
A.J. Ajibade
Department of Anatomy, Faculty of Basic Medical Sciences, P.M.B. 4000, Lautech Ogbomoso, Oyo State, Nigeria
P.B. Fakunle
Department of Anatomy, Faculty of Basic Medical Sciences, P.M.B. 4000, Lautech Ogbomoso, Oyo State, Nigeria
P.D. Shallie
Department of Anatomy, Faculty of Basic Medical Sciences, P.M.B. 4000, Lautech Ogbomoso, Oyo State, Nigeria
Current Research in Neuroscience
Year: 2012 | Volume: 2 | Issue: 1 | Page No.: 1-10







ABSTRACT
Several neurological disorders following abuse of artesunate used in chloroquine resistant malarial cases have been reported but little attention has been given the effects of this drug on histology of cerebellar cortex and Nissl substances. Twenty-four adult wistar rats of both sexes weighing between 150 and 230 g were randomly separated into four groups, N = 6. Rats in group D (control), received distilled water throughout exposure period while groups A, B and C (experimental groups) received 4 mg kg-1 b.wt. of artesunate orally on treatment day 1. Group A continued with (4 mg kg-1 b.wt.) for the next 3 days while group B and C received 2 mg kg-1 b.wt. of artesunate orally for the next 6 and 13 days, respectively. Groups A, B and C rats were sacrificed respectively on 5th, 8th and 15th day of treatment while Group D rats were sacrificed on the 15th day. Cerebellum of each rat was carefully dissected out and fixed in 10% formal saline for routine histological techniques. Histological findings showed normal cortical layers in control rats compared with degenerated and loss of purkinje cells, cellular hypertrophy with intercellular vacuolation appearing in the stroma of cerebellar cortex of treated rats. Nissl substances in cerebellar cortex of treated rats stained less intensely and appeared degenerated compared to more intensely stained and distinctly distributed Nissl substances in control rats. The observed vacuolation and neuronal loss in the cortical layers of the treated rats may adversely affect cerebellar functions while less intensely stained and degenerative changes in the Nissl substances in the cerebellar cortex of the treated rats may adversely affect protein synthesis in relation to neuronal functions.
PDF Abstract XML References Citation
Received: February 23, 2011;
Accepted: July 25, 2011;
Published: July 10, 2012
How to cite this article
A.J. Ajibade, P.B. Fakunle and P.D. Shallie, 2012. Some Histological Observations and Microstructural Changes in the Nissl Substances in the Cerebellar Cortex of Adult Wistar Rats following Artesunate Administration. Current Research in Neuroscience, 2: 1-10.
DOI: 10.3923/crn.2012.1.10
URL: https://scialert.net/abstract/?doi=crn.2012.1.10
DOI: 10.3923/crn.2012.1.10
URL: https://scialert.net/abstract/?doi=crn.2012.1.10
INTRODUCTION
Artesunate is an antimalarial drug commonly used as an alternative in chloroquine resistant cases of plasmodum falciparum infection (Nwanjo and Oze, 2007). Artemisinin which is the parent compound, is the antimalarial principle of these compounds and is derived from the leaves of a plant called sweet wormwood (Artemisia annua). Artemisinin was isolated by the Chinese scientists from Artemisia annua leaves (Ngokere et al., 2004). Artesunate is the most widely used member of artemisinin derivatives in the treatment of chloroquine resistant malaria (Woodrow et al., 2005). The effectiveness of artesunate has been attributed to its rapid and extensive hydrolysis to Dihydroartemisinin (DHA) which is three to five fold more active and more toxic, than the parent compound (Li et al., 2002). Many physicians are now using new drugs like artemisinin derivatives, due to very high rates of treatment failure with conventional treatment. Although, number of clinical studies mainly conducted in Africa and Asia, have identified a favorable tolerability profile for these drugs, however, many patients have experienced adverse neurological effects (Price, 2000). Artemisinin derivatives are potent antimalarial drugs, but concern has been raised as to their neurotoxic potential (Li et al., 2006). Several studies have shown that high doses of artesunate can produce neurotoxicity such as selective damage to brain stem centers in mice and rats (Nontprasert et al., 2002). All the adverse effects of artesunate are still not well known, therefore, the potential for neurotoxicity and other form of toxicity of artesunate is an issue that must be studied (Luo et al., 2003). The safety profile of artesiminin derivatives has been questioned because of their potential neurotoxic effects (Santiago et al., 2008). Neurotoxic symptoms were observed in rats with 50 and 100 mg kg-1 from day 6 upward following artemether administration (Oyemitan et al., 2007) Similarly, Obianime and Aprioku (2011) had reported that artesunate and DHA may be toxic to the liver, testis and hematopoietic cells resulting from increase in oxidative stress via stimulation of protein kinase C activity.
Changes in serum glucose and triacylglycerol levels induced by the Co-administration of Two Different Types of Antimalarial Drugs among Some Plasmodium falciparum malarial patients in edo-delta region of Nigeria had been reported by Onyesom and Agho (2011). It has been shown that artesunate and dihydroartemisinin interacted more strongly with Fe (III) PPIX than artemisinin. Although, findings have shown that hemin and endoperoxide lactone derived antimalarials slowly react to give rise to several stereoisomers of artesunate while in contrast, only heme (Fe2) was found to react with artemisinin based drugs in previous studies (Mpiana et al., 2007). Artequin, a combination of artesunate and mefloquine has been reported to be effective against multidrug-resistant Plasmodium falciparum malaria in Nigeria (Agomo et al., 2007). Neurotoxicity has been detected in the various regions of the rat brain following monosodium glutamate administration (Waggas, 2009). Adebayo et al. (2009) also had reported brain damage in rats that were treated with chemical neurotoxins. Long term exposure of rats to electromagnetic field has been shown to be detrimental to the cerebellum (Ozra et al., 2010). The neuropathological effect of Tributyltin (TRB) on the neurons of the cerebral ganglia in snails has been reported by Essawy et al. (2011). Aluminium acetate has similarly, been implicated as a neuro-environmental factor responsible for neurodegenerative diseases. (Sushma et al., 2006).
The cerebellum consists of three distinct layers and contains five major types of neurons (Llinas et al., 2004) The molecular layer is the outermost layer which contains few nerve cells. The purkinje cell layer is the middle layer that contains a monolayer of purkinje cells sandwiched between the molecular and granular layers. The granular layer is the inner layer that consists of densely packed granule cells. Cerebellar injuries have been reported to result from toxins, autoantibodies, structural lesion and inherited cerebellar degeneration (Hain, 2009). In view of the reported adverse neurological effects of artesunate on the central nervous system, this study investigated the microstructural changes in the Nissl substances and the histology of the cerebellar cortex in adult wistar rats following experimental artesunate administration.
MATERIALS AND METHODS
Twenty four wistar rats of both sexes weighting between 150 and 230 g were used for this study (2009). The rats were maintained under standard laboratory conditions. They were fed daily with rat chow purchased from Global Farms Ogbomosho and water was given to the rats ad libitum. The wistar rats were subjected to a period of three weeks of acclimatization before the treatment. The wistar rats were separated into four groups, each contained six rats. Group A, B and C (n =18) served as treatment groups while group D (n = 6) was the control. The rats in treatment groups A, B, C orally record 4 mg kg-1 b.wt. of artesunate base dissolved in distilled water through orogastric tube the first day. Wistar rats in the treatment group A continued with this dosage for the next three days while rats in group B and C received 2 mg kg-1 once daily for the next six and thirteen days, respectively. This was done to investigate some effects of acute, therapeutic and chronic treatment of artesunate on the cerebellum. The control group D received equal volume of distilled water as contained in the experimental doses.
Artesunate tablets were obtained from Salem pharmacy, Ibadan Oyo State, Nigeria. One tablet which contains 50 mg of the active ingredient was dispersed in 50 mL of distilled water following the method of Mesembe et al. (2004).
The treated rats in group A, B and C were sacrificed by cervical dislocation on the 5th, 8th and 15th day of the study respectively, while the control group D was also sacrificed on the 15th day of the study. Respective tissues specimens were processed for routine histological procedures and sectioned at 6μ and then stained for H and E and nissls substances as previously described by Carleton (1967), respectively. Permanent digital microgrpahs of the desired sections were obtained to record morphologic and microstructural observations using a digital camera. The observations were made using a research microscope.
Permanent digital microgrpahs of the desired sections were obtained to record morphologic and microstructural observations using a digital camera. The observations were made using a research microscope.
RESULTS
Histological findings: Figure 1 shows the cerebellar cortex from control rats. The cerebellar cortex of the control rats showed a characteristic appearance of an outer molecular layer and an inner granular layer. The molecular layer contained distinct neurons. The middle cortical layer contained a monolayer of flash shaped purkinje cells sandwiched between the outer molecular and inner granular layers and the three cortical layers appeared normal in Fig. 1.
Figure 2 shows the cerebellar cortex from artesunate-treated rats. The histological section of the cerebellar cortex of the artesunate-treated rats showed some cortical degenerative changes and vacuolations. The purkinje cell layer showed loss and degeneration of purkinje cells which appeared as vacuolations and smaller sized purkinje cells in the purkinje cell layer (Fig. 2).
Figure 3 shows the section of cerebellar cortex from artesunate treated rats that received an initial 4 mg kg-1 b.wt. dose of artesunate followed by a 2 mg kg-1 b.wt. dose of artesunate for 6 days. The purkinje cell layer particularly showed increased degeneration and loss of purkinje cells. Loss of purkinje cells became more pronounced in Fig. 3 compared with Fig. 2.
Figure 4 shows the treated section of cerebellar cortex from artesunate treated rat after receiving an initial 4 mg kg-1 b.wt. dose of artesunate followed by a 2 mg kg-1 b.wt. dose of artesunte for 13 days. There were vacuolations, distortion of cortical layers degeneration and loss of neurons in this section.
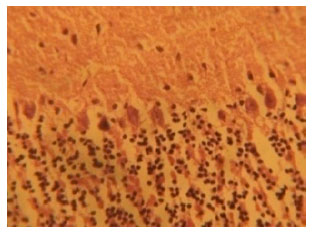 | |
| Fig. 1: | (Group IV): Control section of the cerebellar cortex (Mag. X400) |
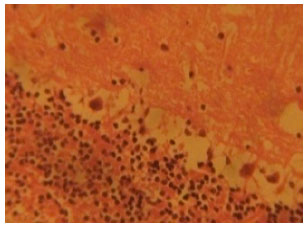 | |
| Fig. 2: | (Group I): Treatment section of the cerebellar cortex that received 4 mg kg-1 of artesunate for 4 days (Mag. X400) |
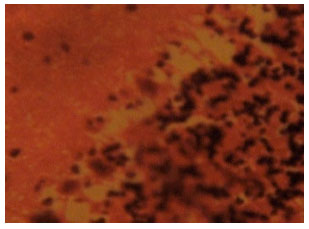 | |
| Fig. 3: | (Group II): Treatment section of the cerebellar cortex that received 4 mg kg-1 1st day and thereafter 2 mg kg-1 for 6 days of artesunate (Mag. X400) |
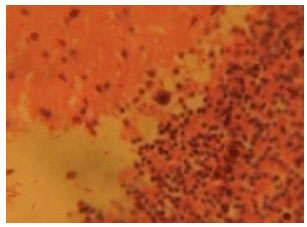 | |
| Fig. 4: | (Group III): Treatment section of the cerebellar cortex that received 4 mg kg-1 1st day and thereafter 2 mg kg-1 for 13 days of artesunate (Mag. X400) |
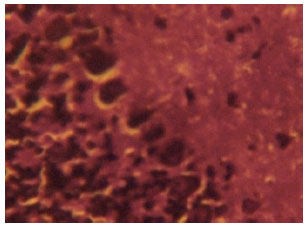 | |
| Fig. 5: | (Group D ): Control section of the cerebellar cortex of wistar rats showing intensely stained Nissl substances in the cortical layers (Mag. X400) |
Increased vacuolations which resulted from loss of cellular components, degeneration and loss of neurons became more evident in this figure. Increased degeneration and loss of purkinje cells became more pronounced in the purkinje cell layer as shown in Fig. 4.
Histochemical findings on nissl substances: The cerebellar cortex of the control rats in group D revealed distinct and intensely stained Nissl substance in the cortical layers. The neurons in the cortical layers appeared normal, distinct and intensely stained (Fig. 5). The purkinje cells are distinct, deeply stained and consist of a monolayer sandwiched between the outer molecular layer and inner granular layer.
The section of the cerebellar cortex from artesunate-treated rats (group A) showed some degenerative changes which made the neurons indistinct. The neurons in the cortical layers appeared to show some degenerative changes which them indistinct with a reduced staining intensity compared with the control section. Loss of neuronal cells and cellular components in the cortical layers are prominent as shown in Fig. 6.
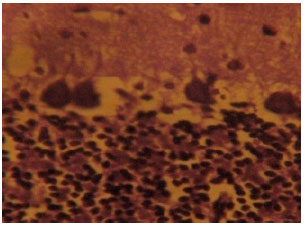 | |
| Fig. 6: | (Group I): Treatment section of the cerebellar cortex that received 4 mg kg-1 of artesunate for 4 days showing less intensely stained and degenerated Nissl substances in the cortical layers Cresyl Violet (Mag. X400) |
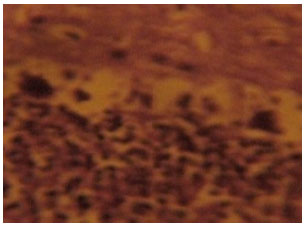 | |
| Fig. 7: | (Group B): Treatment section of the cerebellar cortex that received 4 mg kg-1 1st day and thereafter 2 mg kg-1 for 6 days of artesunate showing less intensely stained and degenerated Nissl substances in the cortical layers Cresyl Violet (Mag. X400) |
Section of the cerebellar cortex from group B treated rats also showed degenerative changes which made the neurons in the three cortical layers indistinct with reduced staining intensity compared with intensely stained and distinct neurons in the cortical layers of the control rats. The Nissl substances in the cortical layers as shown in Fig. 7 appeared degenerative with loss of staining intensity that made the neuronal cells indistinct. Some neurons particularly the purkinje cells in the middle layer appeared to have been lost in this section.
Section of the cerebellar cortex from group C artesunate-treated rats also revealed some degenerative changes in the three cortical layers. The neurons in the three cortical cell layers of Fig. 8 appeared degenerative which made them indistinct with remarkable loss of staining intensity in the three cortical layers compared with the control section. Complete loss of purkinje cells and distortion of cortical layers became prominent in Fig. 8 as shown in this section.
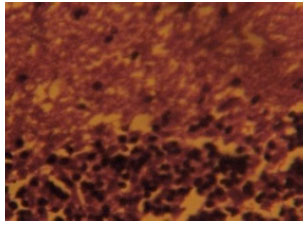 | |
| Fig. 8: | (Group C): Treatment section of the cerebellar cortex that received 4 mg kg-1 1st day and thereafter 2 mg kg-1 for 13 days of artesunate showing less intensely stained and degenerated Nissl substances in the cortical layers Cresyl Violet (Mag. X400) |
DISCUSSION
Histological findings: The result of the haematoxylin and eosin stain showed degenerative changes and loss of cellular components with reduced population of purkinje cells in the treatment groups compared with the control section of the cerebellar cortex of the adult wistar rats. Cells death has been reported to result from neuronal degeneration (Waters et al., 1994). Cell death may result from necrosis or apoptosis. Apoptosis and necrosis differ morphologically and biochemically. Necrosis is a pathological change which may result from toxic, thermal, traumatic and mechanical factors while apoptosis is an organized form self destruction also known as programme cell death that is mediated by intrinsic and active mechanisms (Wyllie et al., 1980). Pathological or accidental cell death is regarded as necrotic and could result from extrinsic insults to the cells such as osmotic, thermal, toxic and traumatic effect (Faber et al., 1981). Cell death in response to neurotoxin might trigger an apoptotic death pathway within brain cells.
Artesunate crosses membrane and affects the cellular integrity of tissue. Artesunate in this study, possibly might have acted as a neurotoxin to the cerebellar cortex thereby affecting neuronal integrity and causing disruption in membrane permeability and tissue homeostasis.. The degenerative effect of artesunate on the cortical layers of the cerebellum observed in this study may be responsible for the cerebellar degeneration. Clinical studies have reported adverse effects with artemisinin derivatives when used alone or in combination with other antimalarials in therapeutic doses. Adverse neurological effects after treatment include acute psychosis, depressant syndrome, sleep disturbance (McIntosh and Olliaro, 2000) and post-cerebellar syndrome-ataxia and slurred speech (Sabchareon et al., 1998). It is not impossible that artesunate had irreversible neurotoxic effect on the cortical layer of the cerebellum which resulted in distortion, loss of cellular component and purkinje cells in the cortical layers of the treated cerebellar section. These findings agree with the earlier report of Ajibade et al. (2006) on cerebellar cortex of wistar rats following quinine administration. The degenerative changes observed in this study may underlie the possible neurological symptoms such as ataxia, tremor slurred speech and gait disturbance following artesunate administration which has been reported Miller and Panosian (1997).
Histochemical findings on nissl substances: The sites in the cytoplasm of basophilic ribosomes stain intensely with such basic dyes as methylene blue, toludine blue and haematoxyhin. These basophilic regions have been described as early as 19th century and were named according to the cells being studied. They were called Nissl bodies in neurons. When appropriate stains are used Rough Endoplasmic Reticulum (R.E.R) and free ribosomes appear under the light microscope as basophilic granular areas called Nissl bodies. The purkinje cells appeared distinct when nissl substances were stained which is consistent with the finding of Gharravi et al. (2007). Neuronal degeneration has been reported to cause a reduction in Nissl boches (Martin et al., 1998). The observed neuronal degeneration in association with loss of Nissl bodies with resultant reduced staining intensity of the Nissl substances in the cerebellar cortex of the artesunate-treated rats in this study is consistent with findings of Ajibade et al. (2009).
Injury to axons or neuronal exhaustion resulting from strong or prolong stimuli causes a reduction in the number of Nissl bodies. This alteration which is called chromatolysis which occurs simultaneously with nuclear migration to the periphery of the perikaryon and consequently the RNA level is reduced (Louis et al., 1986) Chemical and toxic substances affect the Nissl substances thereby influencing their metabolic activity (Davis and Robertson, 1991). The findings from this study agreed with degeneration and vacuolar changes which have been observed in many large brain stem neurons in mice that were treated with trimethyl chloride. These neurons acquired a chromatolytic character with eccentric nuclei and loss of Nissl substances which became progressive in ultra structure (Change et al., 1983). Similarly, research findings have shown that fatigue from over exertion; produced in the brain cells similar to those changes produced by fear resulted in exhaustion and consequently enormous reduction in Nissl substance. These changes were proportional to the amount of exertion which is in agreement with observation on nissl substances in this study. Loss of Nissl substance observed in this study is also supported by the earlier findings of Classen et al. (1999) reporting loss of Nissl substance in neurons and shrinkage of nucleus which became prominent in the cerebellar roof, pontine and vestibular nuclei of dog following intramuscular administration of artemether.
CONCLUSION
This study concluded that chronic administration and high doses of artesunate administered to adult wistar rats resulted in loss of cellular components, distortion of the cortical layers and loss of Nissl substances in the cerebellar cortical layers. These pathological changes may impair cerebellar functions.
ACKNOWLEDGMENT
The authors wish to express their gratitude to the technical staff of the Anatomy Department Lautech, Ogbomoso, Nigeria, for their support in the execution of this present work.
REFERENCES
- Waggas, A.M., 2009. Neuroprotective evaluation of extract of ginger (Zingiber officinale) root in monosodium glutamate-induced toxicity in different brain areas of male albino rats. Pak. J. Biol. Sci., 12: 201-212.
CrossRefPubMedDirect Link - Adebayo, O.L., B.O. Adegbesan and G.A. Adenuga, 2009. Comparison of the effects of low protein diet versus chemical neurotoxins on brain weight, brain lipid peroxidation and antioxidant status of rats. Asian J. Biol. Sci., 2: 7-13.
CrossRefDirect Link - Agomo, P.U., R.A.S. Mustapha, B.G. Omoloye, A.N. Okechukwu and A.G. Mafe et al., 2007. Efficacy and safety of artesunate +mefloquine (Artequin) in the treatment of uncomplicated Falciparum malaria in ijede community, ikorodu LGA, Lagos State, Nigeria. J. Med. Sci., 7: 816-824.
CrossRefDirect Link - Essawy, A.E., S.S. El-Sherief, I.A. Sadek and A.A. Soffar, 2011. Neuropathological Effect of Tributyltin on the Cerebral Ganglia of the Land Snail, Eobania vermiculata Int. J. Zool. Res., 7: 252-262.
Direct Link - Ajibade, A.J. O.A. Adeeyo, D.A. Olusori, T.K. Adenowo, O.O. Ishola, E.A. Ashamu and S.C. Nwangwu, 2009. Microstructural observations on nissl substances in the cerebellar cortex of adult Wistar rats following quinine administration. Trop. J. Pharm. Res., 8: 105-109.
Direct Link - Carleton, H.M., R.A.B. Drury, 1967. Carleton's Histological Technique. 4th Edn., Oxford University Press, Oxford, England, ISBN: 9780192612052, Pages: 432.
Direct Link - Classen, W., B. Attman, P. Gretener, C. Souppart, P. Skelton-Stroud, G. Kirinke, 1999. Differential effects of orally versus parentally administered qinghaosu derivative artemether in dogs. Proccedigns of 7th International Neurotoxicology Association Meeting: Leicester, (INAM'99), International Neurotoxicology Association INAT Abs, UK., pp: 1-47.
- Faber, J.L., K.R. Chien and S. Mittnacht Jr., 1981. Myocardial ischemia: The pathogenesis of irreversible cell injury in ischemia. Am. J. Pathol., 102: 271-281.
Direct Link - Gharravi, A.M., M.J. Golalipour, R. Ghorbani and M. Khazaei, 2007. Effects modification of iron hematoxylin on neuron staining. Pak. J. Biol. Sci., 10: 768-772.
CrossRefPubMedDirect Link - Li, Q.G., S.R. Mog, Y.Z. Si, D.E. Kyle, M. Gettayacamin and W.K. Milhous, 2002. Neurotoxicity and efficacy of arteether related to its exposure times and exposure levels in rodents. Am. J. Trop. Med. Hyg., 66: 516-525.
PubMed - Louis, E., J.A. Verges and F. Flores, 1986. Effect of disordered concentrated nonmagnetic impurities on the periodic Anderson Hamiltonian in one dimension. Phys. Rev. B., 34: 6415-6419.
PubMed - Luo, S.E., H.J. Zhuo and H.B. Huang, 2003. Lipid-peroxidantion damage of embryo and placenta induced by artesunate in rats. Zhejiang Da Xue Xue Bao Yi Xue Ban., 32: 41-45.
PubMed - Mesembe, O.E., A.E. Ivang, G. Udo-Affah, A.O. Igiri and V.A. Fisher et al., 2004. Teratogenic effect of artesunate on the histology of developing spinal cord of wistar rat fetuses. J. Biomed-Afr., 11: 62-64.
Direct Link - Miller, L.G. and C.B. Panosian, 1997. Ataxia and slurred speech after artesunate treatment for falciparum malaria. N. Eng. J. Med., 336: 1328-1329.
PubMedDirect Link - Mpiana, P.T., B.K. Mavakala and Y. Zhi-Wu, 2007. Interaction of artemisinin based antimalarial drugs with hemin in water-DMSO mixture. Int. J. Pharmacol., 3: 302-310.
CrossRefDirect Link - Nontprasert, A., S. Pukrittayakamee, A.M. Dondorp, R. Clemens, S. Looareesuvan and N.J. White, 2002. Neuropathologic toxicity of artemisinin derivatives in a mouse model. Am. J. Trop. Med. Hyg., 67: 423-429.
Direct Link - Nwanjo, H. and G. Oze, 2007. Acute hepatotocixity following administration of artesunate in guinea pigs. Internet J. Toxicol. Vol. 4.
Direct Link - Obianime, A.W. and J.S. Aprioku, 2011. Mechanism of action of artemisinins on biochemical, hematological and reproductive parameters in male guinea pigs. Int. J. Pharmacol., 7: 84-95.
CrossRefDirect Link - Oyemitan, I.A., E.O. Iwalewa, O.E. Ukponmwan, R.O. Akomolafe and O.M. Daniyan, 2007. The involvement of serotonin in artemether-induced behavioural activities. J. Biol. Sci., 7: 575-578.
CrossRefDirect Link - Onyesom, I. and J.E. Agho, 2011. Changes in serum glucose and triacylglycerol levels induced by the co-administration of two different types of antimalarial drugs among some Plasmodium falciparum malarial patients in Edo-delta Region of Nigeria. Asian J. Sci. Res., 4: 78-83.
CrossRefDirect Link - Ozra, A., S.R. Jafar, L. Moradi and G. Saki, 2010. Ultrastructural change of cerebellum in exposed rats to 3mT electromagnetic field. J. Biol. Sci., 10: 526-530.
CrossRefDirect Link - Price, R.N., 2000. Artemisinin drugs: Novel antimalarial agents. Expert Opin. Invest. Drugs, 9: 1815-1827.
CrossRef - Li, Q., L.H. Xie, A. Haeberle, J. Zhang and P. Weina, 2006. The evaluation of radiolabeled artesunate on tissue distribution in rats and protein binding in humans. Am. J. Trop. Med. Hyg., 75: 817-826.
PubMed - Santiago, C., P. de la Cerda and A. Rivera, 2008. Fatal artesunate toxicity in a child. J. Pedeatr. Inf. Dis., 1: 67-75.
Direct Link - Sushma, N.J., U. Sivaiah, N.J. Suraj and K.J. Rao, 2006. Aluminium acetate induced oxidative stress in brain of albino mice. J. Pharmacol. Toxicol., 1: 579-584.
CrossRefDirect Link - Waters, C.M., W. Moser, G. Walkinshaw and I.J. Mitchell, 1994. Death of neurons in the neonatal rodent and primate globus pallidus occurs by a mechanism of apoptosis. Neuroscience, 63: 881-894.
CrossRef - Woodrow, C.J., R.K. Haynes and S. Krishna, 2005. Artemisinins: Mechanism of action. Postgrad. Med. J., 81: 71-78.
Direct Link - Wyllie, A.H., J.F. Kerr and A.R. Currie, 1980. Cell death, the significance of apoptosis. Int. Rev. Cytol., 68: 251-305.
PubMed