
Gautam Patra
Department of Veterinary Parasitology, College of Veterinary Sciences and A.H., Central Agricultural University, Aizawl, Mizoram, India
M. Ayub Ali
Department of Veterinary Biochemistry and Physiology, College of Veterinary Sciences and A.H., Central Agricultural University, Aizawl, Mizoram, India
Kh. Victoria Chanu
Department of Veterinary Biochemistry and Physiology, College of Veterinary Sciences and A.H., Central Agricultural University, Aizawl, Mizoram, India
Jonathan Lalsiamthara
SRF-RCRL, College of Veterinary Sciences and A.H., Central Agricultural University, Aizawl, Mizoram, India
J.L. Kataria
SRF-RCRL, College of Veterinary Sciences and A.H., Central Agricultural University, Aizawl, Mizoram, India
Snigdha Hazarika
Department of Veterinary Pharmacology and Toxicology, College of Veterinary Sciences and A.H., Central Agricultural University, Aizawl, Mizoram, India
David Malsawmkima
Department of Veterinary Microbiology, College of Veterinary Sciences and A.H., Central Agricultural University, Aizawl, Mizoram, India
R. Ravindran
Department of Veterinary Pathology, College of Veterinary Sciences and A.H., Central Agricultural University, Aizawl, Mizoram, India
L. Inaotombi Devi
Department of Medical Laboratory Technology, RIPANS, Aizawl, Mizoram, India
Research Journal of Parasitology
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 43-52







ABSTRACT
An outbreak of coccidiosis in laboratory rats has been described in this study. Clinically, out of 50 rats, 30 were died after showing symptoms of anorexia, emaciation and diarrhea. Post mortem examination showed the large and small intestine distended with necrotic and sloughed off mucosal epithelial cells with intestinal content. The examination of the gut contents revealed numerous Eimeria oocysts. After sporulation, the oocysts were identified as those of Eimeria nieschulzi. The infection due to Eimeria nieschulzi is confirmed by Polymerase Chain Reaction (PCR) after amplifying partial (~1.6 kb) nuclear 18S rDNA from DNA of E. nieschulzi. The histopathological examination showed severe damage of the lamina propria of the intestinal mucosa with numerous coccidian developmental stages in the epithelium of small intestine. Biochemical analysis of serum from infested rats presented a significant increase in serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST), glucose, total proteins, globulin and inorganic phosphorous while a decrease in alkaline phosphatase and blood urea nitrogen with insignificant changes in potassium, sodium, magnesium and chloride.
PDF Abstract XML References Citation
Received: September 15, 2010;
Accepted: December 08, 2010;
Published: February 07, 2011
How to cite this article
Gautam Patra, M. Ayub Ali, Kh. Victoria Chanu, Jonathan Lalsiamthara, J.L. Kataria, Snigdha Hazarika, David Malsawmkima, R. Ravindran and L. Inaotombi Devi, 2011. Molecular Diagnosis of Naturally Infection with Eimeria nieschulzi in Laboratory Rats. Research Journal of Parasitology, 6: 43-52.
URL: https://scialert.net/abstract/?doi=jp.2011.43.52
URL: https://scialert.net/abstract/?doi=jp.2011.43.52
INTRODUCTION
Coccidia are intracellular, obligate protozoa belonging to the phylum-Apicomplexa and are found in a wide variety of wild and domestic vertebrates (Duszynski and Upton, 2001). Coccidiosis affects the living host in many ways, depending upon the tissue preference of the particular parasite involved and the number of oocysts in the initial infection. Most of these parasites attack the mucosa of intestinal tract; therefore, symptoms are predominantly enteric. Two genera of concern are Eimeria and Isospora (Van Kruningen, 1995). Eimeria nieschulzi is an intestinal coccidian uncommon in laboratory rats but is common in wild rats. Pathogenic effects of E. nieschulzi infection are usually seen in young rats less than six months of age. Signs include diarrhea, weakness, emaciation and possibly death. Eimeria species developing in the crypts of enterocytes or in the cells lamina propria mucusa might cause sever coccidiosis (Duszynski and Upton, 2001; Slapeta et al., 2001). Eimeria nieschulzi is considered mildly pathogenic in small numbers. However, after primary infection with large number of sporulated oocysts, pathologic changes may be severe and include crypt hyperplasia, villus atrophy, increase villus to crypt ratio, cellular infiltration, oedema, hyperaemic to haemorrhagic bowel and increased mass of the small intestine. In addition to inducing pathologic lesions, E. nieschulzi infection reduces digestibility of dry matter, organic matter and nitrogen. Also, E. nieschulzi may alter the host response to concurrent or subsequent infection with other parasites (Al-Dahwi et al., 2006). Diagnosis of Eimeria infections and differentiation between species is usually attempted by consideration of clinical signs in the host and biological features of the parasites. The latter include prepatent period, site of development within the intestine and the morphological appearance of oocysts in the faeces and endogenous stages in the intestinal or caecal mucosa. The presence of parasites may be confirmed by post-mortem examination of the host or by fecal examination. Experimentally, passaging oocysts through chickens provide relevant information regarding prepatent periods and immunogenicity against reference strains (Shirley et al., 2005). However, the accurate identification of parasites (irrespective of developmental stage and sex) and their genetic characterization have important implications for systematic (taxonomy and phylogeny, population genetics, ecology and epidemiology) and also centre to diagnosis, treatment and control of the disease they cause (Gasser and Chilton, 2001).
Recently, Polymerase Chain Reaction (PCR) based amplification of DNA have been used for the diagnosis of coccidial parasites of man and animals. A number of approaches have proved to be both specific and highly sensitive for analyses either of parasites grown in vitro or present in tissue samples and clinical materials, for e.g., Toxoplasma (Burg et al., 1989; Guay et al., 1993) Neospora (Holmdahl and Mattsson, 1996; Muller et al., 1996) and Eimeria (Beate et al., 1998; Su et al., 2003; Lew et al., 2003; Kawahara et al., 2008). Although there are some reports of coccidial infection in rats (Sevinc, 2001), pathology and biochemical observation in rat coccidiosis is very rare. The current study aimed to analyze the histopathological changes in the small intestine of laboratory rats naturally infected with E. nieschulzi along with some selected biochemical parameter changes induced by the parasite.
MATERIALS AND METHODS
Of the 50 white albino outbred rats, of either sex and aged 3-4 months maintained in the college animal house, 15-20 rats started showing the symptoms of anorexia, diarrhea and weakness during the month of January, 2010. Within 3 days from the onset of these clinical symptoms, the total number of affected rats rose to 40 with mortality of 30 rats. Preliminary examination of faeces of the affected rats revealed presence of large number of unsporulated oocysts. In order to investigate and confirm the cause of mortality, faeces and blood were collected from a group of 10 rats each without clinical symptoms, with clinical symptoms and those died. Faeces were examined under microscope for coccidian oocysts and serum was separated and kept at -20°C until it was analysed for alanine amino transferees (ALT), aspertate amino transferees (AST), alkaline phosphatase (ALP), glucose, total proteins, albumin and globulin; blood urea nitrogen, inorganic magnesium, potassium, sodium, chloride and phosphorus using standard diagnostic kits available in the market (Coral Biosystem) as biochemical indicators for liver and kidney functions.
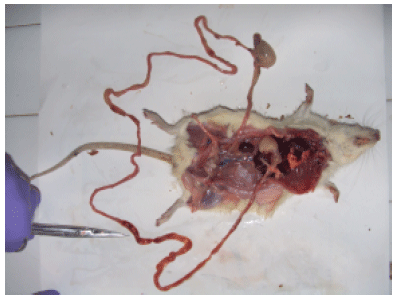 | |
| Fig. 1: | Tympanic changes in gut of laboratory rats |
The post mortem of dead rats were conducted, lesions present were recorded and a piece of intestine showing lesions were also taken from the representative rats for histopathology. Tissues collected were fixed in 10% formalin,embedded in paraffin, sections were cut at 5 μm and stained with Haematoxylin and eosin (H&E). Pooled faecal samples were also collected for sporulation of oocysts in 2.5% potassium dichromate solution. Sporulated oocysts were then identified and were also used for molecular diagnosis.
DNA extraction: The purified oocysts, stored in 2.5% potassium dichromate solution, were washed 4 times by centrifugation (1.4x103 rpm for 5 min, each wash) in autoclaved, high-salinity, phosphate buffered saline (300 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.7 mM NaH2PO4) (Reece et al., 1997). The pellet was resuspended in 200 μL of 5.75% sodium hypochlorite and incubated on ice for 30 min. The oocyst suspension was then diluted with 1 mL of sterile double distilled water and pelleted by centrifugation. The pellet was again washed 3 times with PBS. The pellet was resuspended in 1 mL deionized water and sonicated at high speed using ultrasonicator (OMNI Ruptor400, Made in USA) to disrupt the oocysts. The DNA was extracted from sonicated oocysts and analysed by agarose gel electrophoresis.
PCR amplification and identification of the PCR products: For testing nuclear DNA, the18SrDNA was amplified using primers18SF1, 5'-GCTTGTCTCAAAGATTAAGCC and 18SR2, 5'-AGCGACGGGCGGTGTGTACAA.
PCR: The PCR was done in Thermo Cycler C 1000 (BioRad, USA), with 50 μL of reaction mixtures for each sample that contained 10 mM Tris-HCl (pH 8.3), 50 mM KCl, 2 mM MgCl2, 200 μM of each dNTPs, 100 pmol (25 pmol each of species specific oligonucleotide primer of Eimeria nieschulzi and target DNA in each tube. The optimum temperature for amplification of the DNA was determined by employing different temperature from 57-62°C and subsequently suitable temperature was used for PCR amplification. The amplification procedure was formulated as 94°C for 4.5 min for initial denaturation before adding 1U of Taq DNA polymerase (Bangalore Genei). It is followed by 30 cycles each of denaturation at 94°C for 50 sec, annealing at 62°C for 50 sec and extension step at 72°C for 1 min and the final extension was performed at 72°C for 2 min. Ten-microlitre of the reaction mixtures was electrophoresed in 1.5% agarose gel having ethidium bromide and visualized in a gel documentation system (BioRad, USA).
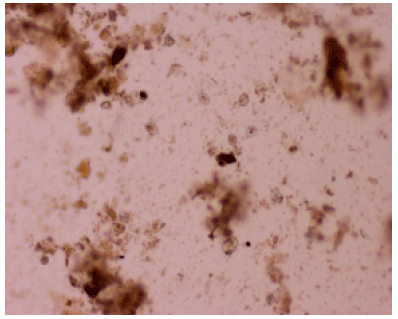 | |
| Fig. 2: | Unsporulated oocysts of Eimeria nieschulzi (10x) |
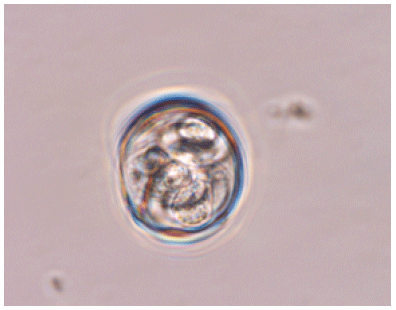 | |
| Fig. 3: | Sporulated oocyst of Eimeria nieschulzi of rat (100x) |
RESULTS
Of 50 rats reared in the college animal house, 30 out of 40 died after showing symptoms of anorexia diarrhea and emaciation. The microscopical examination of faeces content showed numerous oocysts. The sporulated oocysts were ellipsoidal to ovoid, tapering both ends. Wall was smooth, yellowish and measured 22.51-14.77 μm (on average) with no micropyle and it was identified as Eimeria nieschulzi (Fig. 2, 3).
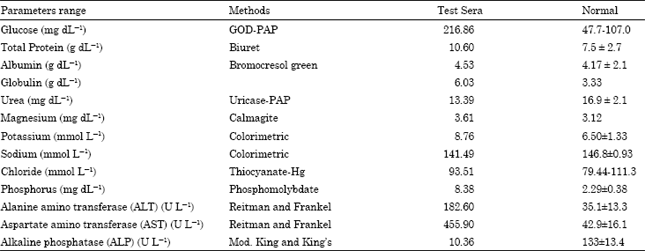
| Table 1: | Serum biochemical profile of laboratory rats infected with Eimeria nieschulzi |
 | |
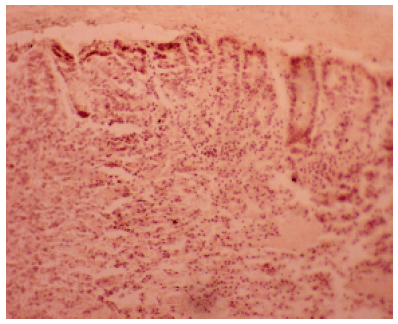 | |
| Fig. 4: | Intestinal mucosa showing cellular infiltration, sloughing of mucosa and developmental stages of Eimeria nieschulzi (10x) |
In histopathological sections, numerous coccidian developmental stages were seen in the epithelial cells. The histopathology included substantial hyperplasia of the crypt enterocytes, damage to the surface mucosa and extensive infiltration of the mononuclear cells in the lamina propria with sloughing of the mucosal epithelium (Fig. 4, 5). Based on clinical symptoms, substantiated by faecal examination and histopathological changes, it was suggesting coccidiosis to be the cause of mortality caused by Eimeria nieschulzi.
Biochemical findings: Liver function test of infested rats showed a significant increased in alanine amino transferase (ALT) and aspertate amino transferase (AST) while the levels of serum urea and alkaline phosphatase (ALP) were decreased with variation in the level of glucose, total protein, globulin and inorganic phosphorus (Table 1).
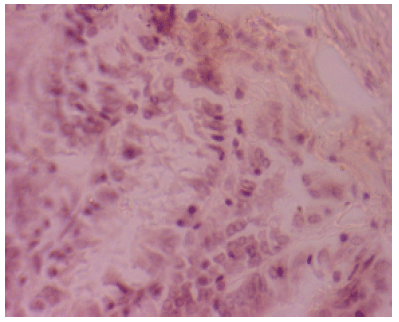 | |
| Fig. 5: | Intestinal mucosa showing cellular infiltration, sloughing of mucosa and developmental stages of Eimeria nieschulzi (40x) |
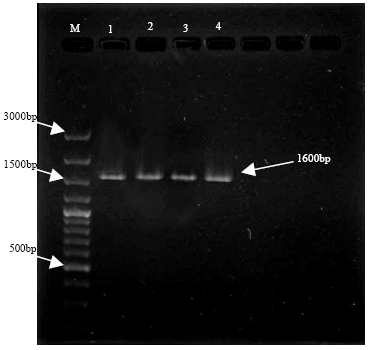 | |
| Fig. 6: | PCR results of partial (~1.6 kb) nuclear 18S rDNA amplified from DNA of E. nieschulzi |
PCR amplification: The primer pair for nuclear DNA specific for nuclear 18S rDNA of E. nieschulzi exclusively amplified their target DNA as shown in Fig. 6.
DISCUSSION
Coccidiosis is caused by a parasitic protozoan, which lives in the gut. Coccidia are obligate intracellular parasites whose development within the cytoplasm of epithelial cells results in the death of parasitized cell. When many cells of the intestinal epithelium are parasitized at one time, the denuded mucosa may bleed freely and intense inflammation involves the lamina propria and sometimes the submucosa. As large numbers of epithelial cells are destroyed the remaining epithelium is stimulated to replace that which was lost in domestic animals (Jones et al., 1997). There is little knowledge about morphology, histopathology of Eimeria sp. that caused coccidiosis in laboratory rats. Rose and Hesketh (1982) investigated infection with Eimeria nieschulzi and noted the changes in increase in gut weight, partial villous atrophy and increased numbers of mast, goblet and pyroninophilic cells. Blagburn and Todd (1984) observed histopathologic changes consisting of an early neutrophil and mononuclear cell infiltration in the small intestine and later, villus atrophy and crypt hyperplasia caused a decrease in the villus-crypt ratio in experimental Eimeria vermiformis infections in Mus musculus. These findings are similar to the observation made in the present investigation.
In the present study, high level of blood glucose has been observed in naturally infestated rats with Eimeria niestulzi. The present findings on blood glucose are in agreement with those of Uday Kumar and Padmavathi (2000) and Hirani et al. (2007). The high level of blood glucose observed in coccidian infected rats may be either due to stress condition leading to the liberation of adrenal cortisones/corticosteroids which induce hyperglycaemia or disturbed carbohydrate metabolism with interference with phosphorylative carbohydrate dissimilation by unidentified material present in intestine, as suggested by Daugherty and Herrick (1952). Marked alteration in intestinal physiology resulting in impaired carbohydrate absorption in acute phase of coccidiosis has also been reported previously (Turk et al., 1977). Waxler (1941) and Pratt (1940, 1941) have reported that in chickens infected with Eimeria tenella, hyperglycaemia develops, and there is a decrease in the amount of stored glycogen. Stein and Marquardt (1973) observed increased uptake of glucose into small intestinal tissue and hepatic portal plasma at 3 days postinoculation and decreased absorption at 8 days postinfection with experimental infection with Eimeria nieschulzi compared to uninfected control rats.
An increased in level of the total serum protein and serum globulin without a marked change in the level of serum albumin was observed in the present investigation. The reason for high level of serum total protein and globulin in the serum profiles of infested rats is due to infection by the Eimeria niestulzi. Clinico-pathologic changes in natural and/or experimental infection in rabbits with Eimeria stiedai reported by various workers (Abdel Ghaffar et al., 1990; Barriga and Arnoni, 1979, 1981; Pakes and Gerrity, 1994) and these changes include increase in β and γ globulin, β-lipoprotein, succinate dehydrogenase (which later declines), bilirubin, alanine transaminase and aspartate transaminase levels in serum. The present finding is in contrast to the findings of Stockdale et al. (1981). Stockdale et al. (1981) observed mild decrease in mean total plasma protein and mean plasma albumin levels in the calves experimentally infected with E. zuernii. Decreases in a-lipoprotein, glucose and protein levels in serum and in alkaline phosphatase activity in the liver have also been reported. The increase in the level of serum enzyme activity of ALT and AST in the present investigation is similar with the findings of Cam et al. (2008) in their experimental study of rabbits with Eimeria stiedai. A decreased in the level of serum alkaline phosphatase activity was observed in the present investigation. The decreased level of serum alkaline phosphatase in the present study is in agreement with the observations made by Abdel Ghaffar et al. (1990), Barriga and Arnoni (1979), Barriga (1981) but is in contrast to the finding of Manning et al. (1994) where they found an increase level of ALP in young rabbits (2 months old) infected with hepatic coccidiosis. Reddy et al. (1988) and Nagalakshmi et al. (1996) reported that Alkaline phosphatase level was significantly reduced in the birds infected by coccidia which is indicative of damages to the bone marrow with severe growth depression as ALP is known to rise during active growth. Similarly, there may be damage to the bone marrow of the coccidia infected rates which results in the decrease activity of the alkaline phosphatase.
Serum analysis also showed slight decreased level of sodium and increased level of potassium. This electrolyte disturbance can be attributed to diarrhea. The present finding is in agreement with the findings of Fiztgerald (1967), who noted only a minor changes in the serum sodium and potassium level in the calves infected with E. bovis. The serum sodium level decreases slightly while the level of potassium marginally increases until the infection is severe. He also noted an increase in serum potassium in severely infected calves. Veli et al. (2004) analyzed coccidian related alterations in intestinal function by ion transport and electrophysiological investigation in rats experimentally infected with Eimeria separata for possible cause of diarrhea due to coccidiosis.
From the present observations, we can conclude that the outbreak of coccidiosis may cause severe problems in the laboratory rats even under modern condition of breeding and maintenance.
REFERENCES
- Abdel Gaffar, F., M. Marzouk, M.B. Ashour and M.N. Mosaad, 1990. Effects of Eimeria labbeana and E. stiedai infection on the activity of some enzymes in the serum and liver of their hosts. Parasitol. Res., 76: 440-443.
PubMed - Barriga, O.O. and J.V. Arnoni, 1979. Eimeria stiedae: Oocyst output and hepatic function of rabbits with graded infections. Exp. Parasitol., 48: 407-414.
CrossRef - Schnitzler, B.E., P.L. Thebo, J.G. Mattsson, F.M. Tomley and M.W. Shirley, 1998. Development of a diagnotic pcr assay for the detection and discrimination of four pathogenic Eimeria Species of the chicken. Avian Pathol., 27: 490-497.
CrossRef - Blagburn, B.L. and K.S. Todd, 1984. Pathological changes and immunity associated with experimental eimeria vermiformis infections in mus musculus. J. Eukaryotic Microbiol., 31: 556-561.
CrossRef - Burg, J.L., C.M. Grover, P. Pouletty and J.C. Boothroyd, 1989. Direct and sensitive detection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chain reaction. J. Clin. Microbiol., 27: 1787-1792.
PubMedDirect Link - Cam, Y., A. Atasever, G. Eraslan, M. Kibar and O. Atalay et al., 2008. Eimeria stiedae: Experimental infection in rabbits and the effect of treatment with toltrazuril and Ivermectin. Exp. Parasitol., 119: 164-172.
CrossRefPubMedDirect Link - Daugherty, J.W. and C.A. Herrick, 1952. Cecal coccidiosis and carbohydrate metabolism in chickens. J. Parasitol., 38: 298-304.
Direct Link - Fiztgerald, P.R., 1967. Effect of bovine coccidiosis on blood serum sodium and potassium levels of calves. Am. J. Vet. Res., 28: 667-670.
PubMed - Gasser, R.B. and N.B. Chilton, 2001. Applications of Single-Strand Conformation Polymorphism (SSCP) to taxonomy, diagnosis, population genetics and molecular evolution of parasitic nematodes. Vet. Parasitol., 101: 201-213.
PubMed - Holmdahl, O.J.M. and J.G. Mattsson, 1996. Rapid and sensitive indentification of Neospora caninum by in vitro amplification of the internal transcribed spacer1. Parasitol., 112: 177-182.
PubMed - Kawahara, F., K. Taira, S. Nagai, H. Onaga, M. Onuma and T. Nunoya, 2008. Detection of five avian Eimeria species by species-specific real-time polymerase chain reaction assay. Avian Dis., 52: 652-656.
PubMed - Lew, A.E., G.R. Anderson, C.M. Minchin, P.J. Jeston and W.K. Jorgensen, 2003. Inter- and intra-strain variation and PCR detection of the internal transcribed spacer 1 (ITS-1) sequences of Australian isolates of Eimeria species from chickens. Vet. Parasitol., 112: 35-50.
PubMed - Muller, N., V. Zimmermann, B. Hentrich and G. Gotterstain, 1996. Diagnosis of Neospora canninum and Toxoplasma gondii by PCR and DNA, hybridization immuno assay. J. Cl. Microbiol., 34: 2850-2852.
Direct Link - Nagalakshmi, D., V.R.B. Sastry, D.K. Agrawal, R.C. Katigar and S.V.S. Verma, 1996. Performance of Broiler Chicks Fed alkali-treated neem kernel cake. Br. Poult. Sci., 37: 809-818.
PubMed - Pratt, I., 1940. The effect of Eimeria tenella (coccidia) upon the blood sugar of the chicken. Trans. Am. Microsc. Soc., 59: 31-37.
Direct Link - Pratt, I., 1941. The effect of Eimeria tenella (coccidia) upon the glycogen stores of the chicken. Am. J. Hyg., 34: 54-62.
Direct Link - Reece, K.S., M.E. Siddal, E.M. Burreson and J.E. Graves, 1997. Phytogenetic analysis of Perkinsus based on actin gene sequences. J. Parasitol., 83: 417-423.
PubMedDirect Link - Rose, M.E. and P. Hesketh, 1982. Coccidiosis: T-lymphocyte-dependent effects of infection with Eimeria nieschulzi in rats. Vet. Immunol. Immunopathol., 3: 499-508.
PubMed - Shirley, M.W., A.L. Smith and F.M. Tomley, 2005. The biology of avian Eimeria with an emphasis on their control by vaccination. Adv. Parasitol., 60: 285-330.
CrossRefPubMedDirect Link - Slapeta, J.R., D. Modry, J. Votypka, M.J. Jirku, M. Obornik, J. Lukes and B. Koudela, 2001. Eimeria telekiin sp. (Apicomplexa: Coccidia) from Lemniscomys striatus (Rodentia: Muridae): Morphology, pathology and phylogeny. Parasitology, 122: 133-143.
PubMed - Stein, A.S. and W.C. Marquardt, 1973. Eimeria nieschulzi: Glucose absorption in infected rats. Exp. Parasitol., 34: 262-267.
CrossRef - Stockdale, P.H.G., A.R. Bainborough, C.B. Bailey and L. Niilo, 1981. Some pathophysiological changes associated with infection of Eimeria zuernii in calves. Can. J. Comp. Med., 45: 34-37.
PubMed - Su, Y.C., A.C.Y. Fei and F.M. Tsai, 2003. Differential diagnosis of five avian Eimeria species by polymerase chain reaction using primers derived from the internal transcribed spacer 1 (ITS-1) sequence. Vet. Parasitol., 117: 221-227.
CrossRefDirect Link - Waxler, S.H., 1941. Changes occurring in the blood and tissue of chickens during coccidiosis and artificial haemorrhage. Am. J. Physiol., 134: 19-26.
Direct Link - Al-Dahwi, Z., L.F. Mayberry, G.A. Conder and J.R. Bristol, 2006. Suppression of extraintestinal and intestinal Nippostrongylus brasilliensis induced eosinophilia by Eimeria nieschulzi. J. Parasitol., 92: 962-970.
PubMed
Dr. Michael Kurth Reply
I´ve some doubts that the identified species is Eimeria nieschulzi. We work for a long time with Eimeria nieschulzi and we never saw such necrotic tissue in the small intestine and never in the large intestine. Lesions caused by Eimeria nieschulzi are normally limited on the small intestine. Further, the shown oocyst in Figure 3 is (at least) not a (typical) Eimeria nieschulzi oocyst. The histological images have unfortunatly a too poor quality to see if the developmental stages are in the villi or in the crypts. With the used primer pair it is not possible to amplify the specific 18S rDNA of Eimeria nieschulzi. By the used primers you can amplify the 18S rDNA of every Eimeria species.The PCR reaction wasn´t a proof without sequencing.