
G.K. Sahu
Department of Microbiology, College of Basic Sciences and Humanities, CCS Haryana Agricultural University, Hisar-125 004, India
S.S. Sindhu
Department of Microbiology, College of Basic Sciences and Humanities, CCS Haryana Agricultural University, Hisar-125 004, India
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 10 | Page No.: 735-749







ABSTRACT
Siderophore production by rhizosphere bacteria contributes to fungal growth inhibition resulting into suppression of plant diseases. Therefore, siderophore-producing Pseudomonas isolates were studied for root rot disease control and plant growth stimulation of green gram. Fifty-eight Pseudomonas strains were obtained from the rhizosphere soil of chickpea and green gram using dilution plating technique. These strains were tested for siderophore production on MM9 medium and succinate medium plates containing chromo-azurol S by spot test method. Siderophore-producing strains formed the halo zone around the colony growth and diameter of halo zone varied with different Pseudomonas strains. Siderophore production was found more on MM9 medium as compared to succinate medium. Bacterial colony growth (G) and halo zone (H) size ratios varied from 1.44 to 10.24 in different Pseudomonas strains on MM9 medium. Siderophore-producing Pseudomonas strains inhibited the growth of phytopathogenic fungi i.e., Fusarium oxysporum, Rhizoctonia solani and Pythium aphanidermatum. Large zones of fungal growth inhibition were observed on PDA medium in comparison to nutrient agar medium plates. Seed inoculation of Pseudomonas cultures in green gram (Vigna radiata L.) caused reduction in root rot disease (R. solani induced) symptoms under pot house conditions and disease control varied from 33.4 to 100% with different Pseudomonas cultures. Coinoculation of different Pseudomonas strains with Bradyrhizobium strain SMR15 in green gram also enhanced the nodule number, nodule fresh weight and plant dry weight at 30 and 60 days of plant growth. Coinoculation of Pseudomonas strain CP56 with Bradyrhizobium strain and R. solani showed maximum 275.8% increase in plant dry weight at 60 days in comparison to control uninoculated plants and completely suppressed the root rot disease under pot house conditions. Thus, inoculation with siderophore-producing Pseudomonas sp. could be utilized for disease control and plant growth promotion of legumes.
PDF Abstract XML References Citation
Received: April 15, 2011;
Accepted: August 28, 2011;
Published: October 28, 2011
How to cite this article
G.K. Sahu and S.S. Sindhu, 2011. Disease Control and Plant Growth Promotion of Green Gram by Siderophore Producing Pseudomonas sp. Research Journal of Microbiology, 6: 735-749.
URL: https://scialert.net/abstract/?doi=jm.2011.735.749
URL: https://scialert.net/abstract/?doi=jm.2011.735.749
INTRODUCTION
Iron is fourth most abundant element on the earth’s crust and is essential for the growth of all living organisms (Neiland, 1981). However, its availability to plants and microorganisms is hindered due to its easy chemical oxidation and formation of insoluble ferric salts (Neilands, 1982). Most of the microorganisms produce extracellular iron transport agents called siderophores, to solubilize and sequester iron (Leong, 1986; Buyer and Sikora, 1990). These siderophores play prominent role in the physiology of symbiotic diazotrophs, soil microflora and in biological control of soil borne plant pathogens (Crowley and Kraemer, 2007; Lemanceau et al., 2007).
The plant rhizosphere is colonized by saprophytic, pathogenic and plant growth promoting strains of bacteria (Miller et al., 1989). The interactions of rhizospheric microorganisms with each other and the environment surrounding the plant root, leads to either beneficial or detrimental effects on plant growth (Weyens et al., 2009; Sindhu et al., 2010; Woyessa and Assefa, 2011). Many species of Azotobacter, Azospirillum, Bacillus, Bradyrhizobium, Burkholderia, Pseudomonas, Rhizobium and Serratia have been found to improve plant growth and are termed as plant growth-promoting rhizobacteria (PGPR) (Kloepper et al., 1980). Some PGPR are also involved in control of plant diseases by suppression of phytopathogenic organisms (Weller, 2007) by various mechanisms viz. antibiosis, production of hydrolytic enzymes and hydrocyanic acid, stimulation of phytoalexins or flavonoid-like compounds in roots and by induction of systemic resistance. PGPR have also been reported to produce siderophores which chelate metal cations and inhibit the spore germination of pathogenic fungi leading to disease suppression (Raaijmakers et al., 1995; Sindhu et al., 2009). Thus, PGPR have emerged an environment friendly alternative to the hazardous pesticides.
The pathogenic fungi i.e., Fusarium oxysporum, Rhizoctonia solani and Pythium aphanidermatum have been found to cause wilt, root rot and damping off disease, respectively in various crops leading to significant losses in crop yield (Siddiqui et al., 2001; Validov et al., 2007; Scherwinski et al., 2008). Recently, seed bacterization with specific PGPR strains inhibited the growth of soil borne root infecting fungi, suppressed the disease and enhanced the growth of various plants under green house and field conditions (Ehteshamul-Haque and Ghaffar, 1993; Kumar, 1996; Ahmad et al., 2008). The fluorescent siderophores were found to control the growth of pathogenic fungi i.e., Pythium ultimum (Gill and Warren, 1988), Gaeumannomyces graminis (Brisbane and Rovira, 1988) and inhibited the germination of Fusarium oxysporum chlamydospores (Simeon et al., 1987). The inhibition was totally or partially reversed if the medium or field soil was amended with iron (III) in the form of Fe-EDTA. Siderophore negative mutants were not inhibitory to these fungi indicating that siderophores contribute to the suppression of certain fungal diseases (Buysens et al., 1996).
Cucumber and spinach seedlings supported more growth of the siderophore producing Pseudomonas strains than nonproducers and also promoted rootlet elongation on cucumber (De Bellis and Ercolani, 2001). Moreover, the coinoculation of Pseudomonas sp. with Bradyrhizobium/Mesorhizobium sp. have been found to cause a significant increase in nodule number, nodule weight and plant dry weight of green gram and chickpea when grown under sterilized chillum jar conditions (Sindhu et al., 1999, 2002). Present study was carried out to find out the prevalence of siderophores in Pseudomonas strains and to understand the role of siderophores in disease control and plant growth promotion of green gram.
MATERIALS AND METHODS
This study was carried out in the Department of Microbiology, CCS Haryana Agricultural University, Hisar on the crop green gram during the month of July, 2008 to June 2010.
Microbial cultures: Fifty-eight Pseudomonas isolates were obtained from the rhizosphere soil of chickpea and green gram by plating of serial dilutions on King’s B (KB) medium plates. Pseudomonas colonies were selected based on typical morphological and pigment production characteristics. The different Pseudomonas isolates were grown on KB medium plates for 2 days at 28+2°C. One percent solution of tetramethyl-p-phenyl-diamine dihydrochloride was added to cover surface of plates. The oxidase positive Pseudomonas isolates gave purple colour on these plates.
Twenty-two reference strains of Pseudomonas and one Bradyrhizobium sp. strain SMR15 were obtained from the Department of Microbiology, CCS Haryana Agricultural University, Hisar. The Pseudomonas strains were maintained by periodic transfer on Luria Bertani agar slants (Sambrook et al., 1989) and Bradyrhizobium strain was maintained on yeast extract mannitol agar (YEMA) medium slants (Somasegaran and Hoben, 1994). The bacterial cultures were stored at 4°C in refrigerator for further use. Three fungal cultures i.e., Fusarium oxysporum, Pythium aphanidermatum and Rhizoctonia solani were obtained from the Department of Plant Pathology, CCS Haryana Agricultural University, Hisar and maintained on potato dextrose agar (PDA) medium slants. Seeds of green gram (Vigna radiata) var. Asha were obtained from RDS Seeds Farm, CCS Haryana Agricultural University, Hisar.
Screening of Pseudomonas sp. for siderophore production: The Universal chemical assay utilizing chromo-azurol S (CAS/iron (III) and hexadecyltrimethyl ammonium bromide) agar plates was used for qualitative detection of siderophores produced by different Pseudomonas strains as described by Schwyn and Neilands (1987b). Siderophore production was also determined on iron deficient succinate medium used by Mayer and Abdallah (1978). Different Pseudomonas cultures were grown on fresh King’s B medium (King et al., 1954) slants. Growth suspension (10 μL) of different Pseudomonas strains was spotted on the CAS incorporated plates and incubated at 28±2°C for 3-4 days. Orange halo zone around the colonies on blue agar plates indicated the presence of siderophores (Schwyn and Neilands, 1987a). The diameter of orange halo zone formed and diameter of the Pseudomonas colony was measured as described by Van Rossum et al. (1994).
Growth inhibition of pathogenic fungi: Three fungi i.e., Fusarium oxysporum, Pythium aphanidermatum and Rhizoctonia solani were transferred to fresh PDA medium plates and incubated for four days at 28±2°C to get complete spore formation. The fungal spore suspension (3.0 mL) was added into sterilized nutrient agar (NA) and PDA medium, mixed uniformly and plated. Growth suspension (10 μL) of different Pseudomonas strains was spotted on fungal spore-incorporated plates as described by Sindhu et al. (1999). The plates were incubated in the BOD incubator at 28±2°C for 4 days. The growth inhibition zones of different fungi were recorded.
Seed bacterization of green gram with Pseudomonas strains: Three siderophore-producing Pseudomonas strains, i.e., MP20, MPS54 and CP56 and one siderophore-lacking strain CPS67 were tested for disease control of green gram under pot house conditions. The earthern pots of 10 kg capacity were filled with sandy loam soil and river sand mixed in 70:30 ratio. Pseudomonas strains were grown on LB medium for 2 days and a Bradyrhizobium sp. strain SMR15 was grown on YEMA medium slopes for 7 days. The growth suspension of each Pseudomonas and Bradyrhizobium cultures was made in 5 mL of sterilized water.
Surface sterilized seeds of green gram (Vigna radiata) cv. Asha were inoculated with broth culture of Bradyrhizobium strain SMR15 alone or with different Pseudomonas strains. Coinoculation of different Pseudomonas strains was done by mixing with growth suspension of Bradyrhizobium in 1:1 ratio (v/v). The viable count in the broth was kept 108-109 cells mL-1 and 10 g seeds were inoculated with 1 mL of bacterial growth suspension (Sindhu et al., 1999). There were nineteen treatments in this experiment. Each treatment had three replications. Growth of 4 days-old R. solani was harvested from PDA plates and fungal growth suspension was prepared in sterilized saline water. Fungal growth suspension (100 mL) was mixed in the 10 kg soil: sand mixture in earthern pots with treatments T2, T6, T7, T10, T11, T14, T15, T18 and T19. The growth suspension of fungus was also inoculated on the roots of green gram plants in the R. solani treatments only.
The plants were grown in the pot house under day light conditions during the month of May-June 2010. Sloger’s nutrient solution was added in the pots as and when required (Sloger, 1969). The plants were uprooted at 30 and 60 days of plant growth and observations were taken for nodule number, nodule fresh weight, plant dry weight and disease index. After washing with tap water, nodules were detached from the roots and dried in the folds of filter paper. The nodules were counted and weighed. Shoot portions of the plants were dried in oven at 90°C for 24 h and weighed.
Disease index and reduction in disease: On the basis of symptoms observed, percent disease index, percent final stand and percent disease control were calculated by following formulae:
 |
Disease control and disease incidence were recorded after 30 and 60 days of sowing. It was calculated on the average of six plants grown per pot.
RESULTS
Certain bacteria such as pseudomonas and bacilli exert beneficial effects on plants either by enhancing crop nutrition, by reducing damages caused by pathogens or pests and by their ability to degrade xenobiotic compounds. These rhizosphere bacteria have emerged as important biological components of agriculture soils with the ability to stimulate plant growth and for protection of plants from pathogens. In this study, siderophore-producing Pseudomonas species were isolated from the rhizosphere soil and were evaluated for plant growth promotion and disease control in green gram.
Screening of Pseudomonas isolates for siderophore production: All the Pseudomonas strains were screened for siderophore production using universal chromo-azurol assay. The presence of iron chelator siderophore was indicated by decolourization of blue coloured ferric CAS complex in an orange halo zone around the colonies on CAS agar plates. Out of 80 Pseudomonas strains screened for siderophore production, 37 strains were found to produce siderophore on MM9 medium (Table 1). The diameter of halo zone varied with different Pseudomonas strains. The Pseudomonas strains MPS52, MPS54, MPS90, CPS59, PS31, PS39, CP20, CP28, CP56, MP14, MP20, MP33 and MP38 showed large halo zone size measuring more than 10 mm on MM9 medium plates. Siderophore production was also determined on iron deficient succinate medium. Only two Pseudomonas strains MPS52 and CP20 showed large halo zone formation (above 10 mm). Rest of the strains showed variation in diameter of halo zone formed.
| Table 1: | Screening of selected Pseudomonas strains for siderophore production using universal chromazurol assay method |
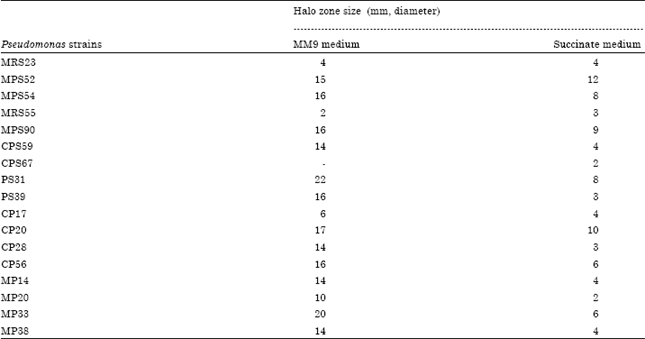 | |
| Different Pseudomonas strains were tested for siderophore production by spotting 10 μL growth suspension on MM9 or iron deficient succinate medium plates (pH 6.8). Observations were recorded after 3 days of growth at 28±2°C | |
It was found that siderophore production was more on MM9 medium as compared to succinate medium. Only 46.25% of Pseudomonas strains showed siderophore production on the two types of media tested in these studies.
The relative siderophore production ability of different Pseudomonas strains was determined by comparing the bacterial colony diameter to halo zone size on MM9 medium plates. It was measured by dividing the halo zone area (0.5H)2 to colony growth area (0.5G)2 Out of 37 Pseudomonas strains, 20 strains showed more than 3.0 ratio of halo zone to colony growth (Table 2). Pseudomonas strain MPS54 showed maximum halo zone to growth ratio (10.24) and minimum halo zone to growth ratio (less than 2.0) was obtained in Pseudomonas strains, namely MRS23, MRS55, MPS90, CPS59, CPS67, CP17, CP28 and MP14.
Interactions of siderophore-producing Pseudomonas strains with phytopathogenic fungi: Selected siderophore-producing Pseudomonas strains were screened for antagonistic activity against three fungi namely, Fusarium oxysporum, Rhizoctonia solani and Pythium aphanidermatum on PDA and NA medium plates. The growth inhibition zones in different fungi varied with different Pseudomonas strains tested (Table 3). The Pseudomonas strains MRS23, MPS54, MRS55, CP17 and CP56 showed maximum antagonistic activity against F. oxysporum on PDA and NA medium plates and inhibition zone diameter was more than 4 mm. Two strains MPS52 and CP20 did not inhibit F. oxysporum growth but these strains were found to inhibit other two fungi R. solani or P. aphanidermatum. On the other hand, Pseudomonas strains MRS23, MPS54, MRS55, CP17 and CP56 showed large growth inhibition zones (more than 4 mm) against R. solani on PDA and NA medium plates. When different Pseudomonas strains were tested for antagonistic activity against P. aphanidermatum, MPS52, MPS54, PS31, CP20 and MP38 showed maximum antagonistic activity against this fungus on PDA medium plates (Table 3).
| Table 2: | Relative siderophore production by different strains of Pseudomonas sp. on MM9 medium plates |
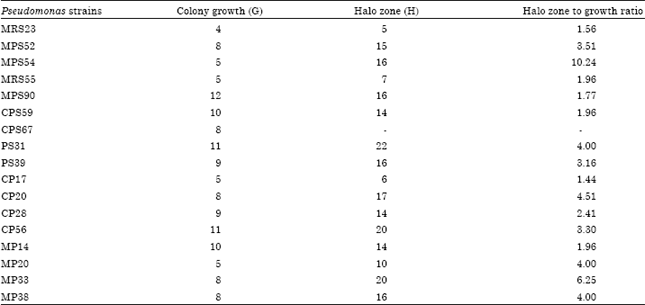 | |
| G: Growth expressed as colony diameter (mm), H: Siderophore production expressed as halo zone diameter (mm), H/G: Halo zone area (0.5 H)2 divided by growth area (0.5 G)2 | |
| Table 3: | Antagonistic activity of Pseudomonas strains on growth inhibition of phytopathogenic fungi on PDA and NA medium plates |
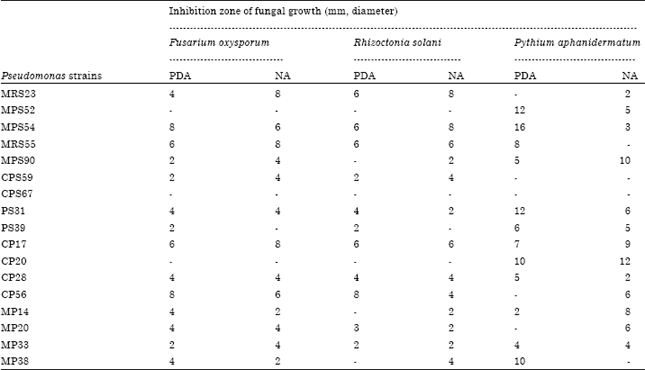 | |
| Antagonistic activity of the selected siderophore-producing strains was determined on potato dextrose agar (PDA) and nutrient agar (NA) medium by spot test method | |
Two strains MPS90 and CP20 showed large growth inhibition zone (more than 10 mm) against P. aphanidermatum on NA medium plates.
Pseudomonas strain CPS67 did not inhibit the growth of any of the fungi. It was observed that Pseudomonas strains that showed large inhibition zones against P. aphanidermatum were comparatively less inhibitory to F. oxysporum and R. solani on the PDA as well as on NA medium plates. In majority of strains, the diameter of inhibition zone was larger on NA medium than to PDA medium plates. Usually, the Pseudomonas strains which showed more siderophore production (large H:G ratio) (Table 3) showed more growth inhibition of the phytopathogenic fungi. For example, Pseudomonas strains having large H:G ratio such as MPS54, PS31, PS39, CP56 and MP33 showed sufficient growth inhibition of different phytopathogenic fungi.
Effect of Pseudomonas, Bradyrhizobium and R. solani inoculation on symbiotic parameters and disease control: Green gram plants were inoculated either singly with Bradyrhizobium sp. strain SMR15 or siderophore-producing Pseudomonas strains MP20, MPS54, CP56 or siderophore-lacking strain CPS67 and with root rot causing fungi Rhizoctonia solani. Coinoculation of Bradyrhizobium strain, Pseudomonas strains or R. solani was also done and inoculated plants were harvested after 30 and 60 days of plant growth under pot house conditions. At 30 days of plant growth, the coinoculation of Bradyrhizobium strain with Pseudomonas strains MP20, MPS54 or CP56 enhanced the nodule number, nodule weight and plant dry weights as compared to single inoculated or control plants (Table 4). The inoculation with R. solani caused root rot disease in 83.7% of inoculated plants. The coinoculation of Pseudomonas strains with R. solani lowered the disease incidence and 16.3% disease reduction was observed with Pseudomonas strains MP20 and CPS67 whereas 66.7% disease reduction was observed with MPS54.
| Table 4: | Coinoculation of selected Pseudomonas strains with Bradyrhizobium strain for nodulation and control of root rot disease of green gram at 30 days of plant growth |
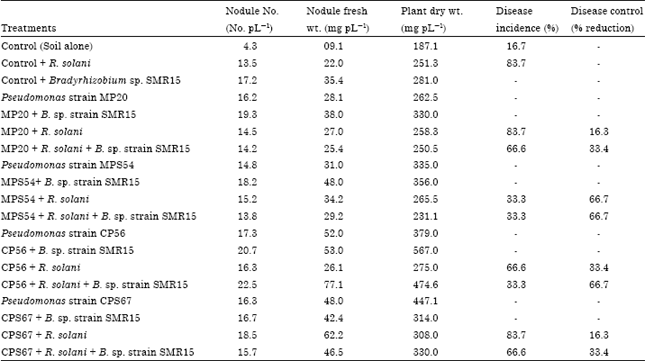 | |
| The figures are average values of 6 plants. Disease incidence is the % of plants infected and disease control is the % reduction of diseased plants after inoculation with bacteria. The values of nodule number, nodule fresh weight and plant dry weight are calculated on per plant basis | |
| Table 5: | Coinoculation of selected Pseudomonas strains with Bradyrhizobium strain for nodulation and control of root rot disease of green gram at 60 days of plant growth |
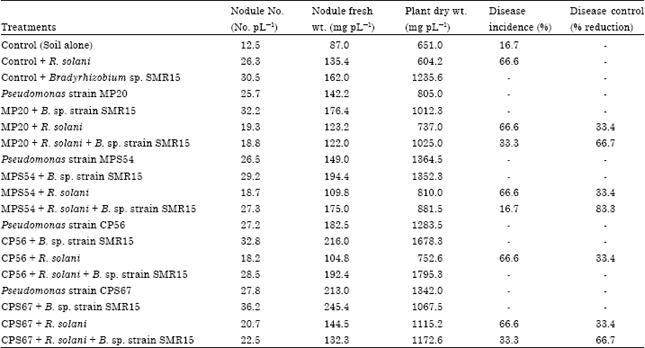 | |
| The figures are average values of 6 plants. Disease incidence is the % of plants infected and disease control is the % reduction of diseased plants after inoculation with bacteria. The values of nodule number, nodule fresh weight and plant dry weight are calculated on per plant basis | |
Maximum plant biomass enhancement (3.03 times) was found on coinoculation of Pseudomonas strain CP56 with Bradyrhizobium strains SMR15 as compared to uninoculated control plants and 2.01 times increase was observed in comparison to Bradyrhizobium inoculated plants.
At 60 days of plant growth, single inoculation with Bradyrhizobium strain SMR15 or with either Pseudomonas strains MPS54, CP56 or CPS67 or coinoculation treatments of Pseudomonas with Bradyrhizobium resulted in significant increase in nodule number, nodule fresh weight and plant dry weight as compared to control uninoculated plants (Table 5). The coinoculation of Pseudomonas strain CP56, Bradyrhizobium strain and R. solani showed maximum 275.8% increase in plant dry weight in comparison to control uninoculated plants. Similarly, the coinoculation of Pseudomonas strain CPS67 with Bradyrhizobium strain and R. solani caused 180.1% increase in plant dry weight as compared to uninoculated control plants. Maximum increase in nodule number and nodule fresh weight was observed on coinoculation of Bradyrhizobium strain with Pseudomonas strains CPS67 or CP56. Single inoculation of different Pseudomonas cultures with R. solani caused only 33.4% disease control whereas coinoculation of Bradyrhizobium and Pseudomonas strains MP20 or CPS67 caused 66.7% disease control. There were no disease symptoms on green gram plants when coinoculated with Bradyrhizobium strain and Pseudomonas strain CP56, indicating that this bacterial strain completely controlled the root rot disease under pot house conditions.
DISCUSSION
The plant rhizosphere is an important ecological environment in the soil for plant-microbe interactions (Weyens et al., 2009; Tahat et al., 2010). The early root colonizing microorganisms, in and around the growing roots, may interact with each other and with the plant in a way that may result in symbiotic, associative, neutralistic or detrimental effects (Vikram and Hamzehzarghani, 2008; Saber et al., 2009; Osman et al., 2011), depending upon the type of microorganisms involved, nutrient status in soil, abiotic and biotic soil environment and the plant defense system (Benizri et al., 2001). Fluorescent pseudomonads in the plant rhizosphere have been found to improve the plant growth through multitudinous factors viz. production of plant growth promoting substances (Ahmad et al., 2008; Sindhu et al., 2010), early colonization of root surfaces (Beniziri et al., 2001), secretions of vitamins (Derylo and Skorupsca, 1993) and through suppresion of plant diseases by production of antibiotics, siderophores, hydrolytic enzymes and HCN (Stockwell and Stack, 2007; Sindhu et al., 2009; Ahemad and Khan, 2011).
Both fluorescent and nonfluorescent Pseudomonas isolates were obtained in this study from the chickpea and green gram rhizosphere soil samples. Fifty-eight of the selected 65 Gram negative bacterial isolates showed oxidative metabolism, were Kovac’s oxidase positive and belonged to Pseudomonas. Similarly, Gupta et al. (1998) isolated 121 rhizobacteria from the rhizotic zones of green gram using 7 selective and 4 non-selective media. Gram negative bacteria accounted for 65% and the dominant genera were Pseudomonas, Bacillus, Enterobacter, Proteus and Klebsiella. Similarly, Pseudomonas was reported as the most predominant genera (42%) followed by Bacillus (28%) and Enterobacter (21%) from a total of 105 bacteria isolated from rhizosphere and rhizoplane of groundnut (Baig et al., 2002).
Pseudomonas species have been found to produce different types of siderophores such as pyoverdines, pseudobactins, colourless nocardamine, pyochelin, salicyclic acid and cepabactin (Meyer and Abdallah, 1980; Verma and Chincholkar, 2007). In this study, out of 80 Pseudomonas strains screened for siderophore production, only 46.25% strains produced siderophore on MM9 medium and succinate medium (Table 1). Ksiezniak and Kobus (1993) tested 1653 microbial species including bacteria, actinomycetes and fungi for siderophore production and highest siderophore producing bacteria belonged to genera Pseudomonas and Erwinia. Similarly, siderophore production was observed in 74.2% of Rhizobium/Bradyrhizobium strains infecting pigeon pea, out of 31 strains tested (Duhan et al., 1998).
The diameter of halo zone varied with different Pseudomonas strains. Thirteen Pseudomonas strains MPS52, MPS54, MPS90, CPS59, PS31, PS39, CP20, CP28, CP56, MP14, MP20, MP33 and MP38 showed large halo zone (more than 10 mm) on MM9 medium. On iron-deficient succinate medium, only two Pseudomonas strains MPS52 and CP20 showed large halo zone formation (above 10 mm) (Table 1). Thus, it was found that siderophore production was more on MM9 medium as compared to succinate medium. On the contrary, maximum siderophore production (216.23 μg mL-1) was reported in deferrated standard succinate medium (SSM) by Pseudomonas strain PRS9 after 72 h of incubation (Sharma and Johri, 2003a). Succinate was found better carbon source than citrate (SCM) media for siderophore production; however, defferation of media resulted in increased siderophore production in all the strains. Djibaoui and Bensoltane (2005) also reported that the ability of Pseudomonas to grow and produce siderophores is dependent on the iron content and the type of carbon sources in the medium. Highest siderophores concentration was obtained in succinate medium with Pseudomonas fluorescens. Alexander and Zuberer (1991) found that several Pseudomonas isolates obtained from grass species did not show siderophore on CAS agar but produced siderophore in liquid culture. Sharma and Johri (2003b) also reported low levels of siderophore production by Pseudomonas strains in complex media like King’s B medium, trypticase soya medium and nutrient agar medium.
Growth inhibition zones of the three fungi namely, Fusarium oxysporum, Rhizoctonia solani and Pythium aphanidermatum varied with different Pseudomonas strains tested on PDA and NA medium plates. The Pseudomonas strains MPS54, MRS55, CP17 and CP56 showed maximum antagonistic activity against F. oxysporum and inhibition zone diameter was more than 6 mm (Table 3). Pseudomonas strains MRS23, MPS54, MRS55, CP17 and CP56 showed large growth inhibition zones (more than 4 mm) against R. solani on PDA and NA medium plates. Strains MPS52, MPS54, PS31, CP20 and MP38 showed maximum antagonistic activity against P. aphanidermatum on PDA medium plates. Pseudomonas strains that showed large inhibition zones against P. aphanidermatum were comparatively less inhibitory to F. oxysporum and R. solani. The diameter of inhibition zone was larger on NA medium than to PDA medium plates in majority of the strains. Similar effects of medium composition on inhibition of fungal growth were observed by Goel et al. (2000). They reported that Pseudomonas strain MRS16 inhibited growth of different pathogenic fungi (Aspergillus sp., Fusarium oxysporum, Pythium aphanidermatum and Rhizoctonia solani) in vitro. Larger inhibition zones were obtained on nutrient agar and King’s B media compared to potato dextrose agar and pigment production media.
Becker and Cook (1988) observed zone of inhibition against Pythium ultimum var. sporangiferrum by pseudomonads isolated from roots of wheat. Siderophore-negative mutants of strain B324 were not inhibitory to Pythium indicating that fungal growth inhibition was due to production of siderophores by this strain. Inhibition of germination of chlamydospores of Fusarium oxysporum in soil was also correlated with production of siderophores by fluorescent pseudomanads (Simeon et al., 1987). The inhibition was totally or partially reversed by adding Fe (III) to the system. Yeole et al. (2001) found that twelve fluorescent pseudomonad isolates obtained from chilli, cotton, groundnut and soybean inhibited the growth of 12 test soil borne plant pathogenic fungi in Fe-deficient King’s B medium. The inhibition ranged from 3.3 to 15% and all Pseudomonas isolates produced siderophores under iron-deficient conditions. The inhibition was curtailed by 24-60% in the presence of iron (50 μm) when siderophore production was abolished. Screening of 563 bacteria obtained from the roots of pea, lentil and chickpea showed that 76% isolates produced siderophores, 5% isolates showed ACC deaminase activity and 7% isolates were capable of indole production (Hynes et al., 2008). Twenty-six isolates (5%) suppressed the growth of Pythium species strain p88-p3, 7% suppressed the growth of Fusarium avenaceum and 9% suppressed the growth of R. solani CKP7. Ahmad et al. (2008) reported that siderophore production and antifungal activity was exhibited by 10 to 12.77% of Azotobacter and Pseudomonas isolates. Pseudomonas Ps5 and Bacillus B1 isolates showed broad-spectrum antifungal activity on Muller-Hinton medium against Aspergillus, Fusarium and Rhizoctonia bataticola.
Different microbial antagonists have been found to control the root rot disease caused by Rhizoctonia solani in different crops (Abeysinghe, 2009; Dar et al., 2011; Osman et al., 2011). In this study, inoculation of green gram with R. solani caused root rot disease in 83.7% of inoculated plants (Table 4) and the coinoculation of Pseudomonas strains with R. solani lowered the disease incidence. Disease reduction varied from 16.3% to 66.7% in coinoculated plants. At 60 days of plant growth, inoculation of different Pseudomonas cultures with R. solani caused only 33.4% disease control whereas coinoculation of Bradyrhizobium and Pseudomonas strains caused 66.7% (with MP20 and CPS67 strains) to 83.3% (with MPS54 strain) disease control. On coinoculation of Bradyrhizobium strain with Pseudomonas strain CP56, no disease symptoms were observed on green gram plants indicating that this bacterial strain completely controlled the root rot disease under pot house conditions. Similar effects of siderophore-producing bacteria on disease suppression were reported on wilt disease caused by F. oxysporum (Duijff et al., 1994) and Pythium-induced post emergence damping off disease in tomato (Buysens et al., 1996). The observed antagonism and protection of plants was attributed to the siderophore-mediated iron competition. Kloepper et al. (1980) showed that inoculation with Pseudomonas strain B10 as a cell suspension and pure pseudobactin at 10 μm concentrations caused significant increases in plant growth compared with water treated controls. Boruah and Kumar (2002) reported that seed bacterization with Pseudomonas strains that displayed in vitro antibiosis against many pathogenic fungi, improved germination, shoot height, root length, fresh and dry mass, enhanced yield and chlorophyll content of leaves in the five test crop plants under field conditions. They reported that the plant growth promotion was due to siderophore production where disease suppression was due to the antibiotic substance. Similarly, Sharma and Johri (2003a) found that strains GRP3A and PRS9 which showed antagonistic activity against Colletotrichum dematium, Rhizoctonia solani and Sclerotium rolfsii showed significant increase in germination percentage and plant growth of maize seeds.
During these studies, inoculation with Bradyrhizobium strain SMR15 or with Pseudomonas strains MPS54, CP56 or CPS67 or coinoculation of Pseudomonas with Bradyrhizobium resulted in significant increase in nodule number, nodule fresh weight and plant dry weight as compared to control uninoculated plants at 30 and 60 days of plant growth (Table 4, 5). The coinoculation of fungi with Pseudomonas strain CP56 and Bradyrhizobium strain showed maximum enhancement in nodule number (1.93 times) and plant dry weight (4.4 times) in comparison to uninoculated control plants. The coinoculation of Pseudomonas strain CP56 with Bradyrhizobium strain and R. solani showed maximum 275.8% increase in plant dry weight whereas coinoculation of Pseudomonas strain CPS67 caused 180.1% increase in plant dry weight in comparison to control uninoculated plants. Similar enhancement of nodulation and nitrogen fixation after coinoculation of Bradyrhizobium and Pseudomonas were also observed in other legumes. For example, Bolton et al. (1990) studied coinoculation effect of R. leguminosarum and a toxin-producing Pseudomonas sp. in pea roots under Leonard jar conditions and found that Pseudomonas sp. increased the number of nodules formed by R. leguminosarum on pea roots. Mahmoud and Abd-Allah (2001) reported that siderophore producing Penicillium chrysogenum and P. aeruginosa significantly enhanced nodulation and nitrogen fixation of mungbean compared with plants infected with Bradyrhizobium strain alone. The enhancement of nodulation after coinoculation of Bradyrhizobium and Pseudomonas could be attributed to increased bradyrhizobial population and colonization of legume roots (Chebotar et al., 2001), elongation and stimulation of rootlets (De Bellis and Ercolani, 2001) and enhancement of flavonoids in roots after application of Pseudomonas strains which act as inducer of nodulation genes (Peter and Verma, 1990).
CONCLUSION
Siderophore-producing Pseudomonas strains isolated from the rhizosphere soil of chickpea and green gram were found to inhibit the growth of wilt-causing fungi (Fusarium oxysporum), root rot-causing fungi (Rhizoctonia solani) and damping-off disease-causing fungi (Pythium aphanidermatum) on medium plates. Seed inoculation of antagonistic Pseudomonas cultures caused 33.4 to 100 per cent reduction of root rot disease in green gram under pot house conditions. Coinoculation of Pseudomonas strains with Bradyrhizobium strain SMR15 in green gram enhanced the nodule number, nodule fresh weight and plant dry weight as compared to Bradyrhizobium-inoculated or uninoculated control plants. The results suggested that microorganisms from the soil could replace the use of pesticides and nitrogenous fertilizers to reduce the pollution of soil and water.
REFERENCES
- Abeysinghe, S., 2009. Effect of combined use of Bacillus subtilis CA32 and Trichoderma harzianum RU01 on biological control of Rhizoctonia solani on Solanum melongena and Capsicum annuum. Plant Pathol. J., 8: 9-16.
CrossRefDirect Link - Ahemad, M. and M.S. Khan, 2011. Assessment of plant growth promoting activities of rhizobacterium Pseudomonas putida under insecticide-stress. Microbiol. J., 1: 54-64.
CrossRefDirect Link - Ahmad, F., I. Ahmad and M.S. Khan, 2008. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res., 168: 173-181.
CrossRefDirect Link - Becker, J.O. and R.J. Cook, 1988. Role of siderophores in suppression of Pythium species and production of increased-growth response of wheat by fluorescent pseudomonads. Phytopathology, 78: 778-782.
CrossRefDirect Link - Benizri, E., E. Baudoin and A. Guckert, 2001. Root colonization by inoculated plant growth-promoting rhizobacteria. Biocontrol. Sci. Technol., 11: 557-574.
CrossRefDirect Link - Boruah, H.P.D. and B.S. Kumar, 2002. Plant disease suppression and growth promotion by a fluorescent Pseudomonas strain. Folia Microbiol., 47: 137-143.
PubMedDirect Link - Brisbane, P.G. and A.D. Rovira, 1988. Mechanism of inhibition of Gaeumannomyces graminis var. tritici by fluorescent pseudomonads. Plant Pathol., 37: 104-111.
CrossRefDirect Link - Buyer, J.S. and L.J. Sikora, 1990. Rhizosphere interactions and siderophore. Plant Soil, 129: 101-107.
CrossRefDirect Link - Buysens, S., K. Heungens, J. Poppe and M. Hofte, 1996. Involvement of pyochelin and pyoverdin in suppression of Pythium-induced damping-off of tomato by Pseudomonas aeruginosa 7 NSK2. Applied Environ. Microbiol., 62: 865-871.
Direct Link - Dar, G.H., M.A. Beig, F.A. Ahanger, N.A. Ganai and M.A. Ahangar, 2011. Management of root rot caused by Rhizoctonia solani and Fusarium oxysporum in blue pine (Pinus wallichiana) through use of fungal antagonists. Asian J. Plant Pathol., 5: 62-74.
CrossRefDirect Link - De Bellis, P. and G.L. Ercolani, 2001. Growth interaction during bacterial colonization of seedling rootlets. Applied Environ. Microbiol., 67: 1945-1948.
CrossRefDirect Link - Derylo, M. and A. Skorupska, 1993. Enhancement of symbiotic nitrogen fixation by vitamin-secreting fluorescent Pseudomonas. Plant Soil, 154: 211-217.
CrossRefDirect Link - Kumar, B.S., 1996. Crop improvement and disease suppression by a Bacillus sp. SR2 from peanut rhizosphere. Indian J. Exp. Biol., 34: 794-798.
PubMedDirect Link - Duhan, J.S., S.S. Dudeja and A.L. Khurana, 1998. Siderophore production in relation to N2 fixation and iron uptake in pigeonpea-Rhizobium symbiosis. Folia Microbiol., 43: 421-426.
CrossRefDirect Link - Duijff, B.J., P.A.H.M. Bakker and M. Schippers, 1994. Suppression of Fusarium wilt of carnation by Pseudomonas putida WCS358 at different levels of disease incidence and iron availability. Biocontrol Sci. Technol., 4: 270-288.
CrossRefDirect Link - Ehteshamul-Haque, S. and A. Ghaffar, 1993. Use of rhizobia in the control of root rot diseases of sunflower, okra, soybean and mungbean. J. Phytopathol., 138: 157-163.
CrossRefDirect Link - Gill, P.R. and G.J. Warren, 1988. An iron antagonized fungistatic agent that is not required for iron assimilation from a flourescent rhizosphere pseudomonad. J. Bacteriol., 17: 163-170.
Direct Link - Goel, A.K., S.S. Sindhu and K.R. Dadarwal, 2000. Pigment diverse mutants of Pseudomonas sp.: Inhibition of fungal growth and stimulation of growth of Cicer arietinum. Biol. Plant., 43: 563-569.
CrossRefDirect Link - Hynes, R.K., G.C. Leung, D.L. Hirkala and L.M. Nelson, 2008. Isolation, selection and characterization of beneficial rhizobacteria from pea, lentil and chickpea grown in Western Canada. Can. J. Microbiol., 54: 248-258.
CrossRefPubMedDirect Link - King, E.O., M.K. Ward and D.E. Raney, 1954. Two simple media for the demonstration of pyocyanin and fluorescin. Transl. Res., 44: 301-307.
PubMedDirect Link - Kloepper, J.W., J. Leong, M. Teintze and M.N. Schroth, 1980. Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature, 286: 885-886.
CrossRefDirect Link - Lemanceau, P., A. Robin, S. Mazurier and G. Vansuyt, 2007. Implication of Pyoverdines in the Interactions of Fluorescent Pseudomonads with Soil Microflora and Plants in the Rhizosphere. In: Microbial Siderophores, Verma, A. and S. Chincholkar, (Eds.). Springer-Verlag, Berlin, Heidelberg, pp: 165-192.
- Leong, J., 1986. Siderophores: Their biochemistry and possible role in the biocontrol of plant pathogens. Annu. Rev. Phytopathol., 24: 187-209.
CrossRefDirect Link - Mahmoud, A.L.E. and M.A. Abd-allah, 2001. Siderophores production by some microorganisms and their effect on Bradyrhizobium-mungbean symbiosis. Int. J. Agric. Biol., 3: 157-162.
Direct Link - Mayer, J.M. and M.A. Abdallah, 1978. The fluorescent pigment of Pseudomonas fluorescens: Biosynthesis, purification and physicochemical properties. J. Gen. Microbiol., 107: 319-328.
Direct Link - Meyer, J.M. and M.A. Abdallah, 1980. The siderochromes of nonfluorescent pseudomonads. Production of nocardamine by Pseudomonas stutzeri. J. Gen. Microbiol., 118: 125-129.
CrossRefDirect Link - Miller, H.J., G. Henken and J.A. van Veen, 1989. Variation and composition of bacterial populations in the rhizospheres of maize, wheat and grass cultivars. Can. J. Microbiol., 35: 656-660.
CrossRefDirect Link - Neilands, J.B., 1982. Microbial envelope proteins related to iron. Annu. Rev. Microbiol., 36: 285-309.
CrossRefPubMedDirect Link - Osman, M.E.H., M.M. El-Sheekh, M.A. Metwally, A.E.A. Ismail and M.M. Ismail, 2011. Antagonistic activity of some fungi and cyanobacteria species against Rhizoctonia solani. Int. J. Plant Pathol., 2: 101-114.
CrossRefDirect Link - Peter, N.K. and D.P.S. Verma, 1990. Phenolic compounds as regulators of gene expression in plant-microbe interaction. Mol. Plant-Microbe Interact., 3: 4-8.
PubMed - Raaijmakers, J.M., M. Leeman, M.M.P. van Oorschot, I.V. Sluis, B. Schippers and P.A.H.M. Bakker, 1995. Dose-response relationships in biological control of Fusarium wilt of radish by Pseudomonas spp. Phytopathology, 85: 1075-1081.
CrossRef - Rachid, D. and B. Ahmed, 2005. Effect of iron and growth inhibitors on siderophores production by Pseudomonas fluorescens. Afr. J. Biotechnol., 4: 697-702.
Direct Link - Saber, W.I.A., K.M. Abd El-Hai and K.M. Ghoneem, 2009. Synergistic effect of Trichoderma and Rhizobium on both biocontrol of chocolate spot disease and induction of nodulation, physiological activities and productivity of Vicia faba. Res. J. Microbiol., 4: 286-300.
CrossRefDirect Link - Scherwinski, K., R. Grosch and G. Berg, 2008. Effect of bacterial antagonists on lettuce: Active biocontrol of Rhizoctonia solani and negligible, short-term effects on nontarget microorganisms. FEMS Microiol. Ecol., 64: 106-116.
CrossRef - Schwyn, B. and J.B. Neilands, 1987. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem., 160: 47-56.
CrossRefPubMedDirect Link - Sharma, A. and B.N. Johri, 2003. Combat of iron-deprivation through a plant growth promoting fluorescent Pseudomonas strain GRP3A in mung bean (Vigna radiata L. Wilzeck). Microbiol. Res., 158: 77-81.
CrossRef - Sharma, A. and B.N. Johri, 2003. Growth promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9 in maize (Zea mays L.) under iron limiting conditions. Microbiol. Res., 158: 243-248.
CrossRefPubMedDirect Link - Siddiqui, I.A., S. Ehetshamul-Haque and S.S. Shaukat, 2001. Use of rhizobacteria in the control of root rot-root knot disease complex of mungbean. J. Phytopathol., 149: 337-346.
CrossRefDirect Link - Sindhu, S.S., S.K. Gupta and K.R. Dadarwal, 1999. Antagonistic effect of Pseudomonas spp. on pathogenic fungi and enhancement of growth of green gram (Vigna radiata). Biol. Fertil. Soils, 29: 62-68.
CrossRefDirect Link - Sindhu, S.S., S. Suneja, A.K. Goel, N. Parmar and K.R. Dadarwal, 2002. Plant growth promoting effects of Pseudomonas sp. on coinoculation with Mesorhizobium sp. Cicer strain under sterile and wilt sick soil conditions. Applied Soil Ecol., 19: 57-64.
CrossRefDirect Link - Sloger, C., 1969. Symbiotic effectiveness and N2 fixation in nodulated soybean. Plant Physiol., 44: 1666-1668.
Direct Link - Stockwell, V.O. and J.P. Stack, 2007. Using Pseudomonas spp. for integrated biological control. Phytopathology, 97: 244-249.
CrossRefDirect Link - Tahat, M.M., K. Sijam and R. Othman, 2010. Mycorrhizal fungi as a biocontrol agent. Plant Pathol. J., 9: 198-207.
CrossRefDirect Link - Validov, S., F. Kamilova, S. Qi, D. Stephan, J.J. Wang, N. Makarova and B. Lugtenberg, 2007. Selection of bacteria able to control Fusarium oxysporum f.sp. radicis-lycopersici in stonewool substrate. J. Applied Microbiol., 102: 461-471.
PubMed - Van Rossum, D., A. Muyotcha, H.W. van Verseveld, A.H. Stouthamer and F.C. Boogred, 1994. Siderophore production by Bradyrhizobium sp. strains nodulating groundnut. Plant Soil, 163: 177-187.
CrossRefDirect Link - Vikram, A. and H. Hamzehzarghani, 2008. Effect of phosphate solubilizing bacteria on nodulation and growth parameters of greengram (Vigna radiata L. Wilczek). Res. J. Microbiol., 3: 62-72.
CrossRefDirect Link - Weller, D.M., 2007. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology, 97: 250-256.
CrossRef - Weyens, N., D. van der Lelie, S. Taghavi, L. Newman and J. Vangronsveld, 2009. Exploiting plant-microbe partnerships to improve biomass production and remediation. Trends Biotechnol., 27: 591-598.
CrossRefPubMedDirect Link - Yeole, R.D., B.P. Dave and H.C. Dube, 2001. Siderophore production by fluorescent pseudomonads colonizing roots of certain crop plants. Indian J. Exp. Biol., 39: 464-464.
PubMed - Woyessa, D. and F. Assefa, 2011. Effect of plant growth promoting rhizobacteria on growth and yield of Tef (Eragrostis tef Zucc. Trotter) under greenhouse condition. Res. J. Microbiol., 6: 343-355.
CrossRef - Alexander, D.B. and D.A. Zuberer, 1991. Use of chrome azurol S reagents to evaluate siderophore production by Rhizosphere bacteria. Biol. Fertil. Soils, 12: 39-45.
CrossRefDirect Link - Chebotar, V.K., C.A. Asis and S. Akao, 2001. Production of growth-promoting substances and high colonization ability of rhizobacteria enhance the nitrogen fixation of soybean when coinoculated with Bradyrhizobium japonicum. Biol. Fertil. Soils, 34: 427-432.
CrossRef
zawthulwin Reply
I want to read for blackgram cultivation guide