
Rabia Hamid
Department of Biochemistry, University of Kashmir, Srinagar-190006, Jammu and Kashmir, India
Akbar Masood
Department of Biochemistry, University of Kashmir, Srinagar-190006, Jammu and Kashmir, India
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 3 | Page No.: 293-303







ABSTRACT
Lectins are carbohydrate binding proteins present in most plants, especially seeds and tubers like cereals, potatoes and beans. It is now well established that many lectins are toxic, inflammatory, resistant to cooking and digestive enzymes and present in much of our food and sometimes cause "food poisoning." The global pattern of varying prevalence of diseases such as coeliac disease, autoimmune diseases, rheumatoid arthritis, obesity, cardiovascular disease and insulin dependant diabetes mellitus, suggests that some dietary factor specific to plant foods could initiate these diseases. Of the food lectins, grain/cereal lectins, dairy lectins and legume lectins are the most common ones associated with aggravation of inflammatory and digestive diseases in the body and improvement of these diseases and/or symptoms when avoided. Recent research has suggested that these lectins may effectively serve as a vehicle allowing foreign proteins to invade our natural gut defenses and cause damage well beyond the gut, commonly in joints, brain, skin and various body glands. With continued exposure of the gut by these toxic food lectins a persistent stimulation of the body’s defense mechanism in a dysfunctional manner occurs, which manifests as an autoimmune disease. If the lectins in diet are causative in initiating all these diseases, it should be possible to identify the responsible constituents and modify or remove them so as to make the diet healthier. Here we present a brief account of lectin toxicity research and show how these proteins have become the focus of intense interest for biologists.
PDF Abstract XML References Citation
How to cite this article
Rabia Hamid and Akbar Masood, 2009. Dietary Lectins as Disease Causing Toxicants. Pakistan Journal of Nutrition, 8: 293-303.
DOI: 10.3923/pjn.2009.293.303
URL: https://scialert.net/abstract/?doi=pjn.2009.293.303
DOI: 10.3923/pjn.2009.293.303
URL: https://scialert.net/abstract/?doi=pjn.2009.293.303
INTRODUCTION
Lectins are carbohydrate binding proteins present in most plants, especially seeds like cereals, beans, etc., in tubers like potatoes and also in animals. Lectins selectively bind carbohydrates and importantly, the carbohydrate moieties of the glycoproteins that decorate the surface of most animal cells. Dietary lectins act as protein antigens which bind to surface glycoproteins (or glycolipids) on erythrocytes or lymphocytes (Goldstein and Etzler, 1983). They function as both allergens and hemagglutinins and are present in small amounts in 30% of foods, more so in a whole-grain diet (Nachbar and Oppenheim, 1980). Lectins have potent in vivo effects. When consumed in excess by sensitive individuals, they can cause 3 primary physiological reactions: they can cause severe intestinal damage disrupting digestion and causing nutrient deficiencies; they can provoke IgG and IgM antibodies causing food allergies and other immune responses (Gell and Coombs, 1975) and they can bind to erythrocytes, simultaneously with immune factors, causing hemagglutination and anemia. Of the 119 known dietary lectins, about half are panhemagglutinins, clumping all blood types. The remainder are blood-type specific. In general, lectins alter host resistance to infection, cause failure to thrive and can even lead to death in experimental animals (Nachbar and Oppenheim, 1980).
History: During the last 125 years, lectins have come a long way, since their first detection in plants as hemagglutinins to their present status as ubiquitous recognition molecules with myriad exciting functions and applications and have been shown to possess a remarkable array of biological activities (Sharon and Lis, 2004). In vitro they have been shown to effect lymphocyte mitogenesis both stimulating (Lis and Sharon, 1977) and inhibiting (Nachbar et al., 1980), with the lymphocytes of the gastrointestinal tract being the most susceptible (Elson et al., 1979), to possess the ability to aggregate immunoglobulins (Rouge et al., 1978), to trigger the alternate complement pathway (Forsdyke and David 1978), to inhibit fungal growth (Lis and Sharon, 1977) and to induce histamine release from basophils and mast cells (Siraganian and Siraganian, 1975). Their in vivo effects have been shown to be equally as impressive. Given orally some lectins can interact with the mucosa of the gastrointestinal tract, to cause acute gastrointestinal symptoms, failure to thrive and even death (Liener, 1974; Jaffe, 1969; Jayne-Williams and Burgess, 1974; Pusztai et al., 1979; Manage et al., 1972; Freed and Buckley, 1978) in experimental animals. Administered parenterally they can alter host resistance to infection (Lawrence and Schell, 1977; Schell and Lawrence, 1977; Dickinson et al., 1978) or to tumor challenge (Schwartz and Pappas, 1976). Under certain circumstances they may also be highly allergenic (Mitchell and Clarke, 1979; Willoughby et al., 1979). As the knowledge of their biological potency has expanded so too has speculation regarding their dietary role in a variety of pathological conditions ranging from dental caries (Rolla, 1976; Rogers, 1979; Mirth et al., 1979) to inflammatory bowel (Forsdyke, 1978) and celiac disease (Weiser and Douglas, 1976). The important point is that some of the lectins consumed in everyday foods act as chemical messengers that can in fact bind to the sugars of cells in the gut and the blood cells, initiating an inflammatory response. In wheat, gliadin, a component of gluten and an isolectin of Wheat Germ Agglutinin (WGA), is capable of activating NF kappa beta proteins which, when up-regulated, are involved in almost every acute and chronic inflammatory disorder including neurodegenerative disease, inflammatory bowel disease, infectious and autoimmune diseases (Jones and David, 2005). High levels of soybean agglutinin has been shown to produce toxic effects on mammalian organs and metabolism such as induced growth of gut, depletion of lipid in carcass, atrophy of spleen and kidney and reduction in circulating insulin (Zang et al., 2006). Scientific literature shows that dietary lectins can dramatically reduce Natural Killer (NK) cell activity directly and through disruption of intestinal flora. Natural killer cells are one of the body’s most important defenses against viruses and other invaders. Considering the vast and varied biological activities that lectins can affect it is surprising that there exists a paucity of studies related to the nutritional consequences of dietary lectins.
The reasons for the experimental gap are obscure but may, in part, be due to a lack of awareness within the medical community, of which foods in our diet contain lectin activity and the extent of exposure. A handful of investigators led by Jaffe (1969), Liener (1974); Loma and McPherson (1990) have, over the last 40 years, established a definitive relationship between the poor nutritive value of raw bean (legume) diet and its content of phytoagglutinin. Till recent times, no representatives of any plant family other than the Leguminosae have been studied for their nutritive action with the possible exception of the castor bean lectin of the family Euphorbiacae (Liener, 1974; Jaffe, 1969). Yet lectins are not exclusively found in legumes but are widely distributed throughout the plant kingdom (Gold and Balding, 1975).
Lectins in foods and human reactions: Lectins are found in a wide range of vegetables and some fruits (Nachbar and Oppenheim, 1980). In humans they have been reported to cause damage, including mass food poisoning from raw or under-cooked kidney beans (Freed, 1979; Leiner et al., 1986) and hemolytic anemia and jaundice from Mexican fava beans in Glucose-6-Phosphate dehydrogenase deficient individuals (Breneman, 1984). Plant lectins that are not efficiently degraded by digestive enzymes and that have an affinity for the surface of gut epithelial cells, such as those present in the Leguminosae family, can be poisonous (Vasconcelos and Oliveira, 2004). Acute symptoms following ingestion include nausea, vomiting and diarrhea. Long-term intake in rodent models is characterized by increased cell turnover, gut hyperplasia and weight loss. Areas of epithelial cell necrosis and even zones of complete epithelial cell denudation are seen in biopsies of the stomach and intestine of mammals (Lorenzsonn and Olsen, 1982) and insects (Sauvion et al., 2004) fed plant lectins. Indeed, the plant lectin may function as a natural insecticide. Epithelial cell microvilli particularly are affected by lectin exposure, which initiates disruption and shedding of these membrane rich surface projections (Hart et al., 1988). Confusingly, however, when cells are treated with lectins in vitro, even at very high doses, necrosis is not observed, though many other responses have been noted including mitogenesis (Kilpatrick, 1999), vacuole formation (Edelson and Cohn, 1974) and inhibition of exocytosis (Boehm and Huck, 1998).
Until recently the main use of lectins was as histology and blood transfusion reagents, but in the past few decades it has been realised that many lectins are toxic, inflammatory, or both; resistant to cooking and digestive enzymes and present in much of our food (Van Damme et al., 1998). It is thus no surprise that they sometimes cause "food poisoning." But the really disturbing finding came with the discovery in 1989that some food lectins get past the gut wall and deposit themselves in distant organs (Pusztai et al., 1989; Wang et al., 1998). So they could be one of the important environmental factors that cause real life diseases.
The edible portions of fresh and processed foods show lectin activity including such common foods as salad ingredients, fruits, spices, dry cereals and roasted nuts (Table 1). Many of the other food plants have been shown to contain phytohemagglutinins (Nachbar et al., 1980). While, in most cases, the significance of the latter is somewhat obscured since the non edible parts of the plant have been tested, nevertheless, it is quite apparent that exposure to dietary lectins is a frequent and widespread event. Although both cooking and the normal digestive processes might be expected to blunt or abrogate dietary lectin activity, this need not necessarily be the case. Liener (Liener, 1974) has pointed out that dry heat may not completely destroy lectin activity. This phenomenon is clearly illustrated in the finding of hemagglutinating activity in the processed wheat germ, peanuts and dry cereals (Table 1). Similar findings for wheat germ have been presented by Brady et al. (1978). In addition, several of the lectins have been found to be resistant to proteolytic digestion e.g., wheat germ agglutinin (Brady et al., 1978), tomato lectin (Nachbar et al., 1980), navy bean lectin (Pusztai et al., 1979) and when looked for, have been recovered intact in stool (Liener, 1974; Pusztai et al., 1979; Brady et al., 1978). Ingestion of lectins also has major cholecystokinin-mediated effects on gastrointestinal Unless otherwise stated all lectins listed in Table 1 agglutinate human erythrocytes of A, B and 0 blood types. N.T: Not tested, N.I: Activity not inhibited by battery of simple sugars function and growth.
| Table 1: | Agglutinating activities of lectins in the edible parts of various foodstuffs |
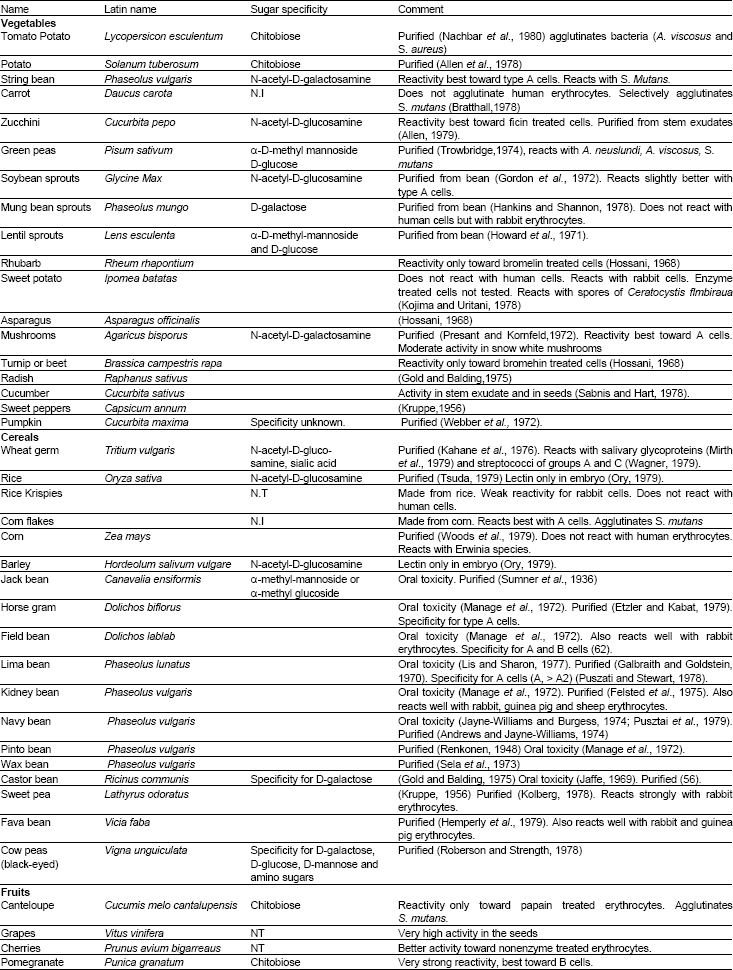 | |
| Table 1: | Continue |
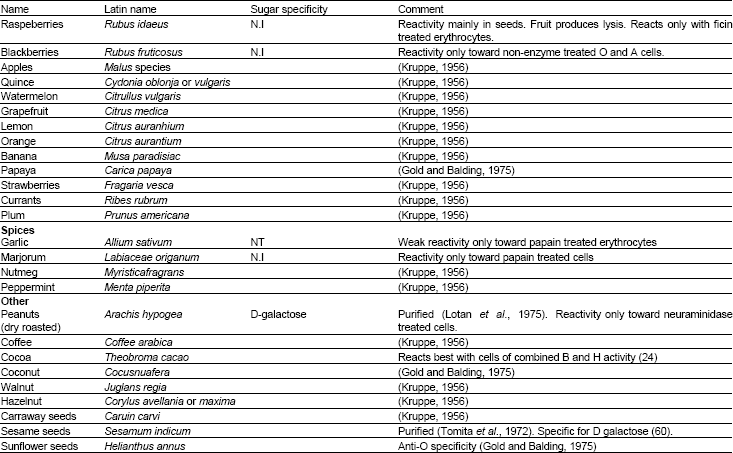 | |
It can be said that at least some lectins in foodstuffs will survive one or both degradative processes to interact with cells, secretions and microflora of the digestive tract resulting in, as yet unknown, functional consequences. Given the significant exposure of the populace to dietary lectins and the unusual breadth of biological activities potentially affected, it is obvious that future investigations of their nutritional effects will have to encompass a wider spectrum of functional parameters than heretofore tested.
Lectins in diseases of gut: That lectins cause diseases is not surprising because diet is one part of the environment that is manipulable and because lectins have excellent antidotes, at least in vitro. Because of their precise carbohydrate specificities, lectins can be blocked by simple sugars and oligosaccharides. Wheat lectin, for example, is blocked by the sugar N-acetyl glucosamine and its polymers (Goldstein and Poretz, 1986). These natural compounds are potentially exploitable as drugs should lectin induced diseases be identified. Wheat gliadin, which causes coeliac disease, contains a lectin like substance that binds to human intestinal mucosa (Kolberg and Sollid, 1985) and this has been debated as the "coeliac disease toxin" for over 30years (Weiser and Douglas, 1976). But coeliac disease is already managed by gluten avoidance, so nothing would change were the lectin hypothesis proved. On the other hand, wheat lectin also binds to glomerular capillary walls, mesangial cells and tubules of human kidney and (in rodents) binds IgA and induces IgA mesangial deposits. This suggests that in humans IgA nephropathy might be caused or aggravated by wheat lectin; indeed a trial of gluten avoidance in children with this disease reported reduced proteinuria and immune complex levels (Coppo et al., 1992). It has also been shown that local (gut) and systemic metabolism of rats is altered by consumption of raw bean (Phaseolus vulgaris L. var. athropurpurea) and these changes have been attributed to be probably mediated primarily by lectins in raw bean diet (De Moya et al., 2003). In another study the main antinutritive factors found in kidney beans were found to be generally the lectins, which were shown to interfere with gut and systemic metabolism and impair growth. In particular, they induced hyperplasia and hypertrophy of the small intestine and alterations in organ weights and function in rats (Marzo et al., 2002). At high dietary levels, lectins cause severe damage to the structure of the brush borders of the small intestine (King et al., 1986; Pusztai et al., 1988). Among the effects observed in the small intestine of lectin fed rodents is stripping away of the mucous coat to expose naked mucosa and overgrowth of the mucosa by abnormal bacteria and protozoa (Banwell et al.,1988).
Lectins also cause discharge of histamine from gastric mast cells (Greer and Pusztai, 1985) which stimulates acid secretion. So, the three main pathogenic factors for peptic ulcer, namely acid stimulation, failure of the mucous defense layer and abnormal bacterial proliferation (Helicobacter pylori) are all theoretically linked to lectins. If true, blocking these effects by oligosaccharides would represent an attractive and more physiological treatment for peptic ulcer than suppressing stomach acid. The mucus stripping effect of lectins (Freed and Buckley, 1978) also offers an explanation for the anecdotal finding of many allergists that a "stone age diet," which eliminates most starchy foods and therefore most lectins, protects against common upper respiratory viral infections: without lectins in the throat the nasopharyngeal mucus lining would be more effective as a barrier to viruses. Epithelial cells lining the GI tract in vivo, unlike cells in vitro, are constantly exposed to mechanical stress and consequently, frequently suffer plasma membrane disruptions (McNeil and Ito, 1989). However, cell death is not the only outcome of this type of injury: cells are capable of rapidly repairing and thereby surviving plasma membrane disruptions (McNeil and Kirchhausen, 2005). One key step of the repair mechanism, reviewed by Steinhardt, 2005 is exocytotic (Steinhardt et al., 1994). For large disruptions, this exocytotic reaction functions by adding a ‘patch’ of intracellular membrane to plasma membrane surrounding the disruption site (Terasaki et al., 1997).
Therefore, the mechanism of lectin toxicity in vivo could be due to an inhibitory effect on the exocytosis-based, constitutive membrane repair and consequent death of wounded gut epithelial cells. It has been found that those lectins that bind strongly to the cell surfaces potently inhibit repair (Miyake et al., 2007). Lectins are therefore toxic when a cell exposed to them experiences plasma membrane disruption. Since, mucus secretion is evoked by membrane disruptions in mucus producing GI tract cells (Miyake et al., 2006), it therefore seems likely that lectins also block secretion of this important lubricant. It was found in cell culture studies that a lectin (WGA) potently blocked ionophore-induced mucus secretion. Moreover, the copious mucus secretion normally observed from the cells when wounded, was dramatically reduced after treatment with WGA, while the dead cells were strikingly increased in density along the injury site. Thus, lectins also inhibit repair of resident gut epithelial cells and additionally, as the study showed, inhibit secretion of mucus by the cell subpopulation (goblet cells) that produces this lubricant. The mechanism of lectin inhibition of membrane repair remains unclear. Exocytosis, which is required for membrane repair (Steinhardt, 2005), is likely targeted by this class of toxin. Previous studies have shown that lectins can inhibit exocytosis (Boehm and Huck, 1998) and it has been shown that, in particular, mucin exocytosis, which is coupled to repair in the cells, is potently inhibited by lectins. Moreover, it has also been found that inhibition of repair is rapid in onset and rapidly reversed by lectin wash off (< 5 min after rinsing of cells with lectin-free medium containing competitive sugar). Therefore, inhibition is dependent on surface binding and not on other, longer-term events (minutes-hours) such as lectin-induced cell vacuolation (Edelson and Cohn, 1974). This inhibition of membrane repair also leads to diseases other than those in the gut. At least one form of muscular dystrophy is caused by a failure in the membrane repair mechanism (Bansal et al., 2003). It has been shown here that binding of cell surface glycoproteins by lectins interferes with the exocytotic events associated with membrane repair (mucus secretion) and that lectins potently block repair. Therefore, it has been proposed that a second category of disease can also be explained as a failure in the plasma membrane repair mechanism. Lectins, it is hypothesized, are toxic when present in the GI tract based on two, inter-related effects. First, resealing failure occurs within the general population of GI tract cells normally exposed to membrane disrupting levels of mechanical stress, leading to their necrosis. The second lectin-induced effect is exocytotic failure within the subpopulation of GI tract cells that normally secrete mucus, leading to a decrease in protective, lubricating mucus secretion and a consequent increase in the incidence of mechanically-induced membrane disruption events. Because lectins, based on the damage they do to the lining of the GI tract and their hypertrophic effect, have been implicated in, respectively, celiac disease (Weiser and Douglas, 1976) and cancer (Evans et al., 2002), knowledge of this mechanism may have implications beyond a better understanding of food poisoning.
Lectins in autoimmune disease: Of particular interest is also the implication of lectins for autoimmune diseases. Lectins stimulate class II HLA antigens on cells that do not normally display them, such as pancreatic islet and thyroid cells. The islet cell determinant to which cytotoxic autoantibodies bind in insulin dependent diabetes mellitus is the disaccharide N-acetyl lactosamine (Uchigata et al., 1987) which must bind tomato lectin if present and probably also the lectins of wheat, potato and peanuts. This would result in islet cells expressing the class II HLA antigens and foreign antigen together, a sitting duck for autoimmune attack. Certain foods (wheat, soya) are indeed diabetogenic in genetically susceptible mice (Scott and Kolb, 1996). Insulin dependent diabetes therefore is another potential lectin disease and could possibly be prevented by prophylactic oligosaccharides. Another suspect lectin disease is rheumatoid arthritis. The normal human IgG molecule possesses carbohydrate side chains, which terminate with galactose. In rheumatoid arthritis much of the galactose is missing, so that the subterminal sugar N-acetyl glucosamine is exposed instead. These deficient IgG molecules feature strongly in the circulating immune complexes that cause fever and other symptoms (Bond et al., 1995). In diet responsive rheumatoid arthritis one of the commonest trigger foods is wheat and wheat lectin is specific for N-acetyl glucosamine the sugar that is normally hidden but exposed in rheumatoid arthritis. This suggests that N-acetyl glucosamine oligomers such as chitotetraose (derived from the chitin that forms crustacean shells) might be an effective treatment for diet associated rheumatoid arthritis. Interestingly, the health food trade has already siezed on N-acetyl glucosamine as an anti-arthritic supplement (Toohey, 1997). There is also evidence for how the interaction of dietary lectins with enterocytes and lymphocytes may facilitate the translocation of both dietary and gut-derived pathogenic antigens to peripheral tissues, which in turn causes persistent peripheral antigenic stimulation. In genetically susceptible individuals, this antigenic stimulation may ultimately result in the expression of overt rheumatoid arthritis (RA) via molecular mimicry, a process whereby foreign peptides, similar in structure to endogenous peptides, may cause antibodies or T-lymphocytes to cross-react with both foreign and endogenous peptides and thereby break immunological tolerance. By eliminating dietary elements, particularly lectins, which adversely influence both enterocyte and lymphocyte structure and function, it is proposed that the peripheral antigenic stimulus (both pathogenic and dietary) will be reduced and thereby result in a diminution of disease symptoms in certain patients with RA (Cordain et al., 2000). Dietary lectins, present in beans and other edible plant products, pose a potential threat due to their capacity to induce histamine release from basophils and several other dietary lectins can trigger human basophils to release IL-4 and IL-13. Since lectins can enter the circulation after oral uptake, they might play a role in inducing the so-called early IL-4 required to switch the immune response towards a Th2 response and typeI allergy which leads to hypersensitivity responses in certain individuals (Haas et al., 1999).
Lectins and blood type diet: Another aspect of food lectin toxicity stems from “The blood type diet” theory advocated by D'Adamo (1996). Its basic premise is that ABO blood type is the most important factor in determining a healthy diet. Many dieticians, physicians and nutritional scientists claim the theory lacks scientific evidence. The cornerstone of this theory is that lectins in foods react differently with each ABO blood type and according to this hypothesis "lectins" which interact with the different ABO type "antigens" are described as incompatible and harmful and therefore the selection of different foods for A, AB, B and O types to minimize reactions with these lectins. D'Adamo bases his ideas on the ABO classification of Karl Landsteiner and Jan Jansky and some of the many other tissue surface antigens and classification systems, in particular the Lewis antigen system for ABH secretor status. Although, it has been claimed that there are many ABO specific lectins in foods (D'Adamo, 1996) this claim is, for a number of cited cases, unsubstantiated by established biochemical research, which has not found differences in how the lectins react with a given human ABO type. A common criticism is that lectins which are preferential for a particular ABO type are not found in foods (except for one or two rare exceptions, e.g. lima bean) and that lectins with ABO specificity are more frequently found in non-food plants or animals (Els et al., 1998; Nachbar and Oppenheim, 1980; Sharon et al., 2000).
Despite these criticisms there does seem to be a correlation between diet and blood type as a chemical reaction occurs between the blood and the foods that one eats and this reaction is part of genetic inheritance. It is amazing but true that today, in the twenty first century, our immune and digestive systems still maintain favoritism for foods that our blood type ancestors ate. Lectins, abundant and diverse proteins found in foods, have agglutinating properties that affect our blood. They are a powerful way for organisms to attach themselves to other organisms in nature. Lots of germs and even the immune systems, have used this superglue to their benefit. For example, cells in the liver’s bile ducts have lectins on their surfaces to help them snatch up bacteria and parasites. Bacteria and other microbes have lectins on their surfaces, as well, which work rather like suction cups, so they can attach to the slippery mucosal lining of the body.
When food containing protein lectins that are incompatible with one’s blood type antigen are eaten, the lectins target an organ or bodily systems (kidney, liver, brain, stomach, etc) and begin to agglutinate blood cells in that area. Many food lectins have characteristics that are close enough to a certain blood type antigen to make it an ‘enemy’ to another. For example, milk has B-like qualities, if a person with Type A blood drinks it, his/her system will immediately start the agglutinating process and reject it. Lectins therefore, on one hand agglutinate with other germs and harmful invaders for proper removal by the immune systems as happens when cancer cells or a population of pathogens force to stick together for better disposal by natural defense system. On the other hand food that is rendered beneficial for one blood type may be an ‘enemy’ to other blood type. Agglutination in long term causes chronic problems such as diseases and complications to organs late in life.
But if we all eat lectins, why don't we all get insulin dependent diabetes, rheumatoid arthritis, IgA nephropathy and peptic ulcers? Partly because of biological variation in the glycoconjugates that coat our cells and partly because these are protected behind a fine screen of sialic acid molecules, attached to the glycoprotein tips (Uchigata et al., 1987). But the sialic acid molecules can be stripped off by the enzyme neuraminidase, present in several micro-organisms such as influenza viruses and streptococci. This may explain why diabetes and rheumatoid arthritis tend to occur as sequelae of infections. This facilitation of lectins by micro-organisms throws a new light on post infectious diseases and makes the folklore cure of fasting during a fever seem sensible. Alternative medicine popularisers are already publishing articles about dietary lectins (D'Adamo, 1998), often with more enthusiasm than caution, so patients are starting to ask about them and doctors need to be armed with facts. The same comment applies to entrepreneurs at the opposite end of the commercial spectrum. Many lectins are powerful allergens and prohevein, the principal allergen of rubber latex, is one. It has been engineered into transgenic tomatoes for its fungistatic properties (Lee and Raikhel, 1995). So, we can expect an outbreak of tomato allergy in the near future among latex sensitive individuals.
Protecting against lectins: Because lectins are so prevalent in a typical diet, undertaking a supplement regimen to help combat the damaging effects of lectins can help contribute to optimal health, improve the health of the intestinal tract and contribute to weight loss. Certain seaweeds, especially those high in the sugar fucose and mucilaginous vegetables like okra have the ability to bind to lectins in a way that makes them unavailable to the vulnerable cells of the gut. These foods act as sacrificial decoys and attach to the problematic lectins that would ordinarily attach and bind to gut epithelial cells. A specific glycoprotein, N-Acetylglucosamine (NAG), is also a favorite target for dietary lectins and is concentrated in connective tissue. Supplementation with NAG is an excellent strategy for lectin protection. Another sugar with similar activity is D-mannose, which is capable of binding to lectins located on the cells of microorganisms. Some bacteria responsible for urinary tract infections contain lectins specific for the sugar mannose and use these lectins to bind tightly to mannose-rich tissue in the bladder walls, initiating urinary tract infections (Lenard and Wright, 2003). As with fucose and NAG, supplementation with D-mannose provides a decoy for these lectins and protects the bladder. Supplementing prior to a meal with these decoy sugars allows for the binding of potentially harmful lectins and protection from attack. This concept of lectin-shielding devices has exciting clinical application now and in the future (Rudiger et al., 2000).
Conclusion: The subject of lectins is very broad and deserves more discussion. There are even some lectins that are beneficial to the body, such as those found in some species of edible snails, which may be capable of preventing the metastasis of cancer cells (Schumacher et al., 1994). The involvement of lectins in our health and their relationship to degenerative disease is still an emerging science. Studies performed on animals will continue to be the model in the future for the study of lectins. The glycosylation of the human gut is basically similar to that of higher animals and it may be confidently predicted that the effects of dietary lectins will have similarities in both humans and animals. In short, dietary lectins, by their chemical reactivity with cell surface receptors on the intestinal epithelium, are metabolic signals for the gut and are capable of modulating immune and hormone functions (Pusztai, 1993). Studies in laboratory animals have shown that ingested lectins have a wide range of effects that might be relevant to human diseases. These include changes in the differentiation (Jordinson et al., 1996) as well as the proliferation of intestinal and colonic cells (Ryder et al., 1994a, b). Dietary lectins may also affect the intestinal flora (Pusztai et al., 1993) and bacterial lectins in turn can activate intestinal cells (Grant et al., 1995) which might be an important contributory factor in the prevalence of many diet associated diseases.
REFERENCES
- Allen, A.K., N.N. Desai, A. Neuberger and J.M. Creeth, 1978. Properties of potato lectin and the nature of its glycoprotein linkages. Biochem. J., 171: 665-674.
PubMedDirect Link - Bansal, D., K. Miyake, S.S. Vogel, S. Groh and C.C. Chen, 2003. Defective membrane repair in dysferlin-deficient muscular dystrophy. Nature, 423: 168-172.
PubMedDirect Link - Boehm, S. and S. Huck, 1998. Presynaptic inhibition by concanavalin: Are alphalatrotoxin receptors involved in action potential-dependent transmitter release? J. Neurochem., 71: 2421-2430.
PubMedDirect Link - Brady, P.G., A.M. Vannier and J.G. Banwell, 1978. Identification of the dietary lectin, wheat germ agglutinin, in human intestinal contents. Gastroenterology, 75: 236-239.
PubMedDirect Link - Cordain, L., L. Toohey, M.J. Smith and M.S. Hickey, 2000. Modulation of immune function by dietary lectins in rheumatoid arthritis. Br. J. Nutr., 83: 207-217.
PubMedDirect Link - De Moya, C.C., G. Grant, G. Fruhbeck, E. Urdaneta, M. Garcia, F. Marzo and S. Santidrian, 2003. Local (gut) and systemic metabolism of rats is altered by consumption of raw bean Phaseolus vulgaris, L. var. athropurpurea. Br. J. Nutr., 89: 311-318.
Direct Link - Dickinson, A.G., H. Fraser, I. McConnell and G.W. Outram, 1978. Mitogenic stimulation of the host enhances susceptibility to scrapie. Nature, 272: 54-55.
CrossRefDirect Link - Edelson, P.J. and Z.A. Cohn, 1974. Effects of concanavalin A on mouse peritoneal macrophages: Stimulation of endocytic activity and inhibition of phagolysosome formation. J. Exp. Med., 140: 1364-1386.
Direct Link - Elson, C.O., J.A. Heck and J.A.S. Ban, 1979. T-cell regulation of murine IgA synthesis. J. Exp. Med., 149: 632-643.
Direct Link - Evans, R.C., S. Fear, D. Ashby, A. Hackett and E. Williams, 2002. Diet and colorectal cancer: An investigation of the lectin-galactose hypothesis. Gastroenterology, 122: 1784-1792.
Direct Link - Forsdyke, D.R. and C.M. David, 1978. Comparison of enhancement by heated serum and 2-mercaptoethanol of lymphocyte transformation induced by high concentrations of concanavalin A. Cell Immunol., 36: 86-96.
PubMedDirect Link - Galbraith, W. and I.J. Goldstein, 1970. Phytohema-gglutinins: A new class of metalloproteins, Isolation, purification and some properties of the lectin from Phaseolus lunatus. FEBS Lett., 9: 197-201.
Direct Link - Grant, G., S. Bardocz, S.W. Ewen, D.S. Brown and T.J. Duguid(et al)., 1995. Purified Pseudomonas aeruginosa PA-I lectin induces gut growth when orally ingested by rats. FEMS Immunol. Med. Microbiol., 11: 191-195.
Direct Link - Greer, F. and A. Pusztai, 1985. Toxicity of kidney bean (Phaseolus vulgaris) in rats: Changes in intestinal permeability. Digestion, 32: 42-46.
CrossRefDirect Link - Hankins, C.N. and L.M. Shannon, 1978. The physical and enzymatic properties of a phytohemagglutinin from mung beans. J. Biol. Chem., 253: 7791-7797.
PubMedDirect Link - Haas, H., F.H. Falcone, G. Schramm, K. Haisch and B.F. Gibbs(et al)., 1999. Dietary lectins can induce in vitro release of IL-4 and IL-13 from human basophils. Eur. J. Immunol., 29: 918-927.
Direct Link - Hart, C.A., R.M. Batt, J.R. Saunders and B. Getty, 1988. Lectin-induced damage to the enterocyte brush border: An electron-microscopic study in rabbits. Scand. J. Gastroenterol., 23: 1153-1159.
Direct Link - Howard, I.K., H.J. Sage, M.D. Stein, N.M. Young, M.A. Leon and D.F. Dyckes, 1971. Studies on a phytohemagglutinin from the lentil. II. Multiple forms of Lens culinaris hemagglutinin. J. Biol. Chem., 246: 1590-1595.
PubMedDirect Link - Kahane, I., H. Furthmayr and V.T. Marchesi, 1976. Isolation of membrane glycoproteins by affmity chromatography in the presence of detergents. Biochirn. Biophys. Acta, 426: 464-476.
PubMedDirect Link - Kilpatrick, D.C., 1999. Mechanisms and assessment of lectin-mediated mitogenesis. Mol. Biotechnol., 11: 55-65.
PubMedDirect Link - King, T.P., A. Pusztai, G. Grant and D. Slater, 1986. Immunogold localization of ingested kidney bean Phaseolus vulgaris lectins in epithelial cells of the rat small intestine. Histochem. J., 18: 413-420.
CrossRefDirect Link - Kolberg, J. and L. Sollid, 1985. Lectin activity of gluten identified as wheat germ agglutinin. Biochem. Biophys. Res. Commun., 130: 867-872.
PubMedDirect Link - Kolberg, J., 1978. Isolation and partial characterization of a mitogenic lectin from Lathyrus odoratus seeds. Acta Pathol. Microbiol. Scand. Sect. C, 86: 99-104.
PubMedDirect Link - Lawrence, D.A. and R.F. Schell, 1977. Differential effects of concanavalin A and phytohemagglutinin on murine immunity: Suppression and enhancement of humoral immunity. Cell. Immunol., 31: 155-162.
PubMedDirect Link - Lee, H.I. and N.V. Raikhel, 1995. Prohevein is poorly processed but shows enhanced resistance to a chitin-binding fungus in transgenic tomato plants. Braz. J. Med. Biol. Res., 28: 743-750.
PubMedDirect Link - Liener, I.E., 1974. Phytohemagglutinins: Their nutritional significance. J. Agric. Food Chem., 22: 17-22.
CrossRefDirect Link - Lorenzsonn, V. and W.A. Olsen, 1982. In vivo responses of rat intestinal epithelium to intraluminal dietary lectins. Gastroenterology, 82: 838-848.
PubMedDirect Link - Lorna, L. and F.R.S.H. McPherson, 1990. The effect of the consumption of red kidney beans Phaseolus vulgaris on the growth of rats and the implications for human populations. J. R. Soc. Promotion Health, 110: 222-226.
CrossRefDirect Link - Lotan, R., E. Skutelsky, D. Danon and N. Sharon, 1975. The purification, composition and specificity of the anti-T lectin from peanut Arachis hypogaea. J. Biol. Chem., 250: 8518-8523.
PubMedDirect Link - Marzo, F., R. Alonso, E. Urdaneta, F.J. Arricibita and F. Ibanez, 2002. Nutritional quality of extruded kidney bean (Phaseolus vulgaris L. var. Pinto) and its effects on growth and skeletal muscle nitrogen fractions in rats. J. Anim. Sci., 80: 875-879.
Direct Link - McNeil, P.L. and S. Ito, 1989. Gastrointestinal cell plasma membrane wounding and resealing in vivo. Gastroenterology, 96: 1238-1248.
PubMedDirect Link - McNeil, P.L. and T. Kirchhausen, 2005. Opinion: An emergency response team for membrane repair. Nat. Rev. Mol. Cell Biol., 6: 499-505.
CrossRefDirect Link - Mirth, D.B., C.J. Miller, A. Kingman and W.H. Bowen, 1979. Inhibition of saliva-induced aggregation of Streptococcus mutans by wheat germ agglutinin. Caries Res., 13: 121-131.
PubMedDirect Link - Mitchell, G.F. and A.E. Clarke, 1979. Allergenicity of concanavalin A in mice. Int. Arch. Allergy Applied Immun., 58: 391-401.
CrossRefDirect Link - Miyake, K., T. Tanaka and P.L. McNeil, 2006. Disruption-induced mucus secretion: Repair and protection. PloS Biol., 4: 276-276.
Direct Link - Miyake, K., T. Tanaka and P.L. McNeil, 2007. Lectin-Based Food Poisoning: A new mechanism of protein toxicity. PLoS One, 2: 687-687.
CrossRefDirect Link - Nachbar, M.S. and J.D. Oppenheim, 1980. Lectins in the United States diet a survey of lectins in commonly consumed foods and a review of the literature. Am. J. Clin. Nutr., 33: 2338-2345.
Direct Link - Nachbar, M.S., J.D. Oppenheim and J.O. Thomas, 1980. Lectins in the U.S. diet: Isolation and characterization of a lectin from the tomato Lycopersicon esculentum. J. Biol. Chem., 255: 2056-2061.
Direct Link - Presant, C.A. and S. Kornfeld, 1972. Characterization of the cell surface receptor for the Agaricus bisporus hemagglutinin. J. Biol. Chem., 247: 6937-6945.
Direct Link - Pusztai, A. and J.C. Stewart, 1978. Isolectins of Phaseolus vulgaris Physicochemical studies. Biochim. Biophys. Acta, 536: 38-49.
PubMedDirect Link - Pusztai, A., Clarke, E.M.W. and T.P. King, 1979. The nutritional toxicity of Phaseolus vulgaris lectins. Proc. Nutr. Soc., 38: 115-120.
PubMedDirect Link - Pusztai, A., J.T.A. Oliveira, S. Bardocz, G. Grant and H.M. Wallace, 1988. Dietary Kidney bean Lectin-Induced Hyperplasia and Increases Polyamine Content of the Small Intestine. In: Lectins Biology Biochemistry and Clinical Biochemistry, Hansen, T.C.B. and D.L.J. Freed (Ed.). Sigma Library, St. Louis, pp: 117-120.
- Pusztai, A., 1993. Dietary lectins are metabolic signals for the gut and modulate immune and hormonal functions. Eur. J. Clin. Nutr., 47: 691-699.
PubMedDirect Link - Rogers, A.H., 1979. Biological methods of dental caries prevention-review. Aust. Dent. J., 24: 153-158.
PubMedDirect Link - Rudiger, H., H.C. Siebert, D. Solis, J.J. Barbero and A. Romero(et al)., 2000. Medicinal chemistry based on the sugar code: Fundamentals of lectinology and experimental strategies with lectins as targets. Curr. Med. Chem., 7: 389-416.
Direct Link - Ryder, S.D., N. Parker, D. Ecclestone, M.T. Haqqani and J. Rhodes, 1994. Peanut lectin stimulates proliferation of colonic explants from patients with inflammatory bowel disease and colon polyps. Gastroenterology, 106: 117-124.
PubMedDirect Link - Ryder, S.D., J.A. Smith, E.G.H. Rhodes, N. Parker and J.M. Rhodes, 1994. Proliferative responses of HT29 and Caco-2 human colorectal cancer cells to a panel of lectins. Gastroenterology, 106: 85-93.
PubMedDirect Link - Sabnis, D.D. and J.W. Hart, 1978. Isolation and some properties of a lectin (haemagglutinin) from Cucurbita phloem exudates. Planta, 142: 97-101.
CrossRefDirect Link - Sauvion, N., C. Nardon, G. Febvay, A.M. Gatehouse and Y. Rahbe, 2004. Binding of the insecticidal lectin concanavalin A in pea aphid, Acyrthosiphon pisum (Harris) and induced effects on the structure of midgut epithelial cells. J. Insect Physiol., 50: 1137-1150.
Direct Link - Schell, R.F. and D.A. Lawrence, 1977. Differential effects of concanavalin A and phytohemagglutinin on murine immunity: Suppression and enhancement of cell-mediated immunity. Cell. lmmunol., 31: 142-154.
PubMedDirect Link - Schumacher, U., D. Higgs, M. Loizidou, R. Pickering, A. Leathem and I. Taylor, 1994. Helix pomatia agglutinin binding is a useful prognostic indicator in colorectal carcinoma. Cancer, 74: 3104-3107.
Direct Link - Sela, B.A., H. Lis, N. Sharon and L. Sachs, 1973. Isolectins from wax bean with differential agglutination of normal and transformed mammalian cells. Biochim. Biophys. Acta, 310: 273-277.
PubMedDirect Link - Sharon, N. and H. Lis, 2004. History of lectins from hemagglutinins to biological recognition molecules. Glycobiology, 14: R53-R62.
CrossRefDirect Link - Siraganian, R.P. and P.A. Siraganian, 1975. Mechanism of action of concanavalin A on human basophils. J. Immunol., 114: 886-893.
Direct Link - Steinhardt, R.A., 2005. The mechanisms of cell membrane repair: A tutorial guide to key experiments. Ann. N.Y. Acad. Sci., 1066: 152-165.
PubMedDirect Link - Sumner, J.B., S.F. Howell and A. Zeissig, 1935. Concanavalin A and hemagglutination. Science, 82: 65-66.
CrossRefDirect Link - Steinhardt, R.A., G. Bi and J.M. Alderton, 1994. Cell membrane resealing by a vesicular mechanism similar to neurotransmitter release. Science, 263: 390-393.
CrossRefDirect Link - Trowbridge, I.S., 1973. Mitogenic properties of pea lectin and its chemical derivatives. Proc. Nat. Acad. Sci. USA., 70: 3650-3654.
Direct Link - Tsuda, M., 1979. Purification and characterization of a lectin from rice bran. J. Biochem., 86: 1451-1461.
PubMedDirect Link - Vasconcelos, I.M. and J.T. Oliveira, 2004. Antinutritional properties of plant lectins. Toxicon, 44: 385-403.
PubMedDirect Link - Wagner, M., 1979. Interaction of wheat-germ agglutinin with streptococci and streptococcal cell wall polymers. Immunobiology, 156: 57-64.
PubMedDirect Link - Weiser, M.M. and A.P. Douglas, 1976. An alternative mechanism for gluten toxicity in coeliac disease. The Lancet, 307: 567-569.
CrossRefDirect Link - Willoughby, W.F., J.B. Willoughby, B.B. Cantrell and R. Wheelis, 1979. In vivo responses to inhaled proteins. II: Induction of interstitial pneumomtis and enhancement of immune complex-mediated alveolitis by inhaled concanavalin A. Lab. Invest., 40: 399-414.
PubMedDirect Link - Zang, J., D. Li, X. Piao and X. Tang, 2006. Effects of soybean agglutinin on body composition and organ weights in rats. Arch. Anim. Nutr., 60: 245-253.
Direct Link