
Wulansih D. Astuti
Research Center for Biotechnology, Indonesian Institute of Sciences, 16911 Cibinong, West Java, Indonesia
Komang G. Wiryawan
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Bogor Agricultural University, Jalan Agatis, Campus IPB Darmaga, 16680 Bogor, West Java, Indonesia
LiveDNA: 62.23261
Elizabeth Wina
Indonesian Research Institute for Animal Production, Bogor, Jl. Veteran III, Banjarwaru, P.O. Box 221, 16002 Bogor, West Java, Indonesia
Yantyati Widyastuti
Research Center for Biotechnology, Indonesian Institute of Sciences, 16911 Cibinong, West Java, Indonesia
Sri Suharti
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Bogor Agricultural University, Jalan Agatis, Campus IPB Darmaga, 16680 Bogor, West Java, Indonesia
Roni Ridwan
Research Center for Biotechnology, Indonesian Institute of Sciences, 16911 Cibinong, West Java, Indonesia
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 3 | Page No.: 131-139







ABSTRACT
Background and Objective: Probiotics are widely used in ruminant production, but information about the potential of Lactobacillus plantarum (L. plantarum) as a probiotic for ruminants is still limited. The aim of this research was to select L. plantarum strains as a probiotic for ruminants and to determine their effect on the rumen fermentation system. Materials and Methods: The first experiment was conducted using a randomized block design to select 14 strains of L. plantarum isolated from rumen cattle. The second experiment was arranged in a completely randomized design using two selected L. plantarum strains to determine their effects as a probiotic on rumen fermentation. The substrates used for in vitro fermentation were napier grass (Pennisetum purpureum) and concentrate in a 70:30 ratio. Results: From experiment 1, L. plantarum U32 was selected, because it produced low methane/total gas (27.39%) and strain U40 was selected because it had the highest dry matter and organic matter rumen disappearance (56.45 and 56.44%). In experiment 2, the addition of L. plantarum U32 and U40 as probiotics increased propionic acid and decreased acetic production (p<0.05), which led to a lower A:P ratio (p<0.05). The total volatile fatty acid and in vitro digestibility were not affected by the addition of L. plantarum. Probiotic addition increased lactic acid bacteria and the protozoa population (p<0.05) from the rumen fluid compared to the control. The total rumen bacteria were not significantly changed by the treatments. Conclusion: The addition of L. plantarum strains U32 and U40 as probiotics had beneficial effects for rumen fermentation due to increased propionic acid and decreased methane production.
PDF Abstract XML References Citation
Received: October 17, 2017;
Accepted: February 08, 2018;
Published: February 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Wulansih D. Astuti, Komang G. Wiryawan, Elizabeth Wina, Yantyati Widyastuti, Sri Suharti and Roni Ridwan, 2018. Effects of Selected Lactobacillus plantarum as Probiotic on In vitro Ruminal Fermentation and Microbial Population. Pakistan Journal of Nutrition, 17: 131-139.
DOI: 10.3923/pjn.2018.131.139
URL: https://scialert.net/abstract/?doi=pjn.2018.131.139
DOI: 10.3923/pjn.2018.131.139
URL: https://scialert.net/abstract/?doi=pjn.2018.131.139
INTRODUCTION
Ruminant productivity can be improved by increasing feed utilization through the manipulation of the microbial ecosystem of the rumen. The use of living microbial supplements as probiotics provides a suitable alternative to antibiotics, because it does not leave residues or cause toxicity in livestock products. Probiotics are living microbial feed supplements that may beneficially affect the host animal upon ingestion by improving its intestinal microbial balance1. Probiotics are also defined as living microbes that when administered in adequate amounts, confer a health benefit to the host2.
Major probiotic strains are Lactobacillus, Saccharomyces, Bacillus, Streptococcus and Aspergillus3-5. Bacterial probiotic strains may be classified as lactic acid producing bacteria, lactic acid utilizing bacteria, or other microorganisms6,7. Lactic acid bacteria (LAB) already used as probiotics for ruminants are species of Lactobacillus, Bifidobacterium, Enterococcus, Streptococcus, Bacillus and Propionibacterium8. LAB is naturally found in many different habitats, such as fermented foods, fruits, grains, animal digestive tract or silage and it ferments sugars or carbohydrates to produce lactic acid. In the digestive tract of cattle, LABs were found in the rumen9,10. They could interact with rumen microorganisms and enhanced rumen microbial activity, improved rumen degradability11 and has shown potential to reduce methane emissions12.
One LAB that has the potential to serve as a probiotic is Lactobacillus plantarum, which produces lactic acid from their metabolism. The addition of L. plantarum cultures to in vitro rumen fermentation can increase propionic production and decrease acetic acid production and cumulative methane production13. In contrast, another in vitro study has reported no reductions in methane production14. There is also evidence that L. plantarum significantly increased digested organic matter and increased total volatile fatty acid (VFA) production. The effect of L. plantarum on in vitro rumen fermentation was influenced by dosage and the bacterial strain used in the experiment15.
The cause of improved animal performance by the addition of LAB is not completely clear and the response in ruminants is inconsistent. Strain differences affect the probiotic ability to improve rumen fermentation. Information on the effect of L. plantarum addition on rumen fermentation is lacking in the literature. Therefore, it is necessary to select L. plantarum strains that have a beneficial effect on rumen fermentation. The objective of this study was to select L. plantarum strains as probiotics for ruminants and to evaluate the effect of the L. plantarum addition on in vitro rumen fermentation and the microbial population.
MATERIALS AND METHODS
Lactobacillus plantarum strains: Fourteen strains of L. plantarum were obtained from the collection of the Laboratory of Applied Microbiology, Research Center for Biotechnology, Indonesian Institutes of Sciences, Indonesia. All strains were grown in MRS (de Man Rogosa Sharpe) broth medium and incubated at 39°C for 20 h, under anaerobic conditions, using Hungate tubes flushed with CO2.
In vitro rumen fermentation: Rumen fluids were obtained from two rumen-fistulated Ongole crossbred cattle before the morning feeding (mixed in 1:1 ratio). The rumen-fistulated Ongole crossbred cattle were managed according to the protocols approved by the Ethic Clearance Committee of Indonesian Institute of Sciences (Number 9879/WK/HK/XI/2015). Rumen fluid was filtered through a double layer of cheese cloth for in vitro studies, pooled in pre-warmed bottles, sealed and immediately transported to the laboratory. The substrate used for in vitro rumen fermentation was a mixture of concentrate feed and dried milled elephant grasses (Pennisetum purpureum), with 70:30 ratio. The concentrate consisted of rice bran, corn, corn gluten feed, coconut meal, palm kernel meal, pollard, soybean meal, mineral mix, DCP (dicalcium phosphate) and NaCl and it contained 18% crude protein, 9.8% crude fiber and 70% TDN (Total Digestible Nutrients). Approximately 0.75 g of substrates (consisting of 70% P. purpureum and 30% concentrate proportion) was put inside the serum bottle glass and filled with 75 mL mixture rumen fluid and Mc’Dougall buffer. The bottle was closed with a rubber cap and an aluminum crimp after it was flushed with CO2 gas for 30 sec to obtain anaerobic conditions. Then, the bottle was incubated in a water bath incubator at a temperature of 39°C16.
Experiment 1: In vitro rumen fermentation was conducted to find the best candidates from 14 strains of L. plantarum as ruminant probiotics. The experimental design was arranged in a randomized block design with 3 replications and 15 treatments. The control treatment was rumen fermentation without the addition of L. plantarum and the other 14 treatments were rumen fermentation with L. plantarum addition. One mL of each L. plantarum strain (109 CFU mL–1) was added to each experimental tube. Total gas and methane production were measured at 3, 6, 9, 12 and 24 h of fermentation. Dry matter and organic matter rumen disappearance (DMRD and OMRD) were measured after 24 h of fermentation.
Experiment 2: Two strains of L. plantarum from experiment 1 were selected as probiotic candidates and used for further analysis in experiment 2. The experiment was arranged in a completely randomized design with 5 replications and 3 treatments, consisting of a control (without addition L. plantarum), the addition of L. plantarum strain U32 and the addition of L. plantarum strain U40. One mL of each L. plantarum strain (109 CFU mL–1) was added to each experimental tube. A rumen fluid sample was collected for NH3, VFA, protozoal number, rumen bacterial and LAB population after 4 h of incubation. In vitro dry matter digestibility (DMD) and organic matter digestibility (OMD) was measured after 2×48 h of fermentation, with the addition of pepsin-HCl after 48 h of fermentation.
Parameters measured: The total gas and methane production in experiment 1 was measured using a glass syringe according to the methods of Fievez et al.17. Rumen pH was measured with a pH meter. The concentration of NH3 was measured by the microdiffusion Conway method18. The total VFA concentration and molar proportions of VFA were analyzed using gas chromatography (GC 8A, Shimadzu Corp., Kyoto, Japan, with capillary column type containing 10% of SP-1200, 1% of H3PO4 on 80/100 Cromosorb WAW and nitrogen as the gas carrier). The total rumen bacteria were quantified using 9-85 medium with rolled-tube19. The LAB population was quantified with total plate count (TPC) methods using an MRS agar plate in the form of Colony-Forming Units (CFU) incubated in 39°C for 24 h in an anaerobic condition using an anaerobic jar with anaeropack to reduce the oxygen. The number of protozoa in the buffered rumen fluid was counted after 4 h of incubation under a microscope. The contents of the fermentation tubes were mixed and a 1 mL aliquot was taken and mixed with 1 mL of methyl green formaldehyde saline solution, containing 35% formaldehyde, distilled water, methyl green and NaCl. The stained sample was kept at room temperature. Protozoa populations were counted with Fuchs Rosenthal Counting Chamber (4×4×0.2 mm) under a microscope (10×10)19. Dry matter and organic matter disappearance (DMRD and OMRD) were measured after 24 h of fermentation, while dry matter and organic matter digestibility (DMD and OMD) were measured after 2×48 h of fermentation, with the addition of pepsin-HCl after 48 h of fermentation, using the Tilley and Terry20 method.
Statistical analysis: Data were analyzed by a one-way analysis of variance using SPSS 16 (SPSS, Inc., IBM, Chicago). Significant effects of treatments were determined by Duncan's multiple range test. Significant differences were accepted if p<0.05.
RESULTS
Experiment 1: The total gas production from 24 h in vitro rumen fermentation of each L. plantarum strain (14 strains) was significantly higher than the control treatment (p<0.05) (Table 1). The highest total gas production was observed in the treatment with the addition of L. plantarum strain U32 (146.67 mL), while the control treatment produced the lowest gas production (123.67 mL). A similar result also obtained from methane production and the control treatment produced the lowest methane (34.67 mL). The addition of L. plantarum significantly increased (p<0.05) methane production, but there were no significant differences among L. plantarum strains used in this experiment. The lowest % methane/total gas production resulted from addition of L. plantarum strain U90 (26.77%), which was significantly lower compared to the highest % methane/total gas produced by strain U40 (28.91%). None of the treatments were affected by the final pH of the rumen fluid after 24 h in vitro incubation, which ranged from 6.93-7.00.
| Table 1: | Total gas, methane production, methane production/total gas and pH of in vitro rumen fermentation with the addition of different strains of L. plantarum |
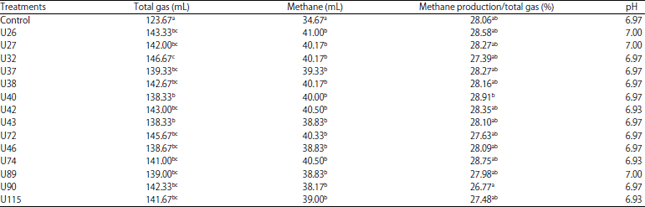 | |
a-c Means with different superscripts within columns significantly differed (p<0.05) | |
| Table 2: | Dry matter and organic matter rumen disappearance, total gas/digested organic matter and methane/digested organic matter of in vitro rumen fermentation with the supplementation of L. plantarum |
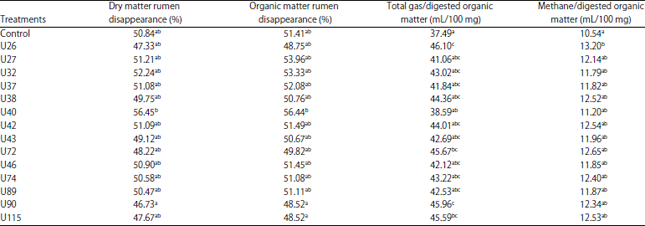 | |
a-cMeans with different superscripts within columns significantly differed (p<0.05) | |
| Table 3: | Volatile fatty acid (VFA) production of in vitro rumen fermentation with the addition of L. plantarum |
 | |
a-cMeans with different superscripts within columns significantly differed (p<0.05), A/P: Acetic acid/propionic acid | |
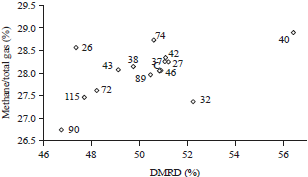 | |
| Fig. 1: | Correlation of DMRD (%) and methane/total gas production (%) from in vitro rumen fermentation with the addition of L. plantarum |
The addition of L. plantarum as a probiotic did not significantly affect in vitro dry matter and organic matter rumen disappearance (DMRD and OMRD) (Table 2) compared to the control. L. plantarum strain U40 resulted in the highest DMRD and OMRD (56.45 and 56.44%, respectively), which were significantly higher than the lowest DMRD and OMRD (46.73 and 53%, respectively) that resulted from strain U90. The production of methane from each 100 g DMRD of feed was calculated to determine which L. plantarum strain produced the lowest methane. Among all strains used in this experiment, L. plantarum strain U40 produced the lowest methane/100 mg digested organic matter, although it was not significantly lower than the highest methane production from L. plantarum strain U26. However, from all treatments, the control without the addition of L. plantarum produced lower methane/100 mg digested organic matter, although it was not significantly different from the other treatments, except from L. plantarum strain U26.
The correlations between in vitro DMRD and % methane/total gas production of each L. plantarum strain used in this experiment are shown in Fig. 1. L. plantarum strain U40 showed the highest DMRD among treatments, but high % methane/total gas production, while strain U32 showed the lower % methane/total gas production but also lower DMRD. L. plantarum strain U26 showed the lowest % methane/total gas production but also had the lowest DMRD among treatment.
Experiment 2: The results from experiment 2 found that L. plantarum strains U32 and U40 were candidates for probiotics. The total and individual VFA productions from in vitro rumen fermentation are shown in Table 3. The total VFA production was not significantly affected by the addition of both L. plantarum strains U32 and U40. The addition of L. plantarum as a probiotic changed the composition of VFA. Acetic acid significantly decreased (p<0.05), while propionic acid significantly increased (p<0.05) compared to the control treatment.
| Table 4: | Rumen fermentation characteristic from in vitro rumen fermentation with the addition of L. plantarum |
DMD: Dry matter digestibility, OMD: Organic matter digestibility, a-cMeans with different superscripts within columns significantly differed (p<0.05) | |
| Table 5: | Rumen microbes population of in vitro rumen fermentation with the supplementation of L. plantarum |
 | |
a-cMeans with different superscripts within columns significantly differed (p<0.05) | |
L. plantarum strain U40 produced the lowest proportion of acetic acid (51.71%), followed by L. plantarum strain U32 (56.74%), which was significantly higher than U40. The control treatment without the addition of L. plantarum produced the highest proportion of acetic acid (63.60%). The highest propionic acid was produced by the addition of L. plantarum strain U40 (25.42%), which was not significantly different from L. plantarum strain U32 (25.28%). The lowest propionic acid was produced by the control, which was significantly lower than the other treatments (21.18%).
Butyric and valeric acid significantly increased (p<0.05) with the addition of L. plantarum strain U40 compared to the control treatment. Although the addition of L. plantarum strain U32 also increased butyric and valeric acid, it was not significantly compared to the control treatment. Higher propionic acid and lower acetic acid production by L. plantarum addition resulted in a significantly (p<0.05) lower acetic:propionic acid (A:P) ratio compared to the control. The predicted methane production was calculated from acetic, propionic and butyric acid production using the Moss equation21. The addition of L. plantarum strains U32 and U40 decreased methane production from in vitro rumen fermentation between 18.43-21.62%.
The in vitro dry matter digestibility (DMD) and organic matter digestibility (OMD) were not significantly affected by the addition of L. plantarum as a probiotic and the values were comparable to the control (Table 4). The production of NH3 significantly increased (p<0.05) with the addition of L. plantarum compared to the control, but both strains showed comparable production of NH3. The NH3 production of the control was 8.48% and it increased to 17.17 and 16.36% with the addition of strains U32 and U40, respectively. Treatments were not significantly effect pH of rumen fluid, which varied between 6.95 (L. plantarum U32 and U40) to 6.98 (control).
The populations of total rumen bacteria, LAB and protozoa are shown in Table 5. Rumen bacteria were not significantly affected by the addition of L. plantarum U32 (10.32 CFU mL–1) and U40 (9.90 CFU mL–1), although it was lower compared to the control (10.57 CFU mL–1). The addition of L. plantarum significantly increased (p<0.05) the LAB population in the rumen. The highest LAB population (log 10) resulted from the addition of L. plantarum strain U32 (8.92 CFU mL–1), which was significantly higher than strain U40 (8.10 CFU mL–1). The control treatment without the addition of L. plantarum resulted in the lowest LAB population (6.64 CFU mL–1), which was significantly lower than the other treatments. Protozoal number (log 10) also significantly increased (p<0.05) with the addition of LAB as probiotics, from 3.72 CFU mL–1 (control) to 4.099 and 4.121 CFU mL–1 with the addition of L. plantarum strains U32 and U40, respectively.
DISCUSSION
The selection of L. plantarum strains as candidates for ruminant probiotics was based on their gas production and rumen disappearance during in vitro rumen fermentation. The total gas production results showed that L. plantarum strain U32 produced the highest gas. The addition of a probiotic must have a beneficial effect for rumen fermentation, such as increased feed digestibility or lowered methane production. A different result was obtained when % methane/total gas production was calculated, which showed strain U90 as the lowest methane producer. Experiment 1 did not measure VFA production, therefore, methane gas production becomes an important parameter for selecting the best candidates for ruminant probiotics.
The increased total gas production was usually associated with the increase of dry matter and organic matter rumen disappearance (DMRD and OMRD). The effects of L. plantarum addition on gas and methane production may appear during the initial stages of in vitro fermentation but are largely absent at the end of the incubation15. From the DMRD parameter, methane production/100 mg digested organic matter can be calculated. The results showed that the lowest amount of methane was produced by L. plantarum strain U40 in this experiment. Lower methane production indicates more efficient rumen fermentation, but low % methane production from strain U90 resulted from poor feed digestibility in the rumen. The digestibility from strain U90 was the lowest among all treatments. Therefore, although L. plantarum strain U90 produced the lowest % methane production, it would not be selected as a probiotic candidate.
The correlation of DMRD and % methane/total gas production is shown in Fig. 1. The selection of the best candidates of L. plantarum strains for probiotics for ruminants was based on the highest DMRD and % methane/total gas production. It can be seen that L. plantarum strain U40 showed the highest DMRD, while strain U32 showed the lowest % methane/total gas production. As previously mentioned, although strain U90 resulted in the lowest methane production, it resulted from poor digestibility. U90 was not selected, because it showed the lowest DMRD. Therefore, L. plantarum strains U40 and U32 can be selected as candidates for ruminant probiotics.
A further experiment on Lactobacillus plantarum strains U40 and U32 was conducted to reveal more details about their effects on in vitro rumen fermentation. In this experiment, the addition of L. plantarum strains U40 and U32 significantly changed the composition of VFA produced from rumen fermentation to higher propionic acid and lower acetic acid production. Total VFA production was affected by the addition of L. plantarum strains U40 and U32, although they were slightly reduced. Increased propionic acid, with the addition of LAB, was reported within the study conducted by Soriano et al.22, but in that experiment, there was no decrease in acetic acid production. The addition of LAB may stimulate the growth of lactic acid utilizing bacteria23, leading to increased production of propionic acid as a result of their metabolism. When higher propionic acid was produced, the supply of H2 for methane production in the rumen was reduced.
The changes in rumen fermentation products to higher propionic acid and lower acetic acid, followed with a lower A:P ratio, indicated the potential of L. plantarum added as a probiotic for ruminants toward more efficient rumen fermentation by lower methane production. More efficient rumen fermentation by increased propionic acid could improve growth efficiency24. Methane produced by ruminants represents an energy loss for the host animal of 2-12% of dietary energy21. Therefore, decreased methane production will increase the energy supply of the animal, followed by increased productivity. O’Brien et al.13 reported high methane reduction with the addition of L. plantarum, with more than a 60% decrease in cumulative methane production from in vitro rumen fermentation, but total VFA production (mM) was significantly reduced. The reduction of methane production was due to the general suppression of the fermentation process, when L. plantarum was directly added into the in vitro bottle. In this experiment, the reduction of methane was approximately 18-20%, without a significant reduction of total VFA production. This result indicated that there was no negative impact on rumen fermentation from the addition of L. plantarum strains U32 and U40.
VFA production in the rumen highly depends on the degree and rate of fermentation25,26. In vitro dry matter digestibility (DMD) and organic matter digestibility (OMD) were not significantly affected by the addition of L. plantarum as a probiotic and the values were comparable with the control (Table 4). In this experiment, the digestibility results were in line with total VFA production. There were no significant changes in either parameters compared with control, which indicated that rumen microbes were still working normally with the addition of L. plantarum as a probiotic measured from rumen fermentation products. An experiment by Contreras-Govea et al.14 reported significantly increased digestibility with the addition of L. plantarum. Increased digestibility showed that the addition of LAB can stimulate fibrolytic bacteria in the rumen. These results showed that the effect of the addition of L. plantarum as a probiotic on rumen fermentation depends on the type of strains, dose and substrate incubated 27.
The addition of L. plantarum strains U40 and U32 did not significantly change the total bacteria population, although a numerical decrease was observed. This result was in agreement with another experiment28 that reported decreased total rumen bacteria at 3 h after the addition of a probiotic consisting of L. plantarum+Propionibacterium. Although other bacteria were added from outside into the rumen ecosystem as a probiotic, it is rare that it can cause significant changes to the total bacteria population. The lack of differences in the total bacteria population caused by treatments could be a reason why the total VFA and digestibility were also not affected by treatments.
Lactic acid bacteria can only have beneficial effects as a probiotic in the rumen if they could survive in the rumen. Although this experiment did not measure the population of L. plantarum specifically, a higher population of LAB by the addition of L. plantarum indicated that LAB added from outside can survive during rumen fermentation and stimulate other LABs to survive in the rumen. Although LAB species are facultative anaerobic bacteria, they survive in both the rumen and intestine29. Lactic acid bacteria could survive in the rumen during in vitro fermentation, particularly when sugar substrates were used for fortification30. The survival of L. plantarum used in this experiment might be because they were isolated from rumen cattle. It might help them to adapt more easily in an anaerobic rumen environment.
The increased protozoal number with the addition of L. plantarum as a probiotic can be correlated with the increased in NH3 production. An increased protozoal number was also reported after the probiotic addition consisted of LAB and Propionibacterium after acidosis induction28. Ammonia was produced by protozoa as a result of the digestion of bacteria by protozoa. The increased protozoal number, along with decreased methane production, was unpredictable, because 9-25% of methanogenesis in the rumen was produced by methanogens associated with protozoa31. In addition, the, defaunation of protozoa has been known as a method to decrease methane production in the rumen. In this experiment, decreased methane production may not be caused by the decrease in methanogens, but it more likely is caused by the change in rumen microbial diversity, because L. plantarum addition stimulated propionate producer and fermentation shifts that divert H2 away from methanogenesis32,33. Supplementation of live LAB was assumed to modify the rumen microbial population and help rumen microbes adapt to lactic acid in the rumen, in order to reduce the incidence of acute ruminal acidosis34-37. Lactic acid utilizer was probably stimulated by lactic acid produced by the L. plantarum addition and produced propionate production as metabolites products.
The implication of this experiment is that the addition of L. plantarum as a probiotic beneficially affected rumen fermentation, which changed to propionate production. Rumen fermentation is more efficient by producing less methane. These results showed that the application of L. plantarum as a probiotic is feasible and negative effects caused by L. plantarum addition were not detected. More experiments regarding an in vivo trial of L. plantarum addition is recommended to fulfill the effects of L. plantarum as a probiotic for ruminants.
CONCLUSION
Based on results from the experiments, L. plantarum strains U32 and U40 were selected as probiotics for ruminants based on their potential to affect rumen fermentation. The results also indicated that the addition of L. plantarum has beneficial effects in the rumen. The addition of L. plantarum strains U32 and U40 on in vitro rumen fermentation as probiotics changed toward more efficient rumen fermentation by increased propionic acid and lowered acetic acid proportions, indicating lower methane production, which provides higher energy for the animal.
SIGNIFICANCE STATEMENT
This study discovers probiotics for ruminants that contain L. plantarum as a lactic acid producer. This study will help the researchers to uncover the effect of L. plantarum addition as a probiotic for rumen fermentation and rumen microbes that many researchers were not able to explore. Thus, a new theory on the role of L. plantarum as a probiotic for ruminants may be arrived at.
ACKNOWLEDGMENT
The authors wish to acknowledge the financial support received from "DIPA Unggulan-LIPI (2015-2017) Food Security and Drug’s Discovery" of the Research Center for Biotechnology-Indonesian Institute of Sciences (LIPI) and LPDP (Lembaga Pengelola Dana Pendidikan) scholarship.
REFERENCES
- Fuller, R., 1989. Probiotics in man and animals. J. Applied Bacteriol., 66: 365-378.
CrossRefPubMedDirect Link - Jiao, P.X., L.Y. Wei, N.D. Walker, F.Z. Liu, L.Y. Chen, K.A. Beauchemin and W.Z. Yang, 2017. Comparison of non-encapsulated and encapsulated active dried yeast on ruminal pH and fermentation, and site and extent of feed digestion in beef heifers fed high-grain diets. Anim. Feed Sci. Technol., 228: 13-22.
CrossRefDirect Link - Antonius, K.G. Wiryawan, A. Thalib and A. Jayanegara, 2015. Digestibility and methane emission of ration based on oil palm by products supplemented with probiotics and banana stem: An in vitro study. Pak. J. Nutr., 14: 37-43.
CrossRefDirect Link - Tannock, G.W., 2001. Molecular assessment of intestinal microflora. Am. J. Clin. Nutr., 73: 410-414.
CrossRefDirect Link - Elghandour, M.M.Y., A.Z.M. Salem, J.S.M. Castaneda, L.M. Camacho, A.E. Kholif and J.C.V. Chagoya, 2015. Direct-fed microbes: A tool for improving the utilization of low quality roughages in ruminants. J. Integr. Agric., 14: 526-533.
CrossRefDirect Link - Wiryawan, K.G. and J.D. Brooker, 1995. Probiotic control of lactate accumulation in acutely grain-fed sheep. Aust. J. Agric. Res., 46: 1555-1568.
CrossRefDirect Link - Seo, J.K., S.W. Kim, M.H. Kim, S.D. Upadhaya, D.K. Kam and J.K. Ha, 2010. Direct-fed microbials for ruminant animals. Asian-Aust. J. Anim. Sci., 23: 1657-1667.
CrossRefDirect Link - Bureenok, S., W. Suksombat and Y. Kawamoto, 2011. Effects of the fermented juice of epiphytic lactic acid bacteria (FJLB) and molasses on digestibility and rumen fermentation characteristics of ruzigrass (Brachiaria ruziziensis) silages. Livest. Sci., 138: 266-271.
CrossRefDirect Link - Cobos, M.A., A.L. de Coss, N.D. Ramirez, S.S Gonzales and R.F. Cerrato, 2011. Pediococcus acidilactici isolated from the rumen of lambs with rumen acidosis, 16S rRNA identification and sensibility to monensin and lasalocid. Res. Vet. Sci., 90: 26-30.
CrossRefPubMedDirect Link - Weinberg, Z.G., 2003. Effect of lactic acid bacteria on animal performance. Indian J. Biotechnol., 2: 378-381.
Direct Link - Lee, S.S., J.T. Hsu, H.C. Mantovani and J.B. Russell, 2002. The effect of bovicin HC5, a bacteriocin from Streptococcus bovis HC5, on ruminal methane production in vitro. FEMS Microbiol. Lett., 217: 51-55.
CrossRefDirect Link - O'Brien, M., T. Hashimoto, A. Senda, T. Nishida and J. Takahashi, 2013. The impact of Lactobacillus plantarum TUA1490L supernatant on in vitro rumen methanogenesis and fermentation. Anaerobe, 22: 137-140.
CrossRefDirect Link - Contreras-Govea, F.E., R.E. Muck, D.R. Mertens and P.J. Weimer, 2011. Microbial inoculant effects on silage and in vitro ruminal fermentation and microbial biomass estimation for alfalfa, bmr corn and corn silages. Anim. Feed Sci. Technol., 163: 2-10.
Direct Link - Ellis, J.L., A. Bannink, I.K. Hindrichsen, R.D. Kinley, W.F. Pellikaan, N. Milora and J. Dijkstra, 2016. The effect of lactic acid bacteria included as a probiotic or silage inoculant on in vitro rumen digestibility, total gas and methane production. Anim. Feed Sci. Technol., 211: 61-74.
CrossRefDirect Link - Theodorou, M.K., B.A. Williams, M.S. Dhanoa, A.B. McAllan and J. France, 1994. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed Sci. Technol., 48: 185-197.
CrossRefDirect Link - Fievez, V., O.J. Babayemi and D. Demeyer, 2005. Estimation of direct and indirect gas production in syringes: A tool to estimate short chain fatty acid production that requires minimal laboratory facilities. Anim. Feed Sci. Technol., 123-124: 197-210.
CrossRefDirect Link - Ogimoto, K. and S. Imai, 1981. Atlas of Rumen Microbiology. Japan Scientific Societies Press, Tokyo, ISBN: 9784762202643, Pages: 231.
Direct Link - Tilley, J.M.A. and R.A. Terry, 1963. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci., 18: 104-111.
CrossRefDirect Link - Moss, A.R., J.P. Jouany and J. Newbold, 2000. Methane production by ruminants: Its contribution to global warming. Annales Zootechnie, 49: 231-253.
CrossRefDirect Link - Soriano, A.P., L.L. Mamuad, S.H. Kim, Y.J. Choi and C.D. Jeong et al., 2014. Effect of Lactobacillus mucosae on in vitro rumen fermentation characteristics of dried brewers grain, methane production and bacterial diversity. Asian-Aust. J. Anim. Sci., 27: 1562-1570.
CrossRefDirect Link - Yoon, I.K. and M.D. Stern, 1995. Influence of direct-fed microbials on ruminal microbial fermentation and performance of ruminants-A review. Asian-Aust. J. Anim. Sci., 8: 533-555.
CrossRefDirect Link - Kenney, N.M., E.S. Vanzant, D.L. Harmon and K.R. McLeod, 2015. Direct-fed microbials containing lactate-producing bacteria influence ruminal fermentation but not lactate utilization in steers fed a high-concentrate diet. J. Anim. Sci., 93: 2336-2348.
CrossRefDirect Link - Dijkstra, J., E. Kebreab, A. Bannink, J. France and S. Lopez, 2005. Application of the gas production technique to feed evaluation systems for ruminants. Anim. Feed Sci. Technol., 123-124: 561-578.
CrossRefDirect Link - Cone, J.W. and P.M. Becker, 2012. Fermentation kinetics and production of volatile fatty acids and microbial protein by starchy feedstuffs. Anim. Feed Sci. Technol., 172: 34-41.
CrossRefDirect Link - Jiao, P.X., F.Z. Liu, K.A. Beauchemin and W.Z. Yang, 2017. Impact of strain and dose of lactic acid bacteria on in vitro ruminal fermentation with varying media pH levels and feed substrates. Anim. Feed Sci. Technol., 224: 1-13.
CrossRefDirect Link - Lettat, A., P. Noziere, M. Silberberg, D.P. Morgavi, C. Berger and C. Martin, 2012. Rumen microbial and fermentation characteristics are affected differently by bacterial probiotic supplementation during induced lactic and subacute acidosis in sheep. BMC Microbiol., Vol. 12.
CrossRefDirect Link - Rodriguez-Palacios, A., H.R. Staempfli, T. Duffield and J.S. Weese, 2009. Isolation of bovine intestinal Lactobacillus plantarum and Pediococcus acidilactici with inhibitory activity against Escherichia coli O157 and F5. J. Applied Microbiol., 106: 393-401.
CrossRefDirect Link - Weinberg, Z.G., R.E. Muck, P.J. Weimer, Y. Chen and M. Gamburg, 2004. Lactic acid bacteria used in inoculants for silage as probiotics for ruminants. Applied Biochem. Biotechnol., 118: 1-9.
CrossRefDirect Link - Newbold, C.J., B. Lassalas and J.P. Jouany, 1995. The importance of methanogens associated with ciliate protozoa in ruminal methane production in vitro. Lett. Applied Microbiol., 21: 230-234.
CrossRefDirect Link - Martin, C., L. Millet, G. Fonty and B. Michalet-Doreau, 2001. Cereal supplementation modified the fibrolytic activity but not the structure of the cellulolytic bacterial community associated with rumen solid digesta. Reprod. Nutr. Dev., 41: 413-424.
CrossRefDirect Link - Morgavi, D.P., E. Forano, C. Martin and C.J. Newbold, 2010. Microbial ecosystem and methanogenesis in ruminants. Animal, 4: 1024-1036.
CrossRefPubMedDirect Link - Nocek, J.E. and W.P. Kautz, 2006. Direct-fed microbial supplementation on ruminal digestion, health, and performance of pre-and postpartum dairy cattle. J. Dairy Sci., 89: 260-266.
CrossRefPubMedDirect Link - Krehbiel, C.R., S.R. Rust, G. Zhang and S.E. Gilliland, 2003. Bacterial direct-fed Microbials in ruminant diets: Performance response and mode of action. J. Anim. Sci., 81: E120-E132.
Direct Link - Beauchemin, K.A., W.Z. Yang, D.P. Morgavi, G.R. Ghorbani, W. Kautz and J.A. Leedle, 2003. Effects of bacterial direct-fed microbials and yeast on site and extent of digestion, blood chemistry and subclinical ruminal acidosis in feedlot cattle. J. Anim. Sci., 81: 1628-1640.
PubMedDirect Link - Ghorbani, G.R., D.P. Morgavi, K.A. Beauchemin and J.A.Z. Leedle, 2002. Effects of bacterial direct-fed microbials on ruminal fermentation, blood variables and the microbial populations of feedlot cattle. J. Anim. Sci., 80: 1977-1985.
PubMedDirect Link