
Sidra Aftab
Department of Microbiology, University of Karachi, Karachi-75270, Pakistan
Samia Ahmed
Department of Microbiology, University of Karachi, Karachi-75270, Pakistan
Sadia Saeed
Department of Microbiology, University of Karachi, Karachi-75270, Pakistan
Sheikh Ajaz Rasool
Department of Microbiology, University of Karachi, Karachi-75270, Pakistan
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 11 | Page No.: 2122-2126







ABSTRACT
Fifty-three alkaliphile bacterial isolates were collected from various fields of cosmopolitan city of Karachi, out of which 25 were alkaline protease producers. Bacillus brevis SSA1 was selected as the best enzyme producer and was used for further physico-chemical studies. B. brevis SSA1 started producing alkaline protease SSA1 after 74 h of incubation at 35°C. Activity potential of enzyme by agar-well diffusion method was found to be 800 AU mL-1. Although enzyme produced by B. brevis SSA1 was active over a wide range of pH (7-13), however, its optimal pH was found to be 8.0. The optimal temperature for enzyme activity was 60°C while enzyme activity remains stabled up to 80°C but gets inactivated at higher temperatures. Enzyme activity was totally inhibited in the presence of 5.0 mM PMSF that suggests the enzyme was related to serine alkaline protease. EDTA (inhibitor of metalloprotease) did not show any detrimental effect on the enzyme activity. Mercaptoethanol, SDS and H2O2 had no effect on alkaline protease activity. Metal ions like Cd+2, Ag+ and Ni+2 had some inhibitory effect while activity was completely lost in the presence of Zn+2. Alkaline protease SSA1 was not only stable in the presence of commercial detergent but also gave better result of washing with detergent, hence it can be used as additive in detergents for washing purposes.
PDF Abstract XML References Citation
How to cite this article
Sidra Aftab, Samia Ahmed, Sadia Saeed and Sheikh Ajaz Rasool, 2006. Screening, Isolation and Characterization of Alkaline Protease Producing Bacteria from Soil. Pakistan Journal of Biological Sciences, 9: 2122-2126.
DOI: 10.3923/pjbs.2006.2122.2126
URL: https://scialert.net/abstract/?doi=pjbs.2006.2122.2126
DOI: 10.3923/pjbs.2006.2122.2126
URL: https://scialert.net/abstract/?doi=pjbs.2006.2122.2126
INTRODUCTION
Microorganisms excrete a wide variety of proteolytic enzymes, which are also found in mammalian systems. They are molecules of relatively small size and are compact, spherical structures that catalyze the peptide bond cleavage in proteins (Polgar, 1989). They hydrolyze peptide bonds and therefore, lead to the disassembly of proteins. Commercially they are very important as more than 60% of the total enzyme market relies on proteases, isolated from plants, animals, bacteria and fungi. Proteases are (physiologically) necessary for living organisms; they are ubiquitous, found in a wide diversity of sources.
Proteases from microbial sources are preferred over the enzymes from plant or animal sources since they possess almost all the characteristics desired for their biotechnological applications. Microbial proteases represent a good source of enzymes due to a number of characteristics like their broad biochemical diversity, their rapid growth, the limited space required for cell cultivation and the ease with which the enzymes can be genetically manipulated to generate new enzymes for various applications (Rao et al., 1998). Although a wide range of microorganisms are known to date to produce proteases, a large proportion of the commercially available alkaline proteases are derived from Bacillus strains because of its ability to secrete large amounts of alkaline proteases having significant proteolytic activity and stability at considerably high pH and temperatures (Yang et al., 2000).
Proteases (depending on their activity at different pH) may be acidic, neutral or alkaline protease. Alkaline proteases useful for detergent applications were mostly active in the pH range 8-12 and at temperatures between 50 and 70°C (Al-Shehri et al., 2004).
Proteases of the subtilisin group are used in the treatment of burns and wounds. Oral administration of proteases produces an anti inflammatory response in burns patients and speeds up the process of healing (Thangam et al., 2002).
Although a variety of proteolytic fungi and bacteria are known, few provide high activities with commercial success. The main industrial application for alkaliphilic proteases is in the detergent industry (Gupta et al., 1999, 2002), leather tanning process (Hameed et al., 1996), food industries (Kaliz, 1988) and silver removal from X- ray films (Fujiwara et al., 1991).
For an enzyme to be used as a detergent additive it should be stable and active in the presence of typical detergent ingredients, such as surfactants, builders, bleaching agents, bleach activators, fillers, fabric softeners and various other formulation aids (Anwar and Saleemuddin, 2000).
One of the major drawbacks affecting the stability at alkaline pH of enzymes recovered from thermophiles is that enzymes from alkalophiles confer stability in a wide pH range but are usually thermolabile. Thus it is desirable to search for new proteases with novel properties from as many different sources as possible. The present study reports the screening of soil bacteria for their capability of alkaline protease production and characterization of the enzyme from the best producer.
MATERIALS AND METHODS
This study was conducted at the laboratory of Molecular Genetics, Department of Microbiology, University of Karachi, Pakistan.
Isolation of bacteria from soil: Fifty three bacterial strains were isolated on alkaline agar medium (1% glucose, 0.5% peptone, 0.5% yeast extract, 1% KH2PO4, 0.02% MgSO4. 7H2O, 1% Na2CO3 and 2% agar) from various soil fields of Karachi (Fereshteh et al., 2003) during the year 2005 and identified on the basis of morphological, cultural and biochemical characteristics (Cheesbrough, 2000).
Screening for alkaline protease production: All fifty-three strains were screened for their ability of alkaline protease production at 35°C on alkaline agar medium incorporated with casein (0.1% glucose, 0.5% tryptone, 0.25% yeast extract, 1% casein, 0.44% trisodium citrate, 20 mM CaCl2.6H2O and 2% agar) by stab-overlay method as described by Rasool et al. (1996) and Fereshteh et al. (2003). Clear zone around colony was the indication of alkaline protease production.
Extraction and purification of the enzyme: The bacterium that gave biggest zone of inhibition in above described assay was selected as best producer. Hence Bacillus brevis SSA1 was selected as best producer and was grown in alkaline broth at 35°C for 72 h with gentle shaking at 60 rpm. The broth culture was then centrifuged at 6000 rpm at 4°C and supernatant was collected. The enzyme activity and activity units/mL in supernatant was checked by agar-well diffusion method in casein containing media (Rasool et al., 1996). Supernatant was 80% partially purified by ammonium sulfate precipitation and activity was again checked (Adinarayana et al., 2003). Total protein concentration in the enzyme solution was determined by Lowry’s method (Lowry et al., 1951).
Physicochemical characterization of alkaline protease: Activity of the enzyme of Bacillus brevis SSA1 at different temperatures and pH values was checked (Al-Shehri et al., 2004; Thangam and Rajkumar, 2002). Enzyme activity was also determined in the presence of different metal ions, surfactants and protease inhibitors (Joo et al., 2003). Hydrolytic activity of enzyme on different substrates (BSA, gelatin, haemoglobin) was also checked (Adinarayana et al., 2003). Growth curve was also run to determine the active growth phase of enzyme production as per (Gupta et al., 2002).
Washing test: Application of alkaline protease enzyme by Bacillus brevis SSA1 as a detergent additive was studied as per Thangam and Rajkumar (2002). For this three white cotton cloth pieces (5x5 cm) were stained with blood and grass separately and (A = a) following sets were then prepared and studied:
| • | Blood or grass stained cloth dipped in flask with distilled water (100 mL). |
| • | Blood or grass stained cloth dipped in flask with distilled water (100 mL)+1 mL detergent (7 mg mL-1). |
| • | Blood or grass stained cloth dipped in flask with distilled water (100 mL) + 2 mL enzyme solution. |
| • | Blood or grass stained cloth dipped in flask with distilled water (100 mL) + 1 mL detergent (7 mg mL-1) + 2 mL enzyme solution. |
All four flasks were incubated at 60°C for 15 min. After incubation, cloth pieces were taken out, rinsed with water and dried. Visual examination of cloth pieces exhibited the effect of enzyme in removal of stains. Untreated cloth pieces stained with blood and grass were taken as control.
RESULTS AND DISCUSSION
Proteases are group of enzymes, which hydrolyze peptide bonds in proteins and peptides. Microbial proteases have a number of commercial applications. A major commercial use is the addition of microbial proteases to domestic detergents for the digestion of proteinaceous stains of fabrics (Sharmin et al., 2004). It has been reported that the production of extracellular proteases by different microorganisms can be strongly influenced by the culture conditions. So it becomes necessary to understand the nature of proteases and their catalytic potentiality under different conditions. Fifty-three alkaliphiles were isolated from different soil fields of Karachi and were identified up to species level. Bacillus sp. was found to be predominant in soil.
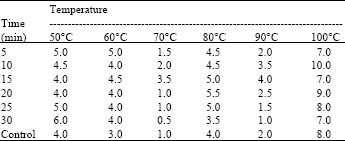
| Table 1: | Temperature stability of alkaline protease enzyme produced by B. brevis SSA1 (Zone of hydrolysis in mm) |
 | |
| Table 2: | pH stability of alkaline protease enzyme produced by B. brevis SSA1 |
 | |
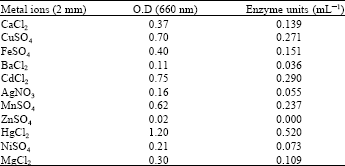
| Table 3: | Effect of metal ions on alkaline protease activity |
 | |
| Table 4: | Washing test of alkaline protease SSA1 |
 | |
| + = Poor removal of stain; ++ = Good removal of stain; +++ = Very good removal of stain | |
Hence 2% were found to be B. coagulans, B. stearothermophilus and B. licheniformis, 4% were B. cereus and B. circulans, 6% were B. alvei, 8% were B. laterosporus, 10% were B. pumilus. 20% were B. brevis and 21% were B. sphaericus and B. macerans. Only 2% were found to be M. luteus. Out of fifty-three strains only 25 (47%) were found to be alkaline protease enzyme producer. Alkaline protease enzyme production by Bacillus species has been previously reported by Aoyama et al. (2000), Feng et al. (2001), Adinarayana et al. (2003), Al-Shehri et al. (2004) and Schallmey et al. (2004). Subtilisin like alkaline proteases is generally bacterial in origin. They are secreted extracellularly for the purpose of scavenging nutrients (Graycar, 1999). In this study the best producer of alkaline protease was identified as Bacillus brevis, designated as Bacillus brevis SSA1 and the enzyme produced by the organism was designated as alkaline protease SSA1. Alkaline protease production has been reported in both Pseudomonas and Bacillus sp. (Horikoshi, 1971; Najafi et al., 2005). B. brevis SSA1 was identified as alkaliphilic, gram-positive, strictly aerobic, motile, catalase positive, VP negative, spore forming bacterium. The alkaline protease production by B. brevis SSA1 was found in alkaline medium (at pH 10.5). Previously, Fereshteh et al. (2003) also reported production of alkaline protease by Bacillus strain L2 at pH 10.5.
The activity of enzyme SSA1 was totally inhibited by PMSF and not affected by treatment with EDTA (metalloprotease inhibitor), which clearly showed that enzyme, belongs to serine group of alkaline protease. Feng et al. (2001) and Kobayashi et al. (1995) also reported complete inhibition by PMSF of alkaline protease activity produced by Bacillus pumilus and Bacillus sp. KSM-K16, respectively.
Alkaline protease production occurs in the late stationary phase of growth and increases as GTP level decreases (Gupta et al., 2002). In our case alkaline protease production also started after 72 h of incubation, hence it can be categorized as secondary metabolite. Thermostable and stability at wide range of pH are desirable properties of any enzyme for industrial applications. In our case the optimum temperature of enzyme activity was 60°C (Table 1), as also reported by Adinarayana et al. (2003) and Denizci et al. (2004) while optimum pH for its activity was recorded as 8.0 (Table 2). Most of the alkaline proteases have been reported to have their optimal pH 8-9 (Gusekand and Kinsella, 1987; Aoyama et al., 2000; Peek et al., 1992). The enzyme possesses proteolytic activity from temperatures below 15 to above 70°C. It showed that it was a thermostable enzyme and this property makes the enzyme an ideal candidate for detergent application. High temperature stability was also reported previously (Gessesse et al., 2003; Al-Shehri et al., 2004). The alkaline protease SSA1 had broad pH range stability 7-13. It makes the enzyme suitable to be used in leather industry for hide-dehairing process because dehairing is carried out at pH values between 8 and 10. Present results are in close proximity with previous researchers (Takami et al., 1992). Ag+ and Ni+2 had some inhibitory effect while activity was completely lost by Zn+2 (Table 3). The enzyme was stabilized in the presence of Ca+2 and to a lesser extent by other metal ions, such as Mn+2, Cd+2 and Fe+2. The enzyme was also stable in the presence of 5 mM SDS and oxidants like H2O2, which are the common ingredients in modern bleach-based detergent formulations.
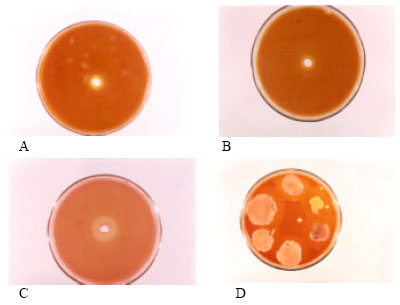 | |
| Fig. 1: | Hydrolytic effect of alkaline protease enzyme on different substrates (A. Gelatin, B. BSA, C. Hemoglobin D. Hydrolysis of casein by different alkaline protease producing bacteria |
It shows the alkaline protease SSA1 stability in detergent ingredients. Najafi et al. (2005) and Joo et al. (2003) also reported the same results previously. Alkaline protease SSA1 was active over a broad range of substrates i.e., casein, BSA, gelatin and haemoglobin (Fig. 1) which makes the enzyme to be used for many novel purposes like due to its gelatinize activity, can be used for recovery of silver from photographic films (Najafi et al., 2005). As alkaline protease SSA1 was not only stable in the presence of commercial detergent but also gave better result of washing with detergent, as compared to only detergent, so it can be used for washing purposes (Table 4).
ACKNOWLEDGMENT
This study is financially supported by Dean Science Project DFS 2005 to Dr. Sh. Ajaz Rasool.
REFERENCES
- Adinarayana, K., P. Ellaiah and D.S. Prasad, 2003. Purification and partial characterization of thermostable serine alkaline protease from a newly isolated Bacillus subtillis PE-11. Pharm. Sci. Technol., 4: E56-E64.
Direct Link - Al-Shehri, M.A. and M.S. Yasser, 2004. Production and some properties of protease produced by Bacillus licheniformis isolated from Tihamet Aseer, Saudi Arabia. Pak. J. Biol. Sci., 7: 1631-1635.
CrossRefDirect Link - Anwar, A. and M. Saleemuddin, 2000. Alkaline protease from Spilosoma obliqua: Potential applications in bio-formulations. Biotechnol. Applied Biochem., 31: 85-89.
CrossRefPubMedDirect Link - Aoyama, M., M. Yasuda, K. Nakachi, N. Kobamoto, H. Oku and F. Kato, 2000. Soybean-milk-coagulating activity of Bacillus pumilus derives from a serine proteinase. Applied Microbiol. Biotechnol., 53: 390-395.
Direct Link - Denizci, A.A, D. Kazan, E.C.A. Abeln and A. Erarslan, 2004. Newly isolated Bacillus clausii GMBAE 42: An alkaline protease producer capable to grow under higly alkaline conditions. J. Applied Microbiol., 96: 320-327.
CrossRefDirect Link - Feng, Y.Y., W.B. Yang, S.L. Ong, J.Y. Hu and W.J. Nig, 2001. Fermentation of starch for enhanced alkaline protease production by constructing an alkalophilic Bacillus pumilus strain. Applied Microbiol. Biotechnol., 57: 153-160.
Direct Link - Fujiwara, N., K. Yamamoto and A. Masui, 1991. Utilization of a thermostable alkaline protease from an alkalophilic thermophile for the recovery of silver from used X-ray film. J. Ferment. Bioeng., 72: 306-308.
Direct Link - Gessesse, A., R. Hatti-Kaul, B. Gashe and B. Mattasson, 2003. Novel alkaline proteases from alkaliphilic bacteria grown using chicken feather. Enz. Microbiol. Technol., 32: 519-524.
CrossRefDirect Link - Gupta, R., K. Gupta, R.K. Saxena and S. Khan, 1999. Bleach stable, alkaline protease from Bacillus sp. Biotechnol. Lett., 21: 135-138.
Direct Link - Gupta, R., Q. Beg, S. Khan and B. Chauhan, 2002. An overview on fermentation, downstream processing of microbial alkaline proteases. Applied Microbiol. Biotechnol., 60: 381-395.
Direct Link - Gusek, T.W. and J.E. Kinsella, 1987. Purification and characterization of the heat stable serine proteinase from Thermomonospora fusca YX. Biochem. J., 246: 511-517.
Direct Link - Hameed, A., M.A. Natt and C.S. Evans, 1996. Production of alkaline protease by a new Bacillus subtilis isolate for use as a bating enzyme in leather treatment. World J. Microbiol. Biotechnol., 12: 289-291.
Direct Link - Horikoshi, K., 1971. Production of alkaline enzymes by alkalophilic microorganisms and alkaline protease produced by Bacillus No. 221. Agric. Biol. Chem., 35: 1407-1414.
Direct Link - Kobayashi, T., Y. Hakamada, S. Adachi, J. Hitomi and T. Yoshimatsue et al., 1995. Purification and properties of an alkaline protease from alkalophilic Bacillus sp. KSM-K16. Applied Microbial. Biotechnol., 43: 473-481.
CrossRef - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Najafi, M.F., D. Deobagkar and D. Deobagkar, 2005. Potential application of protease isolated from Pseudomonas aeruginosa PD100. Electronic J. Biotechnol., 8: 197-203.
Direct Link - Rasool, S.A., S. Ahmed and A. Iqbal, 1996. Streptococcins of indigenous hemolytic streptococci. Nat. Prod. Lett., 08: 67-74.
Direct Link - Rao, M.B., A.M. Tanksale, M.S. Ghatge and V.V. Deshpande, 1998. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev., 62: 597-635.
Direct Link - Schallmey, M., A. Singh and O.P. Ward, 2004. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol., 50: 1-17.
CrossRefPubMedDirect Link - Sharmin, S., M.T. Hossain and M.N. Anwar, 2004. Proteolytic activity of a Lactobacillus species isolated from Rumen. Pak. J. Biol. Sci., 7: 2105-2108.
CrossRefDirect Link - Takami, H., T. Akiba and K. Horikoshi, 1992. Substrate specificity of thermostable alkaline protease from Bacillus sp. No. AH-101. Biosci. Biotechnol. Biochem., 56: 333-3341.
Direct Link - Thangam, E.B. and G.S. Rajkumar, 2002. Purification and characterization of alkaline protease from Alcaligenes faecalis. Biotechnol. Applied Biochem., 35: 149-154.
Direct Link - Yang, J.K., I.L. Shih, Y.M. Tzeng and S.L. Wang, 2000. Production and purification of protease from Bacillus subtilis that can deproteinize crustascean wastes. Enzyme Microbiol. Technol., 26: 406-413.
CrossRef