
H. Sekena Abd EL-Aziem
Department of Cell Biology, National Research Center, Dokki, Cairo, Egypt
A. Zeinab Saleh
Department of Food Science and Nutrition, National Research Center, Dokki, Cairo, Egypt
A. F. Farrag
Department of Dairy, National Research Center, Dokki, Cairo, Egypt
International Journal of Dairy Science
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 126-137







ABSTRACT
The purpose of this study is to investigate the possible antimutagenic effects of whey proteins against genotoxic effects induced by aflatoxins in the mammalian cell system. The micronuclei, chromosomal abnormalities in both somatic and germ cells as well as DNA titrations in male rats were assayed. Whey proteins was added to the diet containing aflatoxins (2.5 mg AFs kg-1) and fed to mature male rats for 9 weeks. The results indicated that aflatoxin significantly (p<0.001) increased chromosomal abnormalities in somatic and germ cells, DNA titration and micronuclei of bone marrow cells. Addition of whey proteins to the aflatoxins contaminated diet resulted a significant decrease in chromosomal aberrations in somatic cells (62.8%, p<0.001), in germ cells (57.9%, p<0.01) and in micronuclei polychromatic erythrocytes (68.2%, p<0.001). It could be concluded that whey protein reduced the mutagenesis and clastogenesis effects of AFs. Although the mechanism of antimutagenicity is unknown, it seems plausible that the whey proteins acts as a blocking agent by chemical or physical interaction with the mutagens. Furthermore, this study revealed that the addition of whey proteins to aflatoxin contaminated diet diminished most of the genotoxic effects of the Aflatoxin.
PDF Abstract XML References
How to cite this article
H. Sekena Abd EL-Aziem, A. Zeinab Saleh and A. F. Farrag, 2007. Impact of Whey Proteins on the Genotoxic Effects of Aflatoxins in Rats. International Journal of Dairy Science, 2: 126-137.
DOI: 10.3923/ijds.2007.126.137
URL: https://scialert.net/abstract/?doi=ijds.2007.126.137
DOI: 10.3923/ijds.2007.126.137
URL: https://scialert.net/abstract/?doi=ijds.2007.126.137
INTRODUCTION
Whey Proteins (WP) is a major component of the protein fraction in bovine milk. Milk contains 5.5 g WP/L and the major protein fractions are β-lactoglobulin (58%) α-lactalbumin (20%) immunoglobuline (13%) and serum albumin (7%) (Ito, 1991). Whey contain a wide range of biologically active proteins, i.e., about 60 indigenous enzymes, vitamin-binding protein, metal-binding protein, immunoglobulins, various growth factors and hormones, these components have been reviewed by IDF (1991), Smithers et al. (1996) and Parodi (1998). In previous studies, a total protein diet has been shown to have immunostimulatory and ant carcinogenic effects in mice and rats (McIntosh et al., 1995). The possible ant mutagenic effects of five different proteins (the proteins casein, Bovine Serum Albumin (BSA), soy protein, total whey proteins and β-lactoglobulin) against the mutagen 4-nitroquinoline 1-oxide (4NQO) were assessed in mammalian cell system, using the Sister Chromatid Exchange (SCE) test in Chinese hamster cells (V79). They showed that BSA significantly inhibited 4NQO-induced SCEs (94%, at 1.15% BSA; p<0.01%). However, soy protein, the whey protein fraction of milk and β-lactoglobulin showed no inhibitory effects. Pepsin-hydrolysed casein inhibited SCE induction by 4NQO and 1-methyl-nitroso-3-nitroguanidine (MNNG) to a similar extent as non-hydrolysed casein (Bosselaers et al., 1994). Whey proteins are also used as a food supplement for immune compromised individuals for the prevention of diet-related cancers (Parodi, 1998).
Numerous studies have shown that whey proteins, which are high in the amino acid cysteine, help to enhance the body’s immune system by raising glutathione levels (Bounous, 2000). Kent et al. (2003) indicated that whey proteins can increase GSH synthesis and protect against oxidant-induced cell death in human prostate cells.
Aflatoxins B1 (AFB1) is among the most common mycotoxins to which human are exposed. Experimental studies have shown that AFB1 is a potent mutagen and hepatocarcinogen (Nixon et al., 1981; Hsieh and Atkinson, 1991). A number of epidemiological investigations have also implicated its role in the etiology of human liver cancer (Van Rensburg et al., 1985; Yeh et al., 1989). A study with cultured human lymphocytes suggested that AFBI was able to induce chromosomal damage through the release of Reactive Oxygen Species (ROS) (Amstad et al., 1984).
Accumulation of intracellular ROS during extended period of oxidative stress is associated with the development of many chronic diseases (Devaux et al., 2001; Nelson et al., 2001). Glutathione (γ-glutamyl-cysteinyl-glycine, GSH) is the most abundant non protein thiol in mammalian cells and functions as an antioxidant to limit oxidant induced damage to lipids, proteins and genetic material (Sen, 1997; Anderson, 1998; Griffith, 1999; Lu, 1999). Depletion of intracellular GSH can lead to the accumulation of intracellular ROS (Esteve et al., 1999; Teramoto et al., 1999). Consequently, maintaining a high intracellular concentration of GSH is therefore critical for cellular defence against oxidative stress.
GSH, in its reduced form, can donate its sulfhydryl proton to quench ROS. Once oxidized, GSH forms a disulfide linkage with a second molecule of oxidized GSH, yielding glutathione disulfide (GSSG). Maintaining a high intracellular GSH: GSSG ratio provides optimal protection against oxidant-induced cell damage (Anderson, 1998).
Diets that are rich in cystine-containing proteins can elevate tissue GSH (Bounous, 2000; Taniguchi et al., 2000). Cystine is the disulfide form of cysteine and is reduced to 2 moles of cysteine for use in cellular GSH synthesis (Anderson, 1998). Bovine whey proteins a product of cheese manufacture, are a cystine-rich protein source (Morr and Ha, 1993). Consumption of cystine-rich whey proteins can increase plasma GSH concentrations in humans (Kennedy et al., 1995a; Micke et al., 2001) and also reduce the incidence of mammary and colon tumors in rats (Baruchel and Viau, 1996; Sekine et al., 1997; Bounous, 2000; Tsuda et al., 2000; Hakkak et al., 2000).
The present study was conducted to determine the ability of whey protein concentrate to reduce the mutagenic effects due to consumption of aflatoxins contaminated diet in male Sprague Dawley rats.
MATERIALS AND METHODS
Experimental Animals
Twenty-eight male, Sprague-Dawley rats (average weight 153 g), obtained from the animal house of the National Research Center, Dokki, Giza. Animals were housed individually in stainless steel cages and kept through the experimental period under good ventilation and hygienic conditions, water and food were available.
Preparation of Rat Diets
A slightly modified AIN-93M semi-purified diet (recommended by the American Institute of Nutrition (Reeves et al., 1993) was used for control in the present study (Table 1). Vitamins and minerals mix were prepared according to Bieri et al. (1977). The same diet was given to the experimental group but after replacement 50% of casein with Whey Protein Concentrate (WPC).
Preparation of Whey Protien Concentrates (WPC)
Sweet whey from Ras cheese manufacture (pH 6.0-6.2) was obtained from Arab Dairy Co., Kaha, Kaluibia, Egypt. Fat and curd fine were removed from the whey by a cream separator. Whey content, 7.5% total solids, 0.75% total protein and 4.5% Lactose.
| Table 1: | Composition of the diet* |
 | |
| *Vitamins and mineral mix. (AIN- 93, American Institute of Nutrition, 1993) | |
| Table 2: | Gross composition of whey protein concentrates (WPC) |
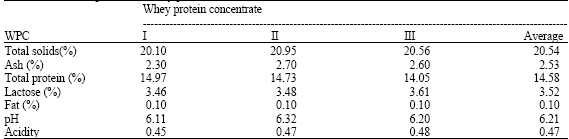 | |
Ultrafiltration of Ras cheese whey was carried out using a carbosep 151 UF system (Orelis, France) equipped with zirconium oxide membrane (Mol. cut off 50,000). Ultrafiltration was carried out in batch mode at 50°C and inlet and outlet pressure of 6 and 4 bar, respectively. Ultrafiltration was continued until to the concentration factor of 20. The gross Chemical composition of whey protein concentrate was determined, TS according to AOAC (1990); Total protein, fat, ash and pH according to Ling (1963); lactose according to Barnett and Tawab (1957). The data is presented in (Table 2).
Aflatoxins
Aflatoxins (AFs) were produced by the method of (Davis et al., 1966). Liquid medium, Yeast Extract Sucrose (YES) inoculated with Aspergillus parasiticus NRRL 3145 and incubated at 25°C for 13 days. At the end of the incubation period, liquid culture was filtered and extracted according to the method of (Park and Bullerman, 1983). The mates were dried at 80°C overnight and extracted by the method of (Shih and Marth, 1971). The Aflatoxins concentration of the crude extract (B1, 2, G1 and 2) were determined and calculated according to (Shannon et al., 1983). The calculation was made relative to the LD50 of the aflatoxin B1. The AFs were thoroughly mixed with the diet to provide the desired level of 2.5 mg/kg diet (Saleh et al., 2000).
Experimental Design
Animals were randomized by weight into four groups (7 rats/group) as follow:-
| Group 1 | : | Control fed on the basal diet. |
| Group 2 | : | Rats were fed AFs-contaminated diet. |
| Group 3 | : | Rats were fed on AFs-contaminated diet after replacement 50% of casein with whey proteins. |
| Group 4 | : | Rats were fed on basal diet after replacement 50% of casein with whey proteins. |
To avoid the fermentation of diets due to addition of whey protein concentrate, fresh diets were prepared every day.
The feeding period lasted for 9 weeks during which rats were weighed and food intake was determined weekly.
At the end of the experimental period, animals were killed and bone marrows were removed for micronucleus analysis, slides were prepared according to Salamone et al (1980). The bone marrow cells were mixed with one drop of fetal calf serum and smeared on clean slides. The slides were fixed in absolute ethyl alcohol for five minutes and then air dried, stained with Giemsa and mounted with DPX. Two thousands PCEs/animal were recorded. Micronuclei were identified as dark blue staining bodies in the cytoplasm of polychromatic erythrocytes (PCEs).
Mitotic chromosomes were prepared and slides were stained with Giemsa in phosphate buffer pH 6.8 according to the method described by Preston et al (1987). One hundred metaphase spreads were checked and the types and frequency of aberrations were recorded and photographed. The testis was removed and meiotic chromosomal slides were prepared according to Russo (2000). Seventy five metaphase spreads were examined in spermatocytes for each animal.
DNA Extraction and Determination
Extraction and determination of DNA was carried out according to method reported by Schneider (1957).
Statistical Analysis
Statistical analysis were carried out using Statistical Programs (SPSS, Version 7.5). Arithmetic mean values and standard errors were computed. Student's t-test was used for statistical evaluation. All statements of significant were based on a probability of p<0.05 or higher.
RESULTS
Cytogenetic Results
Micronuclei in Polychromatic Erythrocytes
It is clearly indicated that the mean values of micronuclei were increased significantly in the groups treated with aflatoxins (p<0.001). The pretreatment with whey proteins significantly reduced (p<0.01) the frequencies of Mn-PCEs to reached 68.2%, although this inhibition in Mn-PCEs were still significantly (p<0.05) higher than control (Table 3 and Fig. 1).
DNA
Treatment with aflatoxins resulted in a significant increase (p<0.001) in the quantity of DNA compared with the control. Pretreatment with whey proteins significantly reduced (p<0.05) the quantities of DNA (Table 4 and Fig. 2).
| Table 3: | Mean±SE of induced micronuclei in polychromatic erythrocytes (MN-PCEs) of rats |
 | |
| *p≤0.05, **p≤0.01, ***p≤0.001 | |
| Table 4: | Mean±SE of DNA concentrations (mg g-1 tissue) in liver of rats induced by aflatoxins |
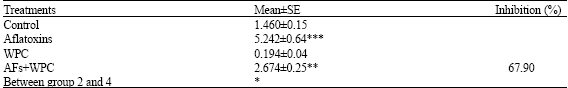 | |
| *p≤0.05, **p≤0.01, ***p≤0.001 | |
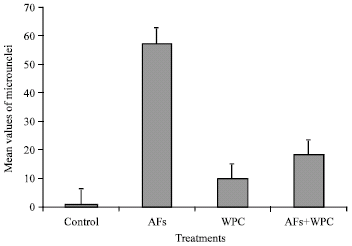 | |
| Fig. 1: | Mean values of induced micronuclei in polychromatic erythrocytes of rats |
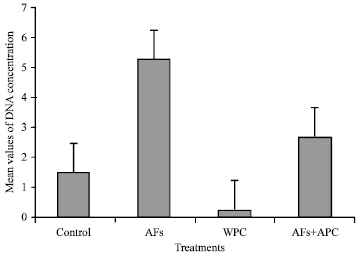 | |
| Fig. 2: | Mean values of measured DNA concentrations (mg g-1 tissue) in liver of rats |
Chromosome Aberrations in Somatic Cells
It shows that aflatoxins induced different structural abnormalities in chromosomes such as gaps, breaks, deletions, centromeric attenuations, end mitosis, end to end associatons and fragments. Numerical aberrations were represented by polyploidy (Table 5). The total chromosomal aberrations were increased significantly in all treated groups than the control. Table 5 showed that whey proteins simultaneously with aflatoxins. It decreased the percentages of the total chromosomal aberrations at all tested aberrations induced by aflatoxins and modulated chromosomal aberrations towards the normal value of the control.
Rats fed diet supplemented with whey proteins showed a few chromosomal aberrations compared with either aflatoxins or control groups. However, some aberrations such as deletions, end to end associations, breaks, fragments, gaps and polyploidy were slightly higher in whey proteins group than those in the control one, but these increase were not significantly.
Statistical analysis showed that there were a significant differences (p<0.001) between aflatoxins and control groups for frequencies of total structural aberrations in all types of aberrations, significant (p<0.01) difference in each of end mitosis, deletions and polyploidy as well as significant difference (p<0.05) in rings stat. Pretreatment with whey proteins significantly (p<0.001) reduced the total structural aberrations induced by aflatoxins by nearly 62.8%, but the total aberrations were still significantly (p<0.01) higher than control (Fig. 3).
| Table 5: | Mean±SE different chromosomal aberrations induced by aflatoxins in bone marrow of male rats |
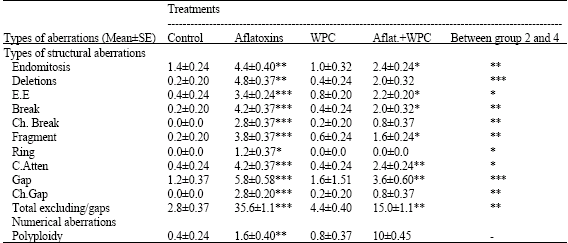 | |
| *p<0.05, **p<0.01, ***p<0.001, C.F = Centric fusion, EE = End to end associations total, ex. g. = Total excluding gaps, Ch. Break = Chromosomal break, C.atten = Centromeric attenuation, Ch. Gap = Chromosomal gap | |
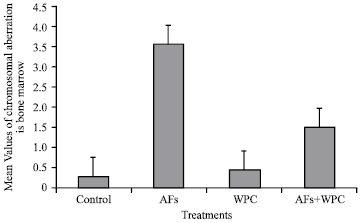 | |
| Fig. 3: | Mean±SE of different chromosomal aberrations in bone marrow of male rats |
Chromosomal Aberrations in Germ Cells
Chromosome examination in spermatocytes (Table 6) showed X-Y univalent, autosomal univalent, chain, rings and ploidy (Fig. 4) revealed that aflatoxins group had more frequencies of chromosomal abnormalities than the control and the other groups. Feeding rats with whey proteins decreased the number of these aberrations. The whey proteins group had few chromosome aberrations compared with either control or aflatoxins group. Statistical analysis showed that there were significant differences (p<0.001) in the frequencies of total structural aberrations in aflatoxins group compared to the negative control. Feeding rats with whey proteins significantly (p<0.001) reduced the total structural aberrations induced by aflatoxins by nearly 57.9%, but the total aberrations were still significantly (p<0.01) higher than control.
DISCUSSION
Bone marrow cells are susceptible to oxidative damage and sensitive to clastogenic chemicals so, it commonly used models for the screening for mutagenicity and/or antimutagenicity (Umegaki et al., 1997).
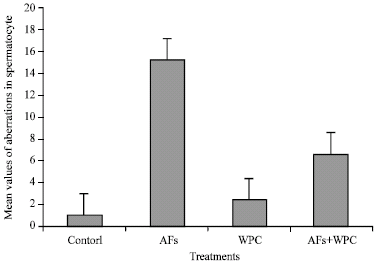 | |
| Fig. 4: | Mean values of different aberrations induced by aflatoxins in spermatocyte of rats |
| Table 6: | Mean±SE of different aberrations induced by aflatoxins in spermatocyte of rats |
 | |
| *p≤0.05, **p≤0.01, ***p≤0.001 | |
The clastogenic action of the above dose of aflatoxins in somatic cells has already been reported for mice and rats bone marrow cells (Krishnamurthy and Neelaram, 1986). AFB1 is a mutagen in Saccharomyces cerevisiae (Sengstag et al., 1996), Escherichia coli, rainbow trout, mice, rat and human cells (Smela et al., 2001).
The current study indicated that AFs induced micronuclei in polychromatic erythrocytes in bone marrow cells of rats. Similar to these results Anwar et al., (1994) and Bárta et al. (1990) observed an increase in the number of polychromatic erythrocytes with micronuclei in the bone marrow cells of mice treated with Aflatoxins.
The present study revealed that AFs increases significantly all types of chromosomal aberrations in somatic cells of rats. In the study of Anwar et al. (1994) chromosomal aberrations were significantly increased in rats treated with AFs compared to the control group at doses above 0.1 μg g-1. Moreover, Bose and Sinha (1991) found that AFs induced chromosomal abnormalities in the bone marrow cells of mice. Salassidis et al.(1991) found that aflatoxin B1 (AFB1) induced structural Chromosomal Aberrations (CA) and Sister Chromatid Exchange (SCE) in rat and mouse hepatocyte.
In the present study, described the role of whey proteins in protecting against the toxic and mutagenic effects of AFs. Whey protein inhibited AFs induced micronuclei by about (68.2%), DNA (67.90%) and chromosomal aberrations (57.9%). Similar results were reported by Bartfay et al. (2003). Mice receiving iron treatments with whey proteins supplementation had significantly lowered concentrations of cytotoxic aldehydes and significantly higher cardiac levels of GPX and GSH activity than did iron-only treated mice. The antimutagenic effects of whey, acetone extracts and protein fractions isolated from milk that had been fermented by L. helveticus L89 were investigated using the mutagen 4-nitroquinoline-N¢-oxide in the Ames test S. typhimurium TA 100. Fermented milk significantly inhibited mutagenesis induced by 4-nitroquinoline-N¢-oxide. (Matar et al., 1997).
Previous reports Fernandes and Shahani (1990), Gilliland (1990) and Goldin (1989) suggested that fermented dairy products exhibit antimutagenic and antitumor activities. Antimutagenic activity of fermented milk has been demonstrated in vitro against a large spectrum of mutagens. Hosono et al. (1988) Bodana and Rao (1990) and Hosoda et al. (1992) and Nadathur et al. (1994, 1995) reported a significant antimutagenic effect of milk against 4-nitroquinoline-N¢-oxide (4NQO) and Moreover, Cassand et al. (1994) reported an inhibitory effect of milk that had been cultured with Bifidobacterium species or Lactobacillus helveticus on mutagenesis induced by 4NQO, 2-nitrofluorene, quercetin and benzopyrene.
Van Boekel et al (1993) demonstrated that casein hydrolysis by pepsin increased inhibitory activity against several mutagens and that the UF permeate from the casein hydrolysates was also antimutagenic. Peptides that were released by pepsin appeared to be even more antimutagenic than casein itself, leading to conclude that antimutagenicity was due to the peptides formed and that limited proteolysis increased the number of sites available for reaction with mutagens (Van Boekel et al., 1993). The inhibitory effect of casein and its pepsin hydrolysate against 4NQO and MNNG has been demonstrated in a mammalian cell culture system (Bosselaers et al., 1994). Because antimutagenic activity has been shown to be greater in fermented milk (Bodana and Rao, 1990; Hosono et al., 1988; Nadathur et al., 1994) and casein hydrolysates (Jongen et al., 1987; Van Boekel et al., 1993) than in unfermented milk and native casein. Furthermore, the detection of inhibitory activity in the whey protein fraction indicates a probable association with antimutagenic compounds of smaller molecular mass, such as peptides. Moreover, peptides resulting from the attack of casein by the combined action of rennet and lactic acid bacteria could have been released in cheese whey protein and have been responsible for this antimutagenic activity of the cheese whey proteins fraction.
The mechanisms by which whey proteins could alter carcinogenesis are unknown. Speculation has focused primarily on increases in tissue GSH levels observed during whey consumption (Bounous et al., 1991). Greater GSH concentrations would tend to be protective because of: (a) the well-known xenobiotic detoxification pathway involving GSH and glutathione S-transferases; (b) GSH free radical detoxification and (c) improved immune responses (Bounous et al., 1989). Other possible mechanisms could include bio-active peptides that are either fragments of whey hydrolysis or contaminants of whey proteins, such as insulin-like growth factor I, which could act on one of many cellular processes to reduce tumor incidence. Studies are currently underway in our laboratory to determine the mechanisms by which whey proteins consumption prevents DMBA-induced breast cancer.
Whey proteins may play a role in both protecting against cancer (Harper, 2000) and also in sensitizing cells to chemotherapy (Tsai et al., 2000). GSH is a major contributor to these observations. Tumor cells contain higher levels of GSH than normal cell tissues. Studies have suggested that an elevated GSH concentration in tumor cells is a useful predictor of resistance or reduced sensitivity to anticancer treatments (Bounous, 2000). In vitro and animal studies has demonstrated a positive correlation between an ingestion of whey proteins and a reduction of intra-cellular GSH activity in cancer cells, a direct contrast to the effect seen in HIV/AIDs patients. Whey proteins are rich in cysteine as well as in GSH precursor peptides. The semi-essential amino acid cysteine is the main source of the free sulfhydryl group of GSH and limits its synthesis (Micke et al., 2002). Results of a 1995 study involving five patients with metastatic carcinoma of the breast and one with metastatic carcinoma of the liver indicated that ingestion of whey proteins might deplete intracellular concentrations of GSH and render them more sensitive to chemotherapy (Kennedy et al., 1995b). A study, conducted by Tsai et al., (2000) demonstrated that a Whey Protein Isolate (WPI) had an enhancing effect on the cytotoxicity of baicalein, a potential anticancer drug, when applied to a human cell line. The whey protein-fed rats that developed mammary tumors had fewer and smaller tumors than the control rats. These observations may further validate the suggested role of whey proteins in tumor prevention or regression. Hakkak et al. (2000) suggest that whey proteins may be one of the most potent dietary sources of cancer prevention identified to date.
The mechanism of antimutagenicity remains speculative. Inhibition might be due either to physical binding by entrapping of the mutagen by the casein micelle or adsorption of the mutagen on the protein molecule, thus preventing its reaction with the target cell. A quenching reaction may occur between the mutagen and the peptides that were derived.
This study revealed that the addition of Whey proteins to aflatoxin contaminated diet diminished most of the deleterious effects of the aflatoxin.
ACKNOWLEDGMENT
The present study was made available through a project contact on (Separation and utilization of whey protein in manufacture functional dairy products) financed by the National Research Center. The authors are greatly indebted to Professor, Safinaz El-Shiking, the projects’s principal investigator for providing facilities and exultant support.
REFERENCES
- Amstad, P., A. Levy, L. Emerit and P. Cerutti, 1984. Evidence for membrane-mediated chromosomal damage by AFB1 in human lymphocytes. Carcinogenesis, 5: 719-723.
PubMedDirect Link - Anderson, M.E., 1998. Glutathione: An overview of biosynthesis and modulation. Chem. Biol. Interac., 111-112: 1-14.
CrossRefDirect Link - Anwar, W.A., M.M. Khalil and C.P. Wild, 1994. Micronuclei, chromosomal aberrations and aflatoxin-albumin adducts in experimental animals after exposure to aflatoxin B1. Mutat. Res., 322: 61-67.
Direct Link - AOAC, 1990. Official Methods of Analysis. 15th Edn., Association of Official Analytical Chemists, Washington, DC., USA., pp: 200-210.
Direct Link - Barnett, A.J.G. and G.A. Tawab, 1957. A rapid method for the determination of lactose in milk and cheese. J. Sci. Food Agric., 8: 437-441.
CrossRefDirect Link - Barta, I., M. Adamkova, T. Petr and J. Bartova, 1990. Dose and time dependence of chromosomal aberration yields of bone marrow cells in male Chinese hamsters after a single i.p injection of aflatoxin B1. Mutat. Res., 244: 189-195.
Direct Link - Bartfay, W.J., M.T. Davis, J.M. Medvesan and S. Lugowsk, 2003. Milk whey protein decreases oxygen free radical production in a murine model of chronic iron-overload cardiomyopathy. J. Food Prot., 2: 85-94.
Direct Link - Baruchel, S. and G. Viau, 1996. In vitro selective modulation of cellular glutathione by a humanized native milk protein isolate in normal cells and rat mammary carcinoma model. Anticancer Res., 16: 1095-1100.
PubMedDirect Link - Bieri, J.C., G.S. Stoewsand, G.M. Briggs, R.W. Phillips, J.C. Woodward and J.J. Knapka, 1977. Report of the American Institute of Nutrition Ad. Hoc. Committee on standards for Nutritional studies. J. Nutr., 107: 1340-1348.
Direct Link - Bose, S. and S.P. Sinha, 1991. Aflatoxin-induced structural chromosomal changes and mitotic disruption in mouse bone marrow. Mutat. Res., 261: 15-19.
Direct Link - Bosselaers, I.E., P.W. Caessens, M.A. van Boekel and G.M. Alink, 1994. Differential effects of milk proteins, BSA and soy protein on 4NQO- or MNNG-induced SCEs in V79 cells. Food Chem. Toxicol., 32: 905-909.
PubMedDirect Link - Bounous, G., F. Gervais, V. Amer, G. Batist and P. Gold, 1989. The influence of dietary whey protein on tissue glutathione and disease of aging. Clin. Invest. Med., 12: 343-349.
PubMedDirect Link - Bounous, G., G. Batist and P. Gold, 1991. Whey protein in cancer prevention. Cancer Lett., 1- 57: 91-94.
PubMedDirect Link - Bounous, G., 2000. Whey protein concentrate (WPC) and glutathione modulation in cancer treatment. Anticancer Res., 20: 4785-4792.
PubMedDirect Link - Cassand, P., H. Abdelali, C. Bouley, G. Denariaz and J.F. Narbone, 1994. Inhibitory effect of dairy products on the mutagenicities of chemicals and dietary mutagens. J. Dairy Res., 61: 545-552.
PubMedDirect Link - Davis, N.D., U.L. Dioner and D.W. Dridge, 1966. Production of aflatoxins B1 and G1 by Aspergillus flavus in a semisynthetic medium. Applied Microbiol., 14: 378-380.
Direct Link - Devaux, C., R. Varin, P. Mulder, V. Richard and C. Thuillez, 2001. Oxidative stress and endothelial dysfunction in heart failure. Therapie, 56: 575-581.
Direct Link - Esteve, J.M., J. Mompo, J. Garcia de la Asuncion, J. Sastre and M. Asensi et al., 1999. Oxidative damage to mitochondrial DNA and glutathione oxidation in apoptosis: Studies in vivo and in vitro. FASEB J., 13: 1055-1064.
Direct Link - Gilliland, S.E., 1990. Health and nutritional benefits from lactic acid bacteria. FEMS Microbiol. Rev., 7: 175-188.
CrossRefPubMedDirect Link - Griffith, O.W., 1999. Biologic and pharmacologic regulation of mammalian glutathione synthesis. Free Radic. Biol. Med., 27: 922-935.
Direct Link - Hakkak, R., S. Korourian, S.R. Shelnutt, S. Lensing, M.J. Ronis and T.M. Badger, 2000. Diets containing whey proteins or soy protein isolate protect against 7,12-dimethylbenz(a)anthracene-induced mammary tumors in female rats. Cancer Epidemiol. Biomark. Prev., 9: 113-117.
PubMedDirect Link - Hosoda, M.H., H. Hashimito, M. Morita and A. Chibaand Hosono, 1992. Antimutagenicity of milk cultured with lactic acid bacteria against N-methyl-N'-nitro-N-nitrosoguanidine. J. Dairy Sci., 75: 976-981.
CrossRefDirect Link - Hosono, A.K.N., A. Shashikanth and H. Otani, 1988. Antimutagenic activity of whole casein on pepper-induced mutagenicity to streptomycin-dependent strain SD 510 of Salmonella typhimurium TA 98. J. Dairy Res., 55: 435-442.
CrossRefDirect Link - Hsieh, D.P. and D.N. Atkinson, 1991. Bisfuranoid mycotoxins: Their genotoxicity and carcinogenicity. Adv. Exp. Med. Biol., 283: 525-532.
PubMedDirect Link - Jongen, W.F., M.S. van Boekel and L.W. van Broekhoven, 1987. Inhibitory effect of cheese and some food constituents on mutagenicity generated in Vicia faba after treatment with nitrite. Food Chem. Toxicol., 25: 141-145.
PubMedDirect Link - Kennedy, R.S., G.P. Konok, G. Bounous, S. Baruchel and T.D. Lee, 1995. The use of a whey protein concentrate in the treatment of patients with metastatic carcinoma: A phase I-II clinical study. Anticancer Res., 15: 2643-2659.
Direct Link - Kennedy, R.S., G.P. Konok, G. Bounous, S. Baruchel and T.D. Lee, 1995. The use of a whey protein concentrate in the treatment of patients with metastatic carcinoma: A phase I-II clinical study. Anticancer Res., 15: 2643-2659.
Direct Link - Kent, K.D., W.J. Harper and J.A. Bomser, 2003. Effect of whey protein isolate on intracellular glutathione and oxidant-induced cell death in human prostate epithelial cells. Toxicol. In Vitro, 17: 27-33.
PubMedDirect Link - Krishnamurthy, P.B. and G.S. Neelaram, 1986. Effect of dietary fat on aflatoxin B1-induced chromosomal aberrations in mice. Toxicol. Lett., 31: 229-234.
CrossRef - Lu, S.C., 1999. Regulation of hepatic glutathione synthesis: Current concepts and controversies. FASEB J., 13: 1169-1183.
CrossRefDirect Link - Matar, C., S.S. Nadathur, A.T. Bakalinsky and J. Goulet, 1997. Antimutagenic effects of Milk fermented by Lactobacillus helveticus L89 and a protease-deficient derivative. J. Dairy Sci., 80: 1965-1970.
Direct Link - McIntosh, G.H., G.O. Regester, R.K. Le Leu, P.J. Royle and G.W. Smithers, 1995. Dairy proteins protect against dimethylhydrazine-induced intestinal cancers in rats J. Nutr., 125: 809-816.
PubMedDirect Link - Micke, P., K.M. Beeh, J.F. Schlaak and R. Buhl, 2001. Oral supplementation with whey proteins increases plasma glutathione levels of HIV-infected patients. Eur. J. Clin. Investigat., 31: 171-178.
Direct Link - Micke, P., K.M. Beeh and R. Buhl, 2002. Effects of long-term supplementation with whey proteins on plasma glutathione levels of HIV-infected patients. Eur. J. Nutr., 41: 12-18.
Direct Link - Morr, C.V. and E.Y.W. Ha, 1993. Whey protein concentrates and isolate processing and functional properties. Crit. Rev. Food Sci. Nutr., 33: 431-476.
Direct Link - Nadathur, S.R., S.J. Gould and A.T. Bakalinsky, 1994. Antimutagenicity of fermented milk. J. Dairy Sci., 77: 3287-3295.
PubMedDirect Link - Nadathur, S.R., S.J. Gould and A.T. Bakalinsky,Nadathur, S.R., S.J. Gould and A.T. Bakalinsky, 1995. Antimutagenicity of an acetone extract of yogurt. Mutat. Res., 334: 213-224.
CrossRef - Nelson, W.G., A.M. De Marzo and T.L. DeWeese, 2001. The molecular pathologenesis of prostate cancer: Implications for prostate cancer prevention. Urology, 57: 39-45.
PubMedDirect Link - Parodi, P.W., 1998. A role for milk proteins in cancer prevention. Aust. J. Dairy Technol., 38: 144-148.
Direct Link - Preston, R.J., B.J. Dean, S. Galloway, H. Holden, A.F. Mcfee and M. Shelby, 1987. Mammalian in vivo cytogenetic assays analysis of chromosome aberrations in bone marrow cells. Mutat. Res./Genet. Toxicol., 189: 157-165.
CrossRefPubMedDirect Link - Reeves, P.G., F.H. Nielsen and G.C. Fahey Jr., 1993. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr., 123: 1939-1951.
CrossRefPubMedDirect Link - Salassidis, K., U. Kulka, E. Schmid, D. Paul and M. Bauchinger, 1991. Induction of chromosome aberrations and sister chromatid exchange by indirectly acting mutagens in immortal mouse and rat hepatocyte lines. Mutagenesis, 6: 59-63.
PubMedDirect Link - Salamone, M., J. Heddle, E. Stuart and M. Kotz, 1980. Towards as improved micronucleus test: Studies on 3 model agents. Mitomycin C. cyclophosphamide and dimethylbenzanthracene. Mutat. Res., 74: 347-356.
CrossRef - Schneider, W.C., 1957. Determination of Nucleic Acids in Tissues by Pentose Analysis. In: Methods in Enzymology, Colowick, S.P. and N.O. Kaplan (Eds.). Vol. 3, Academic Press, New York, USA., pp: 680-684.
Direct Link - Sekine, K., E. W atanabe, J. Nakamura, N. Takasuka and D.J. Kim et al., 1997. Inhibition of azoxymethane-initiated colon tumor by bovine lactoferrin administration in F344 rats. Japanese J. Cancer Res., 88: 523-526.
Direct Link - Sen, C.K., 1997. Nutritional biochemistry of cellular glutathione. Nut. Biochem., 8: 660-672.
CrossRefDirect Link - Sengstag, C., B. Weibel and M. Fasullo, 1996. Genotoxicity of aflatoxin B1: Evidence for a recombination-mediated mechanism in Saccharomyces cerevisiae. Cancer Res., 56: 5457-5465.
Direct Link - Shannon, G.M., O.L. Shotwell and W.F. Kwolek, 1983. Extraction and thin layer chromatography of aflatoxin B1 in mixed feeds. J. Assoc. Anal. Chem., 66: 582-586.
PubMedDirect Link - Smela, M.E., S.S. Currier, E.A. Bailey and J.M. Essigmann, 2001. The chemistry and biology of aflatoxin B1: From mutational spectrometry to carcinogenesis. Carcinogenesis, 22: 535-545.
CrossRefPubMedDirect Link - Smithers, G.W., F.J. Ballard, A.D. Copeland, K.J. De Silva and D.A. Dionysius et al., 1996. New opportunities from the isolation and utilization of whey proteins. J. Dairy Sci., 79: 1454-1459.
Direct Link - Taniguchi, M., A. Yasutake and K. Takedomi, 2000. Effects of dietary sulfur-containing amino acids on oxidative damage in rat liver caused by N-nitrosodimethylamine administration. Br. J. Nutr., 84: 211-217.
PubMedDirect Link - Teramoto, S., T. Tomita, H. Matsui, E. Ohga, T. Matsuse and Y. Ouchi, 1999. Hydrogen peroxide-induced apoptosis and necrosis in human lung fibroblasts: Protective roles of glutathione. Jap. J. Pharmacol., 79: 33-40.
PubMedDirect Link - Tsai, W.Y., W.H. Chang and C.H. Chen, 2000. Enhancing effect of patented whey protein isolate (immunocal) on cytotoxcity of an anticancer drug. Nutr. Cancer, 38: 200-208.
Direct Link - Tsuda, H., K. Sekine, Y. Ushida, T. Kuhara, N. Takasuka, M. Iigo, B.S. Han and M.A. Moore, 2000. Milk and dairy products in cancer prevention: Focus on bovine lactoferrin. Mutat. Res., 462: 227-233.
CrossRefDirect Link - Umegaki, K., H. Uramoto and T. Esashi, 1997. Lack of influence of a long-term high or low vitamin E diet on the oxidative DNA damage in the bone marrow of mice. Int. J. Vitamin Nutr. Res., 67: 149-154.
PubMedDirect Link - Van Boekel, M.S., C.M. Weerens, A. Holstra, C.E. Scheidtweiler and G.M. Alink, 1993. Antimutagenic effects of casein and its digestion products. Food Chem. Toxicol., 31: 731-737.
CrossRef - Van Rensburg, S.J., P. Cook-Mozaffari, D.J. Van Schalwyck, J.J. Van der Watt, T.J. Vincent and I.F. Purchase, 1985. Hepatocellular carcinoma and dietary aflatoxin in mozambique and transkei. Br. J. Cancer, 51: 713-726.
PubMedDirect Link - Yeh, F.S., M.C. Yu, C.C. Mo, S. Luo, M.J. Tong and B.J. Herderson, 1989. Hepatitis B virus, aflatoxins and hepatocellular carcinoma in southern Guangxi, China. Cancer Res., 49: 2506-2509.
PubMedDirect Link