
Abdelnasser S.S. Ibrahim
Department of Botany and Microbiology, Faculty of Science, King Saud University, P.O. Box 2454, Riyadh 11451, Saudi Arabia
Ali A. Al-Salamah
Department of Botany and Microbiology, Faculty of Science, King Saud University, P.O. Box 2454, Riyadh 11451, Saudi Arabia
Research Journal of Microbiology
Year: 2009 | Volume: 4 | Issue: 7 | Page No.: 251-259







ABSTRACT
Media and cultivation conditions were investigated to optimize alkaline protease production by alkaliphilic Bacillus halodurans. This includes different carbon, nitrogen and metals sources in addition to different pH, incubation temperature and aeration level. The specific enzyme activity was increased by about 48.8 fold by optimizing different nutrient sources and cultivation conditions. The maximum specific enzyme activity was obtained in a medium containing 15 g L-1 lactose as the carbon source, 6 g L-1 soybean as the nitrogen source and a 5 mM mixture of Mg, Mn and Ca as trace elements, fermentation for 48 h at 37°C and agitation at 200 rpm. This study indicated the significance of nutrient source and cultivation conditions on the alkaline enzyme production by Bacillus halodurans.
PDF Abstract XML References Citation
How to cite this article
Abdelnasser S.S. Ibrahim and Ali A. Al-Salamah, 2009. Optimisation of Media and Cultivation Conditions for Alkaline Protease Production by Alkaliphilic Bacillus halodurans. Research Journal of Microbiology, 4: 251-259.
URL: https://scialert.net/abstract/?doi=jm.2009.251.259
URL: https://scialert.net/abstract/?doi=jm.2009.251.259
INTRODUCTION
With increasing emphasis on environmental protection, the use of enzymes particularly from extremophiles has gained considerable attention during the last several years. Extremozymes are now replacing chemical catalysts in many industries, including manufacturing of chemicals, textiles, pharmaceuticals, paper, food and agricultural chemicals (Mehta et al., 2006). Alkaliphiles are reported to be a rich source of alkaline active enzymes, e.g., amylase, protease, cellulose and xylanase and other enzymes that have numerous applications in many industrial processes (Horikoshi, 1999a; Oskouie et al., 2008).
Proteases, also known as peptidyl-peptide hydrolases (EC 3.4.21-24 and 99), are degradative enzymes that catalyse the cleavage of peptide bonds in other proteins. Currently, proteases are classified on the basis of three major criteria: the type of reaction catalysed, the chemical nature of the catalytic site and evolutionary relationships with reference structures. Alkaline proteases are referred to as proteolytic enzymes that work optimally in alkaline pH ranges (1, 12). Alkaline proteases are the main enzymes among known proteases and constitute 60-65% of the global industrial enzyme market (Amoozegara et al., 2007). Most of them are used in the food industry in meat tenderisation processes, peptide synthesis, infant formula preparations, baking and brewing. They are also used in the detergent industry as additives, in pharmaceuticals and medical diagnosis as well as in the textile industry in the process of dehairing and leather processing (Tari et al., 2006; Bhaskar et al., 2007; Dodia et al., 2008).
Extracellular protease production in microorganisms is highly influenced by media Variation in components such as C/N ratio, the presence of easily metabolisable sugars like glucose (Gupta et al., 2002a; Ferrero et al., 1996) and the presence of metal ions (Varela et al., 1996). Besides this, several other factors, such as aeration, inoculum density, pH, temperature and incubation time, also affect the amount of protease produced (Hameed et al., 1999; Gupta et al., 2002b). The goal of the present study was to optimise the medium and cultivation conditions for alkaline protease production by alkaliphilic Bacillus halodurans previously isolated from an Egyptian soda lake (Ibrahim and El-diwany, 2007).
MATERIALS AND METHODS
Microorganism and Culture Maintenance
The microorganism used in this study was isolated from indigenous soil samples from Wadi El-Natrun, an Egyptian Soda Lake and were screened using a skim milk agar plate and later in alkaline broth (Ibrahim and El-diwany, 2007). The isolate was identified as Bacillus halodurans according to morphological and biochemical tests and 16S rDNA sequence analysis (Ibrahim and El-diwany, 2007).
Stock cultures of the isolate were stored in 30% glycerol at-70°C. Prior to each experiment, the bacterium was sub-cultured from the frozen stocks onto Horikoshii agar medium (pH 10.5) containing 10 glucose, 5 peptone, 5 yeast extract, 1 KH2PO4, 0.2 MgSO4•7H2O, 10 Na2CO3 and 15 g g L-1 agar (Horikoshi, 1999b). Alkaline broth was used as the basal medium for preliminary studies of bacterial growth and protease production.
Production of Alkaline Proteases
A loop of culture from an agar plate was inoculated into a 50 mL glass tube containing 5 mL of alkaline protease production medium and incubated overnight at 180 rpm and 50°C. This culture was then inoculated into a 500 mL capacity Erlenmeyer flask containing 95 mL of the same medium and incubated at 50°C for 48 h. Cells and insoluble materials were removed by centrifugation at 10,000 g for 10 min at 4°C and the cell-free supernatant was filtered through a 0.45 μm pore-size membrane filter and was used as the source of the crude alkaline protease enzyme. Five different media were used for enzyme production: (1) Horikoshii medium, (2) Luria-Bertani broth (LB) plus 10 g L-1 Na2CO3, (3) tryptic soy broth (TSB) plus 10 g L-1 Na2CO3, (4) nutrient broth (NB) plus 10 g L-1 Na2CO3 and (5) a modification of a medium previously described by Beg et al. (2003) containing 5 casamino acids, 10 glucose, 1 KH2PO4, 3 K2HPO4, 2 Na2SO4, 0.1 MgSO4•7H2O and 10 g L-1 Na2CO3.
Enzyme Assay
Proteolytic activity was assayed by a modification of the method of Kunitz (1947). Samples containing 400 μL of 0.5% (w/v) casein in 50 mM Tris-HCl buffer, pH 10, with 100 μL of enzyme sample were incubated in a water bath at 50°C for 20 min. The enzyme reaction was terminated by addition of 500 μL of 10% (w/v) trichloroacetic acid and incubation at room temperature for 10 min.
The reaction mixture was centrifuged at 10,000 g for 10 min at 4°C and the absorbance was measured against a blank (non-incubated sample) at 280 nm. One unit of protease was defined as the amount of enzyme that releases the equivalent of 1 ìmol of tyrosine per minute under the defined assay conditions. Standard curve of tyrosine was constructed using 10, 20, 30, 40, 50, 60, 70, 80 and 100 mg mL-1 tyrosine in 50 mM glycine-NaOH buffer, pH 10.
Protein Determination
Protein concentrations were determined according to the method described by Bradford (1976). One millilitre of Bradford reagent was added to 50 μL of sample and the absorbance was measured after 5 min at 595 nm. Different concentrations of Bovine Serum Albumin (BSA) were used as a protein standard, including 10, 20, 40, 60, 80 and 100 μg mL-1 in distilled water. One milliliter of Bradford reagent was added to 50 μL BSA standard and after 5 min the absorbance was measured at 595 nm.
Optimization of Alkaline Protease Production
The alkaline protease production optimization studies of Bacillus halodurans included different media and conditions, e.g., five different media with different carbon and nitrogen sources, including cheap nutrient sources with different concentrations and metal ions. The effect of different cultivation conditions, including incubation temperatures (25-55°C), aeration conditions (0-300 rpm), incubation times and inoculum sizes, were investigated. All experiments were done in triplicate.
RESULTS AND DISCUSSION
Optimization of Alkaline Protease Production
The microorganism used in this study was isolated from indigenous soil samples of Wadi El-Natrun, an Egyptian soda lake and were screened using a skim milk agar plate and later in alkaline broth (Ibrahim and El-diwany, 2007). The organism was identified as Bacillus halodurans according to morphological and biochemical tests and 16S rDNA sequence analysis (Ibrahim and El-diwany, 2007). The crude alkaline protease showed reasonable activity at a temperature range of 65 to 75°C with maximum activity at 70°C. The crude enzyme also had a relatively wide pH range of activity between pH 8 to 11, with maximum enzyme activity at pH 10 in 50 mM Tris-HCl buffer, indicating the enzyme is a thermo-alkaline protease (Ibrahim and El-diwany, 2007).
The time course of alkaline protease production showed that maximum enzyme production was seen after 48 h of incubation (data not shown). Five different alkaline media were tested for maximum alkaline protease production by Bacillus halodurans. The results presented in Fig. 1 indicated that Medium I was the best medium for alkaline protease production by Bacillus halodurans (70.2 U mg-1 protein). Therefore, Medium I was selected for further analysis.
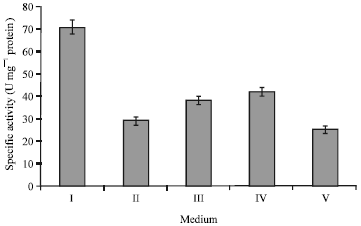 | |
| Fig. 1: | Effects of different media on alkaline protease production by Bacillus halodurans at pH 10.5 and 50°C after 48 h of incubation |
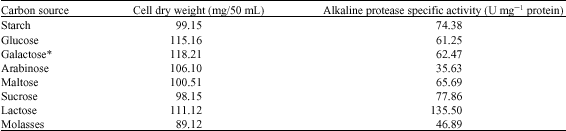
| Table 1: | Effects of different carbon sources on alkaline protease production by Bacillus halodurans at pH 10.5 and 50°C after 72 h of incubation |
 | |
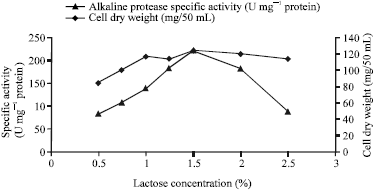 | |
| Fig. 2: | Effects of different lactose concentrations on alkaline protease production by Bacillus halodurans at pH 10.5 and 50°C after 72 h of incubation |
Effects of Carbon Sources
There are several reports that different carbon sources have different influences on extracellular enzyme production by different strains (Chi and Zhao, 2003; Beg et al., 2003; Kanekar et al., 2002). Therefore, the effect of different carbon sources on alkaline protease production by B. halodurans was investigated. The results presented in (Table 1) show that lactose was the best carbon source for protease production with a specific activity of 135.5 U mg-1 protein, which indicated that B. halodurans was able to produce β-galactosidase to hydrolyze lactose and consume glucose and galactose as carbon sources. This result is similar to that reported for alkaline protease production by Salinvibrio sp., where lactose was the best carbon source for enzyme production (Amoozegara et al., 2007). However, increased alkaline protease production was also reported by several other studies that used different sugars, such as starch, sucrose, fructose and maltose (Genckal and Tarib, 2006; Reddy et al., 2008), indicating that the best carbon source for enzyme production is different according to different microorganisms. While glucose showed the highest cell growth (115.16 mg/50 mL), it showed low enzyme production, indicating that alkaline protease production was catabolically repressed by glucose (Mehta et al., 2006; Kaur et al., 2001). The enzyme specific activity increased with increasing lactose concentrations (Fig. 2) with a maximum enzyme production at 1.5% lactose (224.66 U mg-1 protein). Further increases in the lactose concentration resulted in reductions of enzyme production, which was mostly due to catabolic repression of enzyme production by the accumulation of glucose produced after lactose hydrolysis (Mehta et al., 2006). Optimizing the carbon source resulted in an increase in enzyme production of about three fold.
Effects of Different Nitrogen Sources on Alkaline Protease Production
In microorganisms, nitrogen (both organic and inorganic forms) is metabolized to produce amino acids, nucleic acids, proteins and cell wall components. Alkaline protease production depends heavily on the availability of nitrogen sources in the medium, which has regulatory effects on enzyme synthesis (Patel et al., 2005).
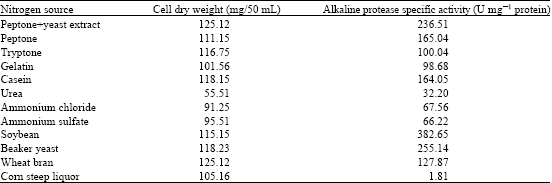
| Table 2: | Effects of different nitrogen sources on alkaline protease production by Bacillus halodurans at pH 10.5 and 50°C 48 h of incubation |
 | |
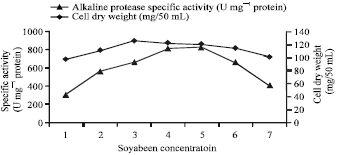 | |
| Fig. 3: | Effect of different soybean concentration on alkaline protease production by Bacillus halodurans at pH 10.5 and 50°C after 48 h incubation |
The effects of the nitrogen source on alkaline protease production by Bacillus halodurans, including different inorganic, organic and cheap nitrogen sources, were examined. As shown in Table 2, the inorganic nitrogen sources urea, ammonium chloride and ammonium sulfate showed the lowest alkaline protease productions of 32.2 protein, 66.22 protein and 67.56 U mg-1 protein, respectively. Interestingly, soybean, which is a cheap nitrogen source, showed the highest enzyme production of the nitrogen sources used (382.65 U mg-1 protein). Soybean was shown to stimulate protease production in the marine bacterium Teredinobacter turnirae (Elibola and Moreira, 2005), while protease production in haloalkaliphilic Bacillus sp. was enhanced by a combination of peptone and yeast extract (Patel et al., 2005). However, the inorganic nitrogen source sodium nitrate was shown to stimulate alkaline protease production in another microorganism (Chi et al., 2007), indicating that the best nitrogen source for enzyme production is different according to different microorganisms. Alkaline protease production by Bacillus halodurans was increased by increasing soybean concentrations and reached a maximum level at 6 g L-1 (835.6 U mg-1 protein) and decreased with further increasing soybean concentrations (Fig. 3). A similar effect of soybean was reported in Teredinobacter turnirae (Elibola and Moreira, 2005). Optimization of the nitrogen source resulted in a further increase in alkaline protease production of about 3.5 fold.
Effects of Different Metals on Alkaline Protease Production by B. halodurans
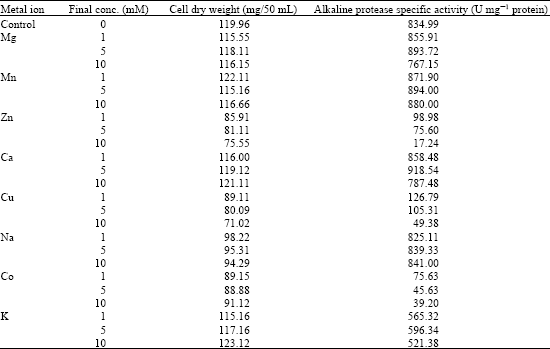
The effects of different metal ions on the production of alkaline protease by Bacillus halodurans were investigated. Among different metals examined, Mg, Mn and Ca showed the highest enzyme productions at concentrations of 5 mM with specific activities of 893.73, 905.70 and 918.54 U mg-1 protein, respectively (Table 3).
| Table 3: | Effects of different metal concentrations on alkaline protease production by Bacillus halodurans at pH 10.5 and 50°C after 48 h of incubation |
 | |
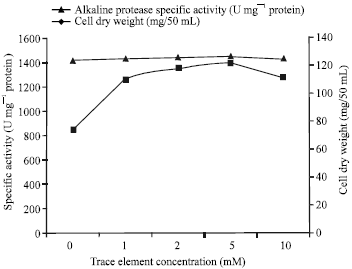 | |
| Fig. 4: | Effects of different concentrations of a trace element mixture (Ca, Mn and Mg) on alkaline protease production by Bacillus halodurans at pH 10.5 and 50°C after 48 h of incubation |
Zn, Cu and Co strongly inhibited enzyme production. Incorporation of Ca, Mn and Mg together in the growth medium showed higher enzyme production than using them individually with maximum production at 5 mM of the trace element mixture (1381 U mg-1 protein) (Fig. 4).
Effects of Different Incubation Periods on Growth and Alkaline Protease Production
It is known that temperature is one of the most critical parameters that has to be controlled in bioprocesses (Chi and Zhao, 2003). The effect of the incubation temperature on alkaline protease production by Bacillus halodurans was investigated at temperatures ranging from 25-55°C.
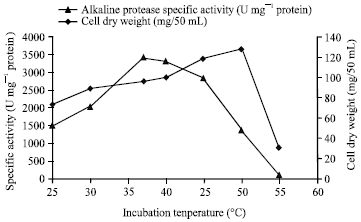 | |
| Fig. 5: | Effects of different incubation temperatures on alkaline protease production by Bacillus halodurans at pH 10.5 and after 48 h of incubation |
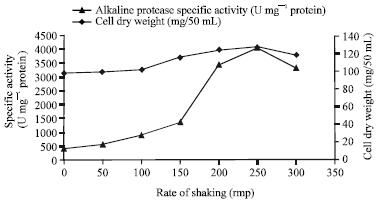 | |
| Fig. 6: | Effects of shaking speed on the production of alkaline protease by Bacillus halodurans at an incubation temperature of 37°C |
The results revealed that maximal cell growth was seen at 50°C (Fig. 5); however, maximum enzyme production was found at 37°C (3416.14 U mg-1 protein) with about a 2.5-fold increase in the specific enzyme activity. Other researchers have reported that there is an inverse relationship between alkaline protease production and cell growth (Genckal and Tarib, 2006). It has been reported that the optimum temperature for alkaline protease production by Aureobasidum pullulans was much lower than the optimum growth temperature (Chi et al., 2007).
Effects of Agitation Rate
Aeration is another important parameter that affects enzyme production in bacteria. It has been reported that enzymes are also susceptible to mechanical force, which may disturb the elaborate shape of complex molecules to such a degree that denaturation occurs (Banerjee et al., 1999; Gupta et al., 2002a; Joo et al., 2002). Alkaline protease production by Bacillus halodurans was investigated at different agitation speeds. The results represented in (Fig. 6) revealed that enzyme production increases with increasing agitation speed. The maximum protease production was observed at 200 rpm (3416 U mg-1 protein). However, enzyme production was reduced by further increases in the agitation rate. A similar profile was determined for the effect of agitation speed on alkaline protease production by Teredinobacter turnirae (Elibola and Moreira, 2005). The effect of inoculum size and age was also investigated, but they showed no significant effects on alkaline protease production by Bacillus halodurans (data not shown).
CONCLUSION
An alkaliphilic bacterium isolated from an alkaline Egyptian soda lake and identified as Bacillus halodurans showed thermo-alkaline protease production. The media and cultivation conditions were investigated to optimise alkaline protease production. The goal of this study was to enhance the enzyme production which was achieved through optimization of nutrients and cultivation conditions.
The specific enzyme activity was increased by about 48.8 fold using different nutrient sources and cultivation conditions. The maximum specific enzyme activity was obtained in a medium containing 15 g lactose, 6 g soybean and 5 mM of a mixture of Mg, Mn and Ca after fermentation at 37°C and 200 rpm.
REFERENCES
- Amoozegar, M.A., A.Z. Fatemi, H.R. Karbalaei-Heidari and M.R. Razavi, 2007. Production of an extracellular alkaline metalloprotease from a newly isolated, moderately halophile, Salinivibrio sp. strain AF-2004. Microbiol. Res., 162: 369-377.
CrossRefPubMedDirect Link - Banerjee, U.C., R.K. Sani, W. Azmi and R. Soni, 1999. Thermostable alkaline protease from Bacillus brevis and its characterization as a laundry detergent additive. Proc. Biochem., 35: 213-219.
CrossRefDirect Link - Beg, Q.K., V. Sahai and R. Gupta, 2003. Statistical media optimization and alkaline protease production from Bacillus mojavensis in a bioreactor. Process Biochem., 39: 203-209.
CrossRefDirect Link - Bhaskar, N., E.S. Sudeepa, H.N. Rashmi and A.T. Selvi, 2007. Partial purification and characterization of protease of Bacillus proteolyticus CFR3001 isolated from fish processing waste and its antibacterial activities. Bioresour. Technol., 98: 2758-2764.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chi, Z., C. Ma, P. Wang and H.F. Li, 2007. Optimization of medium and cultivation conditions for alkaline protease production by the marine yeast Aureobasidium pullulans. Bioresour. Technol., 98: 534-538.
CrossRefDirect Link - Chi, Z. and S. Zhao, 2003. Optimization of medium and cultivation conditions for pullulan production by a new pullulan-producing yeast strain. Enzyme Microb. Technol., 33: 206-211.
CrossRefDirect Link - Dodia, M.S., H.G. Bhimani, C.M. Rawal, R.H. Joshi and S.P. Singh, 2008. Salt dependent resistance against chemical denaturation of alkaline protease from a newly isolated haloalkaliphilic Bacillus sp. Bioresour. Technol., 99: 6223-6227.
PubMedDirect Link - Elibola, M. and A.R. Moreira, 2005. Optimizing some factors affecting alkaline protease production by a marine bacterium Teredinobacter turnirae under solid substrate fermentation. Process Biochem., 40: 1951-1956.
CrossRefDirect Link - Ferrero, M.A., G.R. Castro, C.M. Abate, M.D. Baigori and F. Singeriz, 1996. Thermostable alkaline protease of Bacillus licheniformis MIR 29: Isolation, production and characterization. Applied Microbiol. Biotechnol., 45: 327-332.
CrossRefDirect Link - Genckal, H. and C. Tari, 2006. Alkaline protease production from alkalophilic Bacillus sp. isolated from natural habitats. Enzyme Microb. Technol., 39: 703-710.
CrossRefDirect Link - Gupta, R., Q. Beg, S. Khan and B. Chauhan, 2002. An overview on fermentation, downstream processing and properties of microbial alkaline proteases. Applied Microbiol. Biotechnol., 60: 381-395.
CrossRefDirect Link - Gupta, R., Q. Beg and P. Lorenz, 2002. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol., 59: 15-32.
CrossRefPubMedDirect Link - Hameed, A., T. Keshavarz and C.S. Evans, 1999. Effect of dissolved oxygen tension and pH on the production of extracellular protease from a new isolate of Bacillus subtilis K2, for use in leather processing. J. Chem. Technol. Biotechnol., 74: 5-8.
Direct Link - Horikoshi, K., 1999. Alkaliphilies: Some applications of their products for biotechnology. Microb. Molecul. Biol. Rev., 63: 735-750.
Direct Link - Ibrahim, A.S.S. and A.I. El-diwany, 2007. Isolation and identification of new cellulases producing thermophilic bacteria from an Egyptian hot spring and some properties of the crude enzyme. Aust. J. Basic Applied Sci., 1: 473-478.
Direct Link - Joo, H.S., C.G. Kumar, G.C. Park, K.T. Kim, S.R. Paik and C.S. Chang, 2002. Optimization of the production of an extra cellular alkaline protease from Bacillus horikoshii. Process Biochem., 38: 155-159.
Direct Link - Kanekar, P.P., S.S. Nilegaonkar, S.S. Sarnaik and A.S Kelkar, 2002. Optimization of protease activity of alkaliphilic bacteria isolated from an alkaline lake in India. Bioresour. Technol., 85: 87-93.
PubMedDirect Link - Kaur, S., R.M. Vohra, M. Kapoor, Q. Khalil Beg and G.S. Hoondal, 2001. Enhanced production and characterization of a highly thermostable alkaline protease from Bacillus sp. P-2. World J. Microbiol. Biotechnol., 17: 125-129.
CrossRefDirect Link - Kunitz, M., 1947. Crystalline Soyabean trypsin inhibitor II: General properties. J. Gen. Physiol., 30: 291-310.
Direct Link - Mehta, V.J., J.T. Thumar and S.P. Singh, 2006. Production of alkaline protease from an alkaliphilic actinomycete. Bioresour. Technol., 97: 1650-1654.
CrossRefDirect Link - Oskouie, S.F.G., F. Tabandeh, B. Yakhchali and F. Eftekhar, 2008. Response surface optimization of medium composition for alkaline protease production by Bacillus clausii. Biochem. Eng. J., 39: 37-42.
CrossRefDirect Link - Patel, R., D. Mittal and S.P. Singh, 2005. Extracellular alkaline protease from a newly isolated haloalkaliphilic Bacillus sp.: Production and optimization. Process Biochem., 40: 3569-3575.
CrossRef - Reddy, L.V.A., Y.J. Wee, J.S. Yun and H.W. Ryu, 2008. Optimization of alkaline protease production by batch culture of Bacillus sp. RKY3 through plackett-burman and response surface methodological approaches. Bioresour. Technol., 99: 2242-2249.
CrossRefPubMedDirect Link - Tari, C., H. Genckal and F. Tokatli, 2006. Optimization of a growth medium using a statistical approach for the production of an alkaline protease from a newly isolated Bacillus sp. L21. Process Biochem., 41: 659-665.
CrossRef - Varela, H., M.D. Ferrari, L. Belobradjic, R. Weyrauch and M.L. Loperena, 1996. Effect of medium composition on the production by a new Bacillus subtilis isolate of protease with promising unhairing activity. World J. Microbiol. Biotechnol., 12: 643-645.
CrossRefDirect Link