fig. 4).
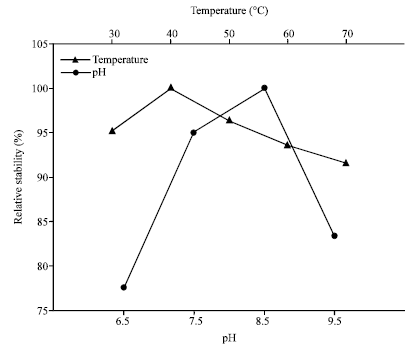 | |
| Fig. 4: | Relative thermal and pH stability of proteolytic activity of the protease isolated from Halobacterium sp. AF1 |
After incubation at 70°C this enzyme retained their proteolytic activity by 95%. Here, enzyme showing highest proteolytic activity for pH 8.5 and considered as 100%. Enzyme retained its proteolytic activity with the relative activity by 95% for pH 7.5 and above 80% (83%) for pH 9.5 (Fig. 4).
Effects of organic solvents and metal ions on the stability of AF1 protease: The relative activity which remained after 30 min of incubation in 25% (v/v) of organic solvent were determined for examining relative stability of protease. For control, organic solvent was replaced by distilled water and measured proteolytic activity was considered as 100%. Here, relative stability was increased to 160% for acetone and above 100% for n-hexane. Relative activity was around 100% for toluene (Fig. 5). The effects of 5 mM metal ions (e.g., Ca2+, Na+, Hg+2, Fe+3) on protease activity was determined by incubating the enzyme with different metal salts at room temperature for 30 min and thereafter the relative activities were determined under standard assay conditions. Only NaCl stimulate the relative stability of protease (Fig. 5). FeCl3 retained the relative stability of protease by 100%. Others, HgCl2 and CaCl2 retained the relative activity over 90%. The protease activity of salt free enzyme samples was considered as 100%.
Effects of surfactant and oxidant on AF1 protease: The enzyme activity of a control sample (without surfactants and oxidants) was considered as 100%. Relative protease activity was retained to around 70% for 1.5% of H2O2 and around 60% for 1.5% of SDS (Fig. 6).
Effects of inhibitors on proteolytic activity of AF1 protease: Here, proteolytic activity of inhibitor free enzyme samples was considered as 100%.
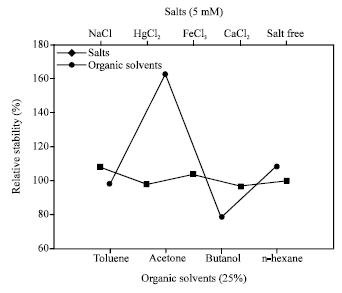 | |
| Fig. 5: | Changes in relative stability of protease isolated from Halobacterium sp. AF1 for different organic solvents and metal ions |
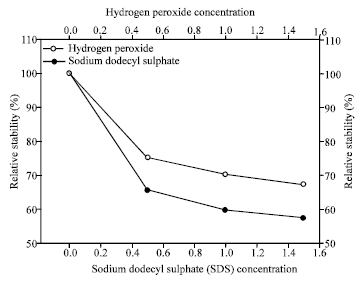 | |
| Fig. 6: | Effect of sodium dodecyl sulphate and H2O2 on the alkaline protease produced by Halobacterium sp. AF1 |
Results suggested that protease retained its relative activity to around 100% for β-mercapto ethanol (2-ME) (Fig. 7) and total protease activity was retained by up to 90% in the presence of 5 mM Ethylene Diamine Tetraacetic Acid (EDTA).
Hydrolysis of protein substrates by AF1 protease: For hydrolysis of protein substrate, various protein substrates (gelatin, casein and BSA) were incubated with protease. Here, the specific protease activity towards casein was taken as a control. Results showed that proteolysis against gelatin (105%) was higher compare to casein (100%) or BSA (bovine serum albumin) (95%) (Fig. 8).
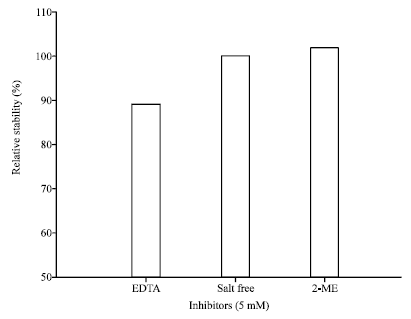 | |
| Fig. 7: | Effect of inhibitors on proteolytic activity of AF1 protease |
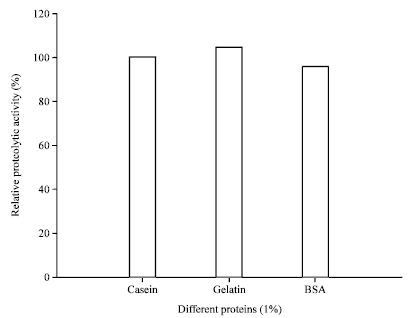 | |
| Fig. 8: | Alkaline protease activity produced from Halobacterium sp. AF1 against different natural substrates |
DISCUSSION
Optimum incubation period for protease production by Halobacterium sp. AF1 was 72 h. Similar incubation period for maximum protease production was observed from Halobacterium sp. SP1(1) at 37°C (Akolkar, 2009). The temperature is a critical parameter which needs to be controlled and this is usually varied from one organism to another (Kumar and Takagi, 1999). Optimum temperature for the production of protease by Halobacterium sp. AF1 was 40°C and incubation time was 72 h. Both bacterial growth and enzymatic activity of Halobacterium sp. was optimum at 40°C was also reported (Anand et al., 2010). The production of alkaline protease by Bacillus halodurans were investigated by Ibrahim and Al-Salamah (2009) while maximal cell growth was seen at 50°C and maximum enzyme production was found at 37°C. The optimum pH for the production of protease by Halobacterium sp. AF1 was 8. The bacterial growth was also highest at this pH. The crude protease enzyme, produced from B. subtilis strain 38, had the optimum pH at 6.25 (Chantawannakula et al., 2002). Malathu et al. (2008) reported an extra-cellular protease from a novel bacterial isolate showing the maximum activity at pH 7.5. Nascimento and Martins (2004) reported an optimum pH of 8.0 for a protease derived from thermophilic Bacillus sp strains SMIA-2. The highest proteolytic activity during this study was 13.5 U mL-1. Haq et al. (2003) reported the maximum protease activity during the course of study was 4.8 U mL-1.
Stability of AF1 protease was highest at 40°C and retained their activity up to 95% at 70°C. Subtilisin-like serine protease from Halobacterium salinarum P-535 reported by Izotova et al. (1983) had maximum activity at 37°C between pH 8-9. Stable protease from Pseudomonas aeruginosa PST-01 was reported to be stable at the temperature below 50°C (Ogino et al., 2007). A solvent stable protease, from Pseudomonas aeruginosa PseA retained 80% of its initial activity after heating for 30 min at 55°C (Gupta and Khare, 2007). AF1 protease showed highest proteolytic activity at pH 8.5. Stability of AF1 protease decreased with increasing or decreasing pH. Capiralla et al. (2002) reported maximum activity of extracellular serine protease from Halobacterium halobium S9 at 40°C and alkaline pH 8.7. Generally, commercial proteases from microorganisms have maximum activities in the alkaline pH range of 8.0-12.0 (Ghorbel et al., 2003).
Acetone and n-hexane stimulated the proteolytic activity of AF1 protease. There are very few reports on solvent tolerance of halophilic archaea (Usami et al., 2003, 2005). Akolkar et al. (2008) suggested that protease isolated from Halobacterium sp. showed 96% (tolerance) relative stability for toluene and 99% for xylene and n-dodecane. Alkaline protease tolerant to xylene, ethanol, acetone, butanol, benzene and chloroform has been reported from salt tolerant Streptomyces clavuligerus strain Mit-1 (Thumar and Singh, 2009). AF1 protease showed magnificent stability for different metal ions. Na+ stimulated the proteolytic activity and Ca2+, Hg+2, Fe+3 showed marginal effect. The protease activity was enhanced with an addition of 5 mM MnSO4, NaCl and CaCl2, resulting in the relative activity of 112, 111 and 117%, respectively, suggesting that metal ions had a capability to protect enzyme against denaturation (Akel et al., 2009).
AF1 protease showed moderate stability to SDS. The enzyme from Halobacterium sp. SP(1) retained 62.15±1.9% activity in presence of 0.5% SDS (anionic detergent) whereas, the neutral detergents like tritonX-100 and tween 80 had no effect on enzyme activity (Akolkar, 2009). AF1 protease was moderately stable for H2O2. H2O2 is known to be a strong oxidizing agent and it mediates oxidative inactivation of proteins. Met has been identified as a primary site for oxidative inactivation of the enzyme and most of the subtilisins containing Met residue next to the catalytic Ser residue undergo oxidative inactivation after the treatment with oxidizing agents such as H2O2 (Siezen and Leunissen, 1997).
AF1 protease was negligibly inhibited by β-mercapto ethanol which indicated that this protease was not cysteine protease. This AF1 protease was not inhibited significantly by EDTA which suggested that the enzyme was not metalloproteases. Metalloprotease inhibitor such as EDTA and cysteine protease inhibitors iodoacetamide was used by Khan et al. (2008). So, this protease was possible serine protease. Extracellular proteases isolated so far from halophilic archaea were serine proteases and most appear closely related to the subtilisin-like serine protease subfamily S8A and have been denoted as halolysins (Decastro et al., 2006). Proteolysis of gelatin by AF1 protease was above 100% and of BSA was around 95%. Thermo-stable serine alkaline protease from Bacillus subtilis PE-11 hydrolysis casein to about 100%, because when assayed with native proteins as substrates, the protease showed a high level of hydrolytic activity (Adinarayana et al., 2003). So, it can be concluded that there is higher possibility of this enzyme in different industrial applications as like as in leather tanning and detergent additives.
CONCLUSIONS
Form the study the following conclusion can be made:
| • | The optimum production of protease (12.4 U ml-1) was achieved at 72 h of culture, at 40°C with the pH 8. Protease produced by Halobacterium sp. AF1 retained their activity up to 92% for 70°C. pH 8.5 was the optimum for Protease production by Halobacterium sp. AF1 and retained their activity up to 83% at pH 9.5 |
| • | Proteolytic activity was stimulated by acetone and n-hexane and retained their activity with toluene up to 98% and for butanol up to 78%. NaCl and FeCl3 stimulated the proteolytic activity but the protease isolated from Halobacterium sp. AF1 retained their activity up to 98 and 97% for HgCl2 and CaCl2, respectively |
| • | β-mercapto ethanol (β-ME) and Ethylene Diamine Tetraacetic Acid (EDTA) showed negligible inhibitory effect toward protease isolated from Halobacterium sp. AF1. Protease isolated from Halobacterium sp. AF1 retained their activity up to 67% for 1.5% H2O2 and 57% for 1.5% SDS (sodium dodecyl sulfate) |
| • | Alkaline protease activity produced from Halobacterium sp. AF1 against gelatin was higher than casein and bovine serum albumin |
REFERENCES
- Adinarayana, K., P. Ellaiah and D.S. Prasad, 2003. Purification and partial characterization of thermostable serine alkaline protease from a newly isolated Bacillus subtilis PE-11. AAPS Pharm. Sci. Technol., 4: 1-9.
Direct Link - Akolkar, A.V., G.M. Deshpande, K.N. Raval, D. Durai, A.S. Nerurkar and A.J. Desai, 2008. Organic solvent tolerance of Halobacterium sp. SP1(1) and its extracellularprotease. J. Basic Microbiol., 48: 421-425.
CrossRefPubMedDirect Link - Akel, H., F. Al-Quadan and T.K. Yousef, 2009. Characterization of a purified thermostable protease from hyperthermophilic Bacillus strain HUTBS71. Eur. J. Sci. Res., 31: 280-288.
Direct Link - Capiralla, H., T. Hiroi, T. Hirokawa and S. Maeda, 2002. Purification and characterization of a hydrophobic amino acid-specific endopeptidase from Halobacterium halobium S9 with potential application in debittering of protein hydrolysates. Process Biochem., 38: 571-579.
CrossRef - Chantawannakula, P., A. Oncharoena, K. Klanbuta, E. Chukeatiroteb and S. Lumyong, 2002. Characterization of proteases of Bacillus subtilis strain 38 isolated from traditionally fermented soybean in Northern Thailand. Sci. Asia, 28: 241-245.
Direct Link - Decastro, R.E., A.M. Julie, M.I. Gimenez, M.K.H. Seitz and J.J. Sanchez, 2006. Haloarchaeal proteases and proteolytic systems. FEMS Microbiol. Rev., 30: 17-35.
CrossRefPubMedDirect Link - Ghorbel, B., A. Sellami-Kamoun and M. Nasri, 2003. Stability studies of protease from Bacillus cereus BG1. Enzyme Microbial. Technol., 32: 513-518.
CrossRef - Gupta, R., B. Chauhan, P. Ramnani and R. Singh, 2005. Bacterial Alkaline Proteases: Recent Trends and Industrial Application. In: Microbial Diversity: Crrent Perspectives and Potential Applications, Satyanarayana, T. and B.N. Johri (Eds.). I.K. International Publishing House Pvt. Ltd., New Dehli, India, ISBN-13: 9788188237432, pp: 769-789.
- Gupta, A. and S.K. Khare, 2007. Enhanced production and characterization of a solvent stable protease from solvent tolerant Pseudomonas aeruginosa PseA. Enzyme Microb. Technol., 42: 11-16.
CrossRefDirect Link - Haq, I., H. Mukhtar, S. Daudi, S. Ali and M.A. Qadeer, 2003. Production of proteases by a locally isolated mould culture under lab conditions. Biotechnology, 2: 30-36.
CrossRefDirect Link - Ibrahim, A.S.S. and A.A. Al-Salamah, 2009. Optimisation of media and cultivation conditions for alkaline protease production by alkaliphilic Bacillus halodurans. Res. J. Microbiol., 4: 251-259.
CrossRefDirect Link - Izotova, L.S., A.Y. Strongin, L.N. Chekulaeva, V.E. Sterkin and V.I. Ostoslavskaya et al., 1983. Purification and properties of serine protease from Halobacterium halobium. J. Bacteriol., 155: 826-830.
PubMedDirect Link - Khan, H., M. Subhan, S. Mehmood, M.F. Durrani, S. Abbas and S. Khan, 2008. Purification and characterization of serine protease from seeds of Holarrhena antidysenterica. Biotechnology, 7: 94-99.
CrossRefDirect Link - Kumar, C.G. and H. Takagi, 1999. Microbial alkaline proteases from a bioindustrial viewpoint. Biotechnol. Adv., 17: 561-594.
CrossRefPubMedDirect Link - Kumar, D., Savitri, N. Thakur, R. Verma and T.C. Bhalla, 2008. Microbial proteases and application as laundry detergent additive. Res. J. Microbiol., 3: 661-672.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Malathu, R., S. Chowdhury, M. Mishra, S. Das and P. Moharana et al., 2008. Characterization and wash performance analysis of microbial extracellular enzymes from East Calcutta Wetland of India. Am. J. Applied Sci., 5: 1650-1661.
CrossRefDirect Link - do Nascimento, W.C.A. and M.L.L. Martins, 2004. Production and properties of an extracellular protease from thermophilic Bacillus sp. Braz. J. Microbiol., 35: 91-96.
CrossRefDirect Link - Ogino, H., T. Uchiho, N. Doukyu, M. Yasuda, K. Ishimi and H. Ishikawa, 2007. Effect of exchange of amino acid residues of the surface region of the PST-01 protease on its organic solvent-stability. Biochem. Biophys. Res. Commu., 358: 1028-1033.
CrossRefPubMedDirect Link - Patel, R., D. Mittal and S.P. Singh, 2005. Extracellular alkaline protease from a newly isolated haloalkaliphilic Bacillus sp.: Production and optimization. Process Biochem., 40: 3569-3575.
CrossRef - Pawar, R., V. Zambare, S. Barve and G. Paratkar, 2009. Application of protease isolated from Bacillus sp. 158 in enzymatic cleansing of contact lenses. Biotechnol., 8: 276-280.
CrossRefDirect Link - Rusal, M.G., I. Faisal and M.M.K. Khan, 2006. Environmental pollution generated from process industries in Bangladesh. Int. J. Environ. Pollut., 28: 144-161.
CrossRefDirect Link - Siezen, R.J. and J.A.M. Leunissen, 1997. Subtilases: The super family of subtilisin-like serine proteases. Protein Sci., 6: 501-523.
CrossRef - Tari, C., H. Genckal and F. Tokatli, 2006. Optimization of a growth medium using a statistical approach for the production of an alkaline protease from a newly isolated Bacillus sp. L21. Process Biochem., 41: 659-665.
CrossRef - Thumar, J.T. and S.P. Singh, 2009. Organic solvent tolerance of an alkaline protease from salt-tolerant alkaliphilic Streptomyces clavuligerus strain Mit-1. J. Ind. Microbiol. Biotechnol., 36: 211-218.
CrossRefPubMedDirect Link - Usami, R., T. Fukushima, T. Mizuki, Y. Yoshida, A. Inoue and K. Horihoshi, 2005. Organic solvent tolerance of halophilic archaea, Haloarcula strains: Effects of NaCl concentration on the tolerance and polar lipid composition. J. Biosci. Bioeng., 99: 169-174.
CrossRefPubMedDirect Link - Gupta, R., Q. Beg and P. Lorenz, 2002. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol., 59: 15-32.
CrossRefPubMedDirect Link - Joo, H.S., C.G. Kumar, G.C. Park, K.T. Kim, S.R. Paik and C.S. Chang, 2002. Optimization of the production of an extracellular alkaline protease from Bacillus horikoshii. Process Biochem., 38: 155-159.
CrossRef