
Mohamed El-anwar H. Osman
Department of Botany, Faculty of Sciences, Tanta University, Tanta 31527, Egypt
Mostafa M. El-Sheekh
Department of Botany, Faculty of Sciences, Tanta University, Tanta 31527, Egypt
Metwally A. Metwally
Department of Botany, Faculty of Sciences, Tanta University, Tanta 31527, Egypt
Abd El-whab A. Ismail
Plant Pathology Institute, Agriculture Research Center, Giza, Egypt
Mona M. Ismail
Department of Botany, Faculty of Sciences, Tanta University, Tanta 31527, Egypt
International Journal of Plant Pathology
Year: 2011 | Volume: 2 | Issue: 3 | Page No.: 101-114







ABSTRACT
This study was conducted to investigate the suppression effect of some antagonistic fungi and cyanobacterial species against Rhizoctonia solani as the causal agent of soybean root rot. Growth of Rhizoctonia solani as the causal agent of root rot of soybeans was inhibited (in vitro and greenhouse conditions) in the presence of some antagonistic fungi (Gliocladium deliquescens, G. virens, Trichoderma hamatum and T. harzianum) and cyanobacterial species (Nostoc entophytum and N. muscurum). The results show that Trichoderma harzianum was the best antagonistic fungi whereas Nostoc entophytum as cyanobacteria showed antifungal activity higher than Nostoc muscurum, the inhibitory effect was dependant on the type of the bioagent. In experiments carried out in greenhouse, the growth parameters (length, weight, carbohydrate, protein and nitrogen) of the infected soybean plants showed different responses to the tested biological agents as compared to untreated infected plant. It could be concluded from the obtained data the fruitful use of the tested biotic factors for controlling rot root of soybean induced by Rhizoctonia solani.
PDF Abstract XML References Citation
Received: April 07, 2011;
Accepted: May 19, 2011;
Published: July 16, 2011
How to cite this article
Mohamed El-anwar H. Osman, Mostafa M. El-Sheekh, Metwally A. Metwally, Abd El-whab A. Ismail and Mona M. Ismail, 2011. Antagonistic Activity of Some Fungi and Cyanobacteria Species against Rhizoctonia solani. International Journal of Plant Pathology, 2: 101-114.
DOI: 10.3923/ijpp.2011.101.114
URL: https://scialert.net/abstract/?doi=ijpp.2011.101.114
DOI: 10.3923/ijpp.2011.101.114
URL: https://scialert.net/abstract/?doi=ijpp.2011.101.114
INTRODUCTION
Plant diseases play direct role in the destruction of natural resources in agriculture. In particular, soil borne pathogens cause important losses, fungi being the most aggressive. The distribution of several phytopathogenic fungi, such as Pythium, Phytophthora, Botrytis, Rhizoctonia and Fusarium, has spread during the last few years due to changes introduced in farming with detrimental effects on crops of economic importance. In addition, not only growing crops but also stored fruits are prey to fungal infections (Chet et al., 1997). Rhizoctonia root rot and hypocotyl rot caused by Rhizoctonia solani, is a common disease of soybean which is the most important commercial crops playing key role in economical and social affairs in Egypt and also an important nitrogen-fixing leguminous crop cultivated for food and feed (Bradley et al., 2002).
Rhizoctonia solani is common soil-inhabiting fungus with a wide host range that includes field crops, vegetables, fruits and ornamentals (Bohlooli et al., 2005). Rhizoctonia Foliar Blight (RFB) of soybean occurs in many tropical and subtropical regions, causing yield reductions of up to 70% and in Brazil, up to 60% (Meyera et al., 2006). Fusarium oxysporum, Rhizoctonia solani, Macrophomina phaseolina and Sclerotium rolfsii are common fungal pathogens to soybean causing damping off, root rot and wilt diseases resulting in serious economic losses (Fayzalla et al., 2009).
Most soil-borne pathogens are difficult to control by conventional control measures such as the use of resistant cultivars and synthetic fungicides (Weller et al., 2002). Rhizoctonia diseases are difficult because this pathogen survives for many years as sclerotia in soil or as mycelium in organic matter under numerous environmental conditions and has an extremely wide host range (Grosch et al., 2003). Moreover, the use of fungicides, besides being expensive and involving risks to the environment associated with the application of chemicals, is not totally affective and may lead to the appearance of new, resistant strains of pathogens (Soylu et al., 2005). As a consequence, there is an increased emphasis on ways to minimize the use of fungicides.
Interest in biological control has increased recently, fuelled by public concerns over the use of chemicals in the environment (Whipps, 2001). Biological control agents for plant diseases are currently being examined as alternatives to synthetic pesticides due to their perceived increased level of safety and minimal environmental impacts (Brimmer and Boland, 2003) and which reduce the disease and are perceived as less harmful than conventional fungicides (Washington et al., 1999). It has long been recognized that the biological control became recently an effective strategy for fighting plant pathogens (Kabeil et al., 2008).
Microorganisms that can grow in the rhizosphere are ideal for use as biocontrol agents against soil-borne pathogens, since the rhizosphere provides the front line of defense for roots against attack by pathogens (Lozovaya et al., 2004). Several antagonistic bacteria and fungi to soil borne pathogens were reported as biocontrol agents of many pathogens induced root rot and wilt diseases (Haggag, 1998). Fungal biological control agents have several mechanisms of action that allow them to control pathogens, including mycoparasitism, production of antibiotics or enzymes, competition for nutrients and the induction of plant host defenses (Brimmer and Boland, 2003). A broad spectrum of fungal antagonists was evaluated as potential Biocontrol Agents (BCAs) against the soil-borne pathogen R. solani (Grosch et al., 2006). Antagonists of phytopathogenic fungi have been used to control plant diseases and 90% of such applications have been carried out with different strains of the fungus Trichoderma (Hermosa et al., 2000). Furthermore, Trichoderma strains are effective in controlling plant diseases and the action of fungal hydrolytic enzymes is considered as the main mechanism involved in the antagonistic process (Szekeres et al., 2004). Gachomo and Kotchoni (2008) revealed the production of volatiles by Trichoderma species against the pathogenic microorganisms. T. harzianum is a well known biological controlling agent against several soil borne phytopathogens (Yadav et al., 2011).
Algae are one of the chief biological agents that have been studied for the control of plant pathogens (Hewedy et al., 2000). Cyanobacteria were found to be a rich source for various products of commercial, pharmaceutical or toxicological interest: primary metabolites, such as proteins, fatty acids, vitamins or pigments (Borowitzka, 1995).
Various strains of cyanobacteria are known to produce intracellular and extracellular metabolites with diverse biological activities such as antialgal, antibacterial, antifungal and antiviral activity (Noaman et al., 2004). They have received little attention as potential biocontrol agents of plant diseases. Kulik (1995) stated that for a number of reasons, cyanobacteria and algae are suitable candidates for exploitation as biocontrol agents of plant pathogenic bacteria and fungi: Cyanobacteria and algae produce a large number of antibacterial and antifungal products, many can grow in quantity in mass culture and they are not a threat to the environment (except for the production of toxic blooms in freshwater and marine habitats and slimy areas on turf by a relatively small number of cyanobacteria).
The aim of the present study is focused on detection the ability of some antagonistic fungi and some cyanobacteria in suppressing root rot diseases of Glycine max L. caused by R. solani in vitro and in vivo and determine their effects on some growth parameters of Glycine max L.
MATERIALS AND METHODS
Biological agents
Fungi: The antagonistic fungi (Gliocladium deliquescens, G. virens, Trichoderma hamatum and T. harzianum) and Pathogenic fungus (Rhizoctonia solani) were procured from Plant Pathology Department, El-Gemmeiza Agricultural Research Station, (El-Gharbia Governorate, Egypt) and incubated on Potato Dextrose Agar (PDA) slants and plates at 28±1°C to establish growth then stored at 5°C in refrigerator.
Blue green algae (Cyanobacteria): Nostoc entophytum and N. muscurum were obtained from Botany Department, Faculty of Science, Tanta University. The identified cyanobacteria inoculated on BG11 (Rippka et al., 1979) nutrient agar slants and left in a diffused light at room temperature (28±2°C) to grow for 12 days thereafter, they were kept in a refrigerator at 4°C.
Plant: Soybean (Glycine max (L.) Merrill) seeds, cultivar (Giza 111) were kindly supplied by the Legumes Department, El-Gemmeiza Agricultural Research Station, Agricultural Research Center, Egypt. All experiments were carried out in Mycology laboratory and greenhouses of Botany Department, Faculty of Science, Tanta University, Tanta, Egypt.
Preparation of antifungal extracts: Cyanobacteria mass from an axenic culture growing in BG11 were separated from the culture medium by centrifugation after 12 days of incubation at 30°C under continuous illumination (30 μE/m2/S). The pellets were dried at 60°C for 24 h (Khan et al., 1988; Vlachos et al., 1996) and their extract (acetone, chloroform, methanol and water) prepared according to the methods of Katircioglu et al. (2006).
Antifungal assay by the agar disc diffusion method: Petri dishes (9 cm in diameter) contains 15 mL of PDA medium were divided into two equal halves, the first half was inoculated with a disk (0.5 cm in diameter) of Rhizoctonia solani and the second half was inoculated with a disk (0.5 cm in diameter) of cyanobacteria extracts or a disk of the tested antagonistic fungi (Bauer et al., 1966; Nair et al., 2005). The percentage of inhibition (I%) was calculated after 4 days of incubation at 28±1°C according to Topps and Wain (1957) equation.
Determination of the total phenolic contents of cyanobacteria: The total phenolic contents of cyanobacteria were determined as described by Jindal and Singh (1975).
Determination of the polysaccharides of cyanobacteria: Polysaccharides of tested cyanobacteria (Intracellular (IPS) and Extracellular Polysaccharides (EPS)) were extracted and determined as the method described by Shi et al. (2007).
Biological control experiment (Preparation of pathogen and biological control inoculation under greenhouse conditions): Pathogencity test is primary test for determination of the suitable concentration of R. solani which the casual agent of rot root under greenhouse conditions in early May 2009. The inoculum was prepared by dispensing 100 g of mixture wheat bran and sand (2:1) in bottles, then moistured with water. Contents of bottles were autoclaved for 20 min at 1.5 atm., then inoculated with R. solani which had been grown on PDA for one week and incubated at 28±1°C for 14 days. Autoclaved soil was placed in greenhouse and infested with inocula of R. solani one week before sowing at the rates of 10, 30, 50, 70 and 90 g kg-1 soil. Pre emergence damping-off was recorded using the following equation after 15 days of sowing as percentage of infected plants.
Starter cultures of both pathogen and antagonistic fungi were cultured in sterilized mixture wheat bran and sand (2:1) in bottles, then moistured with water and incubated at 28±1°C for 14 days. The bottles were shaken daily to mix and spread the fungal inoculum well on the growth substrate. During the season (late May to June 2009), sterilized soil was placed into 25 cm diameter plastic pots, each pot contained 3 Kg soil. Soil infestation was carried out one week before sowing at the rate of 30 g kg-1 R. solani inoculum and the fungal inoculum was mixed with the sterilized soil one week before sowing at the rate of 3% (w/w) while the fresh cyanobacterial inoclume was added at 0.3% (w/w) and kept moist. Twenty sterilized soybean seeds were sown in each plastic pot and replicated five times for each particular treatment. Post-emergence damping off was recorded after 45 days of sowing for each treatment as mention above.
Plant analysis: Measurement of soybeans growth included post-emergence damping-off, surviving seedlings, plant height, fresh and dry weights of shoot and roots, carbohydrate content (Nelson, 1944; Naguib, 1964), protein (Bradford, 1976), nitrogen (Naguib, 1969) and phosphorous content (Allen et al., 1974) after 45 days of sowing.
Statistical analysis: The presented results are the Means±SD (standard deviation) of at least five readings. One way Analysis of Variance (ANOVA) was done using the SAS (1996) program version 6.12. The objective of statistical analysis was to determine any significant different between treatments.
RESULTS
Antifungal activity of the tested antagonistic fungi in vitro: The results presented in Fig. 1 and show that all tested antagonistic fungi (G. deliquescens, G. virens, T. hamatum and T. harzianum) exhibited antifungal activity against Rhizoctonia solani in vitro after 4 days of incubation. The antimicrobial activities of the tested fungi could be arranged in the following sequence? T. harzianum (63%)> G. virens (55%)> T. hamatum (49.8%)> G. deliquescens (46.4%) at p = 5%.
Antifungal activity of the some extracts of the tested cyanobacteria in vitro: The antifungal activity of two cyanobacterial sp. (N. entophytum and N. muscurum) as acetone, chloroform, methanol and water extracts is represented in Fig. 2a and b. The results show that extracts exhibited antifungal activity except water extract of N. muscurum which showed no antifungal activity. However, the strongest antifungal activity was observed in water extract of N. entophytum (44.4%).
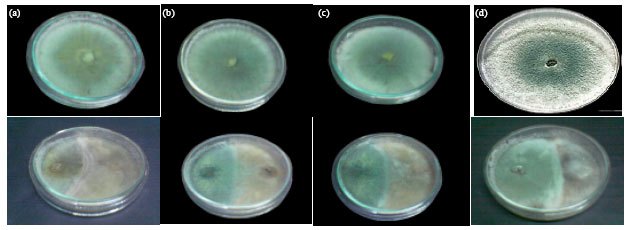 | |
| Fig. 1(a-d): | Antagonistic effect between Rhizoctonia solani and some antagonistic fungi on PDA medium (four days old). (d) T. harzianum (b) G. virens (c) T. hamatum (a) G. deliquescens |
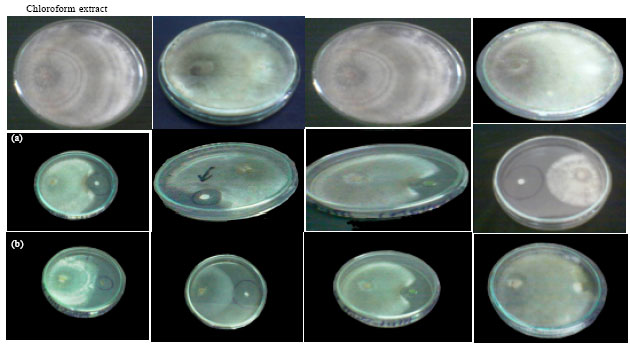 | |
| Fig. 2(a-b): | Antifungal activity of some extracts of (a) Nostoc entophytum and (b) Nostoc muscurum against Rhizoctonia solani on PDA medium (four days old) |
Role of phenol and polysaccharides content as antifungal agents: The relationship between the antifungal activity of the tested cyanobacteria and their polysaccharides and phenol contents were determined by estimating their contents in the tested cyanobacterial species. The results show that as the phenol contents of the tested cyanobacterial species were increased, their antifungal activity was increased (Table 1).
The content of polysaccharides content of N. entophytum was higher than N. muscurum and showed higher antifungal activity than N. muscurum.
| Table 1: | Total phenol contents and polysaccharides of blue green algae |
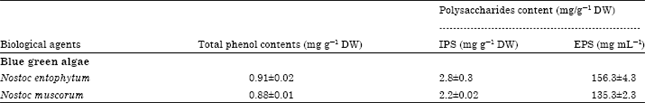 | |
| IPS: Intracellular polysaccharides EPS: Extracellular polysaccharides. Values represent Mean±SD (n = 5) | |
Antifungal activity of the tested organisms under greenhouse conditions: This trial was conducted from May to June 2009. As a general trend, R. solani caused a highly significant reduction in the measured soybean growth parameter such as survival ratio by 77.22% and caused a highly significant increase in the post-emergence damping off by 83.3% after 45 days of sowing at p<0.001(Fig. 3a).
The root depth and shoot length of infected soybeans with R. solani exhibited progressive decreases throughout the cultivation period up to 45 days by about 31 and 48.3%, respectively at p<0.001 (Fig. 3b).
The infection of soybean with inoculum of R. solani was found to cause a highly significant decrease soybean fresh and dry weights amounted by 48.3 and 51.1%, respectively below the healthy control at p<0.001 (Fig. 1b, 3c, 3d).
Under greenhouse conditions, addition of (3 g kg-1) T. harzianum induced highly significant increase in the soybean survival rates by 219.2% at p<0.001 (Fig. 3a). T. harzianum was the more effective than G. virens in increasing soybean root depth and shoot length by 53.8 and 46.4 % as compared to infected plant (Fig. 3b). Addition of 3% T. harzianum or G. virens, separately induced highly significant increase in the total fresh weight above the infected control level by 53.8 and 45.4% (Fig. 3c) and caused a highly significant increase in the total dry weight by 64.5 and 59.8%, respectively at p<0.001 (Fig. 3d).
However, treatment the infected soil with 0.3% N. entophytum or N. muscurum caused highly significant increase in the total fresh weight by 39.5 and 34.4% (Fig. 3c) and led to increase the total dry weight by 57.9 and 51.9%, respectively at p<0.001 above the control value after 45 days of sowing (Fig. 3d).
Compared to control culture, 3% (w/w) R. solani infected soybean had highly significant reduction in carbohydrate contents (DRV, TRV and sucrose) of soybean shoot system amounted by 24.8, 45.8 and 66.1%, respectively (Fig. 3e). The same treatments caused also decrease in root system DRV, TRV and sucrose by 57.8, 26.8 and 14.6% below the control value under greenhouse conditions at p<0.001(Fig. 3f). On the other hand, the carbohydrate content of soybeans shoot and root system cultivated in soil contained 0.3% (w/w) T. harzianum or G. virens had significant increase as compared with infected control after 45 days of sowing under greenhouse condition.
Protein content of the infected soybeans shoot and root showed significant increase by 75.2, 58% as compared with uninfected plant Hence nitrogen contents showed highly significant increase by 58, 42.6%, respectively after 45 days of sowing under greenhouse conditions (Fig. 3g, h). On the other hand, the protein and nitrogen contents of infected soybean treated with tested antagonistic fungi were lower than untreated infected plant but were higher than the uninfected plant (Fig 3g, h).
The algal treatments e.g., N. entophytum and N. muscurum caused significant reduction in the protein (Fig. 3g) and nitrogen content below the infected untreated soybeans after 45 days of sowing but these contents of infected soybean treated with tested algae were higher than uninfected control (Fig. 3h).
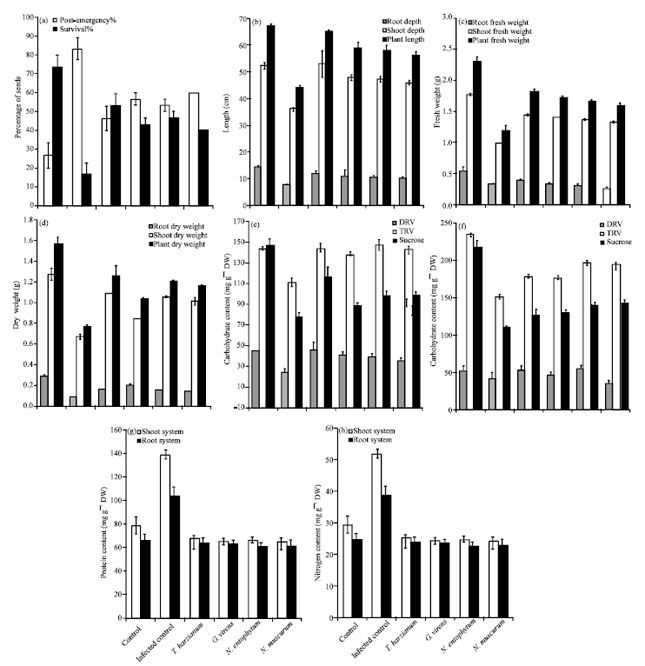 | |
| Fig. 3(a-h): | Effect of the tested bioagents on (a) post damping off and survival percentage (I%) (b) length, (c) fresh, (d) dry weight, (e, h) carbohydrate, (g) protein and (h) nitrogen contents of infected Glysine max L. with Rhizoctonia solani after 45 days of sowing under greenhouse conditions |
The results indicate that the antifungal activity of N. entophytum under greenhouses conditions were also higher than that of N. muscurum.
DISCUSSION
Antifungal activity of the tested organisms in vitro: The results show that all of the tested organisms exhibited inhibitory effect on R. solani in vitro. The inhibitory effects as measured by inhibition ratio were extremely variable as measured by the diameter of the inhibition zone according to the species of the tested organism. With respect to antagonistic fungi, T. harzianum (Fig. 1d) showed the strongest antagonistic effect followed by G. virens (Fig. 1b), T. hamatum (Fig. 1c) then G. deliquescens (Fig. 1a). These results are in accordance with the data obtained by El-Kader (1997) who found that T. harzianum (as a biocide) decreased R. solani growth which the causal organism of bean root rot disease by 69-74% in vitro. Singh and Chand (2006) recorded that T. harzianum gave maximum inhibition of the R. solani (75.55%) followed by G. virens which exhibited 57.77% mycelial growth inhibition under laboratory conditions. Kalai et al. (2008) stated that Trichoderma species are known to have strong antifungal effect partly as a result of their production of extracellular protease and chitinase enzymes which hydrolyse the main constituent of the fungal cell wall.
The obtained results showed that water extract of N. entophytum exhibited high antifungal activity while no activity was observed in water extract of N. muscurum against R. solani. On the other hand, chloroform extract showed marked antifungal activity in case of N. muscurum whereas the chloroform extract of N. entophytum showed lower activity. More or less similar results were reported by Piccardi et al. (2000) recorded that the bioactivity of Nostoc spp. was equally distributed between lipophilic and hydrophilic extracts and was mostly directed against Penicillium expansum and R. solani. El-Sheekh et al. (2006) stated that chloroform was the best solvent for extracting the active material of N. muscurum.
The antifungal activity of cyanobacteria could be attributed to their phenol content and/or polysaccharides content (Table 1). This interpretation based on the results concerning the content of these substances in the tested cyanobacteria where their antifungal effect were increased as their polysaccharides and/or phenol content increased. In agreement with our explanation, there are a number of reports by authors on the antifungal activity of phenolic substance e.g., De Cano et al. (1990) found that phenolic compounds in extracts from cells of N. muscurum significantly inhibited the growth of Candida albicans and Staphylococcus aureus. Furthermore, Samapundo et al. (2007) observed that the phenolic compounds e.g., vanillic and caffeic acid treatments caused reduction in F. verticillioides and F. proliferatum growth. Sekine et al. (2009) detected that phenolic hydroxyl compounds have antifungal activity against white- and brown-rot fungi.
With respect to polysaccharides which play important role as defense mechanism for cyanobacteria and reflect the antifungal activity of the tested cyanobacteria as demonstrated Table 1. This observation has been emphasized by Potin et al. (1999) who found that oligosaccharides from marine algae were used to protect from infections by pathogens. Cuero (1999) revealed that the antimicrobial activity of chitosan is well observed on a wide variety of microorganisms including fungi, algae and some bacteria.
Antifungal activity of the tested organisms under greenhouse conditions: The results show that R. solani caused soybean damping off and reduced the plant length, weight and carbohydrate contents (Fig. 3). Present results support the results obtained by Ismail and Ahmed (2000) who reported that R. solani was the most pathogenic fungus, it caused significant effects in all tested variables (pre, post-emergency damping off, survival plants and plant height) of cotton seedlings. Heydari et al. (2007) observed that R. solani induced damping off symptoms on all emerged and non emerged cotton seedling. Haikal (2008) who showed that filtrates of A. niger, F. culmorium, Penicillium sp. and R. solani inhibited seed germination and seedling development of soybean due to their toxic metabolites in the media in which they were grown. Hwang et al. (2009) recorded that the height, shoot vigour and shoot dry mass of Rhodiola rose were significantly reduced by R. solani infection. Abdullah (2008) stated that R. solani decreased total carbohydrate content of wheat and barley. El-Daly and Haikal (2006) shown that the soil infection with 3% (w/w) R. solani drastically lowered the total carbohydrates of Zea mays.
It could be deduced from the previous mentioned data that T. harzianum and G. virens were the most effective antagonistic fungi to control by Rhizoctonia solani under laboratory conditions (Table 1) so we used the tested species under greenhouse conditions while 3% (w/w) of T. harzianum or G. virens reduced the post-emergency damping-off caused by R. solani and increased the survival rates of seedling. Present results are in agreement with Bazgir and Okhovat, (1996) who reported that the inoculation of T. harzianum to the soil one month before sowing reduced the level of R. solani on Phaseolus vulgaris beans. Trichoderma spp. or G. virens grew on the bran suppressed the spread of R. solani and significantly reduced its inoculum potential (Lewis et al., 1998). Trichoderma spp., Gliocladium spp. and actinomycetes were plays a key role in the sustainability of agriculture systems and indicates the level of health of soil (Gil et al., 2009).
Trichoderma harzianum was more effective than G. virens in controlling the pathogenic effect of Rhizoctonia solani in vitro and under greenhouse conditions. Our results are in conformity with those of Hanson and Howell (2002) who explained that G. virens have good biocontrol activity against Rhizoctonia solani on cotton but lack some of the commercially desirable characteristics found in Trichoderma species. T. harzianum gave maximum protection of the disease (72.72%) while G. virens and Aspergillus sp. were found to be the least effective in controlling root rot of mungbean Under greenhouse conditions (Singh and Chand, 2006).
The results present in Fig. 3 show that the application of antagonistic fungi to infected soybeans at 3% (w/w) increased the length, weight and carbohydrate accumulation of infected soybeans. It could be deduced from the obtained data that G. virens and T. harzianum act as stimulator for infected soybean elongation and weight as compared to untreated infected control. The stimulation effect of the tested fungi differs according to fungal species which correlated with antagonistic ability which confirmed previously in vitro. Our results are in agreement with Chen et al. (1996) who reported that the increased of carbohydrate content might be correlated to increase in growth rate due to the effect of stimulatory effect of the antagonistic fungi. De Paula Junior (2002) stated that T. harzianum increased bean growth in the presence of R. solani. Grosch et al., 2006) stated that Trichoderma sp. either partly or completely controlled the dry mass loss of lettuce caused by R. solani. Biological control agents T. harzianum or B. subtilis or both initiated the increase of carbohydrate content of Z. mays infected with 3% (w/w) R. solani (El-Daly and Haikal, 2006). El-Mohamedy and El-Baky (2008) detected that T. harzianum stimulated carbohydrate accumulation in the infected pea with R. solani. Trichoderma have a strong aggressiveness against phytopathogens and produce trichotoxins that could inhibit plant pathogen and promote plant growth (Gachomo and Kotchoni, 2008).
Data in (Fig. 3g, h) show that the protein and nitrogen contents of infected soybeans were decreased by addition of 3% T. harzianum or G. virens to the soil as compared with infected control under greenhouse conditions. These results are more or less similar to that reported by Naseby et al. (2000) who stated that Trichoderma strains reduced the activity of C and N cycle enzymes in pea. Inbar et al. (1994) reported that T. harzianum caused non significant changes in N content of cucumber and pepper.
The obtained data showed that the experimental cyanobacteria were able to inhibit the post-damping off effect of Rhizoctonia solani and increase the survival rate of soybean seedling under greenhouse conditions (Fig. 3a), hence N. entophytum and N. muscurum increased the number of survival seedling as compared with infested control. This positive effect may be due to their antifungal activity as demonstrated previously in vitro. In this context, Kulik (1995) mentioned that filtrates or cell extracts from cyanobacteria applied to seeds as protectants against damping-off fungi such as Fusarium sp., Pythium sp. and R. solani. De Caire et al. (1990) reported that extracellular products from N. muscurum are promising as a biological control of soybean seedlings damping off. Moore (1996) showed that Nostoc sp. (GSV 224) has potent fungicidal activity and may have use in the treatment of resistant fungal-induced diseases of domestic plants and agricultural crops.
Length, weights and carbohydrate contents of infected soybeans treated with of Nostoc entophytum and N. muscurum (Fig. 3b-f) showed an increase than untreated infected soybeans under greenhouse conditions. This increase may result from the effect of antifungal activity of cyanobacteria which suppressed the toxic effect of Rhizoctonia solani as demonstrated previously under laboratory conditions (Table 1). Very little data have been published on the effect of cyanobacteria on growth parameter of infected plant with Rhizoctonia solani. These results are in agreement with those obtained by Tiedemann et al. (1980) who found that plant biomass yield which inoculated with blue green algae were significantly greater than with the control treatment. Ordog (1999) found that the extract of cyanobacteria contain a special set of biologically active compounds including plant growth regulators which increased root and shoot development. Maqubela et al. (2009) stated that Nostoc spp. inoculation increased maize dry matter. The carbohydrate content of tomato (Lycopersicon esculentum L.) was increased by Nostoc spp. (Al-Khiat, 2009). The above mentioned stimulations in carbohydrate content of the different plant by cyanobacteria could be attributed to the stimulation of photosynthetic process by some factors present in such organisms.
With regard to the effect of tested cyanobacteria on protein and nitrogen contents of infected soybeans under greenhouse conditions, the obtained results show that the nitrogen content of infected soybeans was reduced by application of N. entophytum and N. muscurum (Fig. 3h), although cyanobacteria can fix N in soil (Metting, 1981). These results are in conformity with Adam (1999) who stated that N. muscurum improved the growth and nitrogen contents of noninfected wheat, sorghum, maize and lentil. Al-Khiat (2009) recorded that the tomato (Lycopersicon esculentum L.) protein content increased by Nostoc sp. It could be deduced from the above mentioned data that the infected plant failed to uptake nitrogen from the soil.
CONCLUSION
This work is endeavor for utilization of some antagonistic fungi and some cyanobacterial species as antifungal agent against Rhizoctonia solani which the causal agent of soybeans rot root. Our results indicated that the efficiency of the tested biological treatments (antagonistic fungi and cyanobacteria) for controlling Rhizoctonia solani under laboratory and greenhouse conditions, the degree of efficiency is different according to the types of biological treatments. All tested biological treatments are effective for decreasing the post-emergency damping off of soybeans caused by Rhizoctonia solani and increased some growth parameter e.g., soybean severity, length, weights and carbohydrate content. On the other hand, they have negative effect on protein and nitrogen contents of infected soybeans as compared with untreated infected control under greenhouse conditions. The antifungal activity induced by such biotic factors could be attributed to phenol and polysaccharides content.
REFERENCES
- Adam, M.S., 1999. The promotive effect of the cyanobacterium Nostoc muscorum on the growth of some crop plants. Acta Microbiol. Pol., 48: 163-171.
Direct Link - Bauer, A.W., W.M.M. Kirby, J.C. Sherris and M. Turck, 1966. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol., 45: 493-496.
CrossRefPubMedDirect Link - Bazgir, E. and M. Okhovat, 1996. Biological control of Rhizoctonia solani, the casual agent of damping-off and seed rot of bean by certain isolates of antagonistic fungi. Iran. J. Agricult. Sci., 27: 89-98.
Direct Link - Bohlooli, A., S.M. Okhowat and M. Javan-Nikkhah, 2005. Identification of anastomosis group of Rhizoctonia solani, the causal agent of seed rot and damping-off of bean in Iran. Commun. Agric. Applied Biol. Sci., 70: 137-141.
PubMedDirect Link - Borowitzka, M.A., 1995. Microalgae as sources of pharmaceuticals and other biologically active compounds. J. Applied Phycol., 7: 3-15.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Bradley, C.A., G.L. Hartman, L.M. Wax and W.L. Pedersen, 2002. Influence of herbicides on Rhizoctonia root and hypocotyl rot of soybean. Crop Product., 21: 679-687.
CrossRef - Brimmer, T. and G.J. Boland, 2003. A review of the non-target effects of fungi used to biologically control plant diseases. Agric. Ecosyst. Environ., 100: 3-16.
CrossRefDirect Link - Chen, F., Y. Zhang and S.Y. Guo, 1996. Growth and phycocyanin formation of Spirulina plantensis in photoheterotrophic cultures. Biotechnol. Lett., 18: 603-608.
CrossRef - El-Daly, F.A. and N. Haikal, 2006. Role of biological control on some physiological aspects of Zea mays infected by Rhizoctonia solani. J. Applied Sci., 6: 2794-2798.
CrossRefDirect Link - El-Sheekh, M.M, M.E.H. Osman, M.A. Dyab and M.S. Amer, 2006. Production and characterization of antimicrobial active substance from the cyanobacterium Nostoc muscorum. Environ. Toxicol. Pharmacol., 21: 42-50.
CrossRef - Fayzalla, E.A., E. El-Barougy and M.M. El-Rayes, 2009. Control of soil-borne pathogenic fungi of soybean by biofumigation with mustard seed meal. J. Applied Sci., 5: 2272-2279.
CrossRefDirect Link - Gachomo, W.E. and S.O. Kotchoni, 2008. The use of Trichoderma harzianum and T. viride as potential biocontrol agents against peanut microflora and their effectiveness in reducing aflatoxin contamination of infected kernels. Biotechnology, 7: 439-447.
CrossRefDirect Link - Gil, S.V., S. Pastor and G.J. March, 2009. Quantitative isolation of biocontrol agents Trichoderma spp, Gliocladium spp. and actinomycetes from soil with culture media. Microbiol. Res., 164: 196-205.
CrossRef - Grosch, R, K. Scherwinski, J. Lottmann and G. Berg, 2006. Fungal antagonists of the plant pathogen Rhizoctonia solani: Selection, control efficacy and influence on the indigenous microbial community. Mycol. Res., 110: 1464-1474.
CrossRefPubMedDirect Link - Haikal, N.Z., 2008. Effect of filtrates of pathogenic fungi of soybean on seed germination and seedling parameters. J. Applied Sci. Res., 4: 48-52.
Direct Link - Hanson, L.E. and C.R. Howell, 2002. Biocontrol efficacy and other characteristics of protoplast fusants between Trichoderma koningii and T. virens. Mycol. Res., 106: 321-328.
CrossRef - Heydari, A., I.J. Misaghi and G.M. Balestra, 2007. Pre-emergence herbicides influence the efficacy of fungicides in controlling cotton seedling damping-off in the field. Int. J. Agric. Res., 2: 1049-1053.
CrossRefDirect Link - Hwang, S.F., H.U. Ahmed, K. Ampong-Nyarko, S.E. Strelkov, R.J. Howard and G.D. Turnbull, 2009. Causal agents of root rot and the effect of vesicular-arbuscular mycorrhizal fungi in seedlings of Rhodiola rosea in Alberta, Canada. Plant Pathol. J., 8: 120-126.
CrossRefDirect Link - Inbar, J., M. Abramsky, D. Cohen and I. Chet, 1994. Plant growth enhancement and disease control by Trichoderma harzianum in vegetable seedlings grown under commercial conditions. Eur. J. Plant Pathol., 100: 337-346.
CrossRefDirect Link - Jindal, K.K. and R.N. Singh, 1975. Phenolic content in male and female Carica papaya: A possible physiological marker sex identification of vegetative seedlings. Physiol. Plant., 33: 104-107.
CrossRefDirect Link - Kalai, K., G. Giczey, A. Meszaros, F. Denes and E. Balazs, 2008. Trichoderma chitinase gene expression confers mould resistance. ISHS Acta Hort., 725: 783-790.
Direct Link - Katircioglu, H., Y. Beyatli, B. Aslim, Z. Yuksekdag and T. Atici, 2006. Screening for antimicrobial agent production of some microalgae in freshwater. Intern. J. Microbiol., 2: 1-9.
Direct Link - Khan, N.H., M. Rahman and M.S. Nur-e-Kamal, 1988. Antibacterial activity of Euphorbia thymifolia Linn. Indian J. Med. Res., 87: 395-397.
PubMedDirect Link - Kulik, M.M., 1995. The potential for using cyanobacteria (blue-green algae) and algae in the biological control of plant pathogenic bacteria and fungi. Eur. J. Plant Pathol., 101: 585-599.
Direct Link - Lozovaya, V.V., A.V. Lygin, S. Li, G.L. Hartman and J.M. Widholm, 2004. Biochemical response of soybean roots to Fusarium solani f. sp. glycines infection. Crop Sci., 44: 819-826.
Direct Link - Maqubela, M.P., P.N.S. Mnkeni, O.M. Issa, M.T. Pardo and L.P. D'Acqui, 2009. Nostoc cyanobacterial inoculation in South African agricultural soils enhances soil structure, fertility and maize growth. J. Plant Soil, 315: 79-92.
CrossRefDirect Link - Meyera, M.C., C.J. Buenob, N.L. de Souzab, and J.T. Yorinoric, 2006. Effect of doses of fungicides and plant resistance activators on the control of Rhizoctonia foliar blight of soybean, and on Rhizoctonia solani AG1-IA in vitro development. Crop Protect., 25: 848-854.
CrossRef - Hermosa, M.R., I. Grondona, E.A. Iturriaga, J.M. Diaz-Minguez, C. Castro, E. Monte and I. Garcia-Acha, 2000. Molecular characterization and identification of biocontrol isolates of Trichoderma spp. Applied Environ. Microbiol., 66: 1890-1898.
CrossRefDirect Link - Moore, R.E., 1996. Cyclic peptides and depsipeptides from cyanobacteria: A review. J. Ind. Microbiol., 16: 134-143.
CrossRefDirect Link - Nair, R., T. Kalariya and S. Chanda, 2005. Antibacterial activity of some selected Indian medicinal flora. Turk. J. Biol., 29: 41-47.
Direct Link - Naseby, D.C., J.A. Pascual and J.M. Lynch, 2000. Effect of biocontrol strains of Trichoderma on plant growth, Pythium ultimum populations, soil microbial communities and soil enzyme activities. J. Applied Microbiol., 88: 161-169.
CrossRef - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Noaman, N.H., A.F.M. Khaleafa and S.H. Zaky, 2004. Factors affecting antimicrobial activity of Synechococcus leopoliensis. Microbiol. Res., 159: 395-402.
CrossRefDirect Link - Ordog, V.F., 1999. Beneficial effects of microalgae and cyanobacteria in plant/soil systems, with special regard to their axing and cytokinin like activity. Proceedings of International Workshop and Training Course on Microalgal Biology and Biotechnology Mosonmagyarovar, June 13-26, Hungary, pp: 43-43.
- Piccardi, R., A. Frosini, M.R. Tredic and M.C. Margheri, 2000. Bioactivity in free-living and symbiotic cyanobacteria of the genus Nostoc. J. Applied Phycol., 12: 543-547.
CrossRefDirect Link - Potin, P., B. Kamal, K. Frithjof and K. Bernard, 1999. Oligosaccharide recognition signals and defence reactions in marine plant-microbe interactions. Curr. Opin. Microbiol., 2: 276-283.
CrossRef - Samapundo, S., B. De Meulenaer, D. Osei-Nimoh, Y. Lamboni, J. Debevere and F. Devlieghere, 2007. Can phenolic compounds be used for the protection of corn from fungal invasion and mycotoxin contamination during storage? Food Microbiol., 24: 465-473.
CrossRef - Sekine, N., T. Ashitani, T. Murayama, S. Shibutani, S. Hattori and K. Takahashi, 2009. Bioactivity of latifolin and its derivatives against termites and fungi. J. Agric. Food Chem., 57: 5707-5712.
CrossRefDirect Link - Shi, Y., J. Sheng, F. Yang and Q. Hu, 2007. Purification and identification of polysaccharide from Chlorella pyrenoidosa. Food Chem., 103: 101-105.
CrossRef - Singh, S. and H. Chand, 2006. Screening of bioagents against root rot of mung bean caused by Rhizoctonia solani. Mol. Plant Microbe Interact., 18: 710-721.
PubMed - Soylu, S., E.M. Soylu, S. Kurt and O.K. Ekici, 2005. Antagonistic potentials of rhizosphere-associated bacterial isolates against soil-borne diseases of tomato and pepper caused by Sclerotinia sclerotiorum and Rhizoctonia solani. Pak. J. Biol. Sci., 8: 43-48.
CrossRefDirect Link - Szekeres, A, K. Laszlo, A. Zsuzsanna, K. Ferenc and M. Laszl, 2004. Isolation and characterization of protease overproducing mutants of Trichoderma harzianum. FEMS Microbiol. Lett., 233: 215-222.
CrossRef - Tiedemann, A.R., W. Lopushinsky and P.A. Roger, 1980. Plant and soil responses to a commercial blue-green algae inoculants. Soil Biol. Biochem., 12: 471-475.
CrossRef - Topps, T.H. and R.L. Wain, 1957. Investigation on fungicides III. The fungitoxicity of 3- and 5 alkyl salicylanilide and P-chloronilines. Ann. Applied Biol., 45: 506-514.
CrossRef - Vlachos, V., A.T. Critchley and A. von Holy, 1996. Establishment of protocol for testing antimicrobial activity in southern African macroalgae. Microbios., 88: 115-123.
PubMedDirect Link - Washington, W.S., S. Engleitner S, G. Boontjes and N. Shanmuganathan, 1999. Effect of fungicides, seaweed extracts, tea tree oil and fungal agents on fruit rot and yield in strawberry. Aust. J. Exp. Agric., 39: 487-494.
CrossRefDirect Link - Whipps, J.M., 2001. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot., 52: 487-511.
PubMedDirect Link - Yadav, J., J.P. Verma and K.N. Tiwari, 2011. Plant growth promoting activities of fungi and their effect on chickpea plant growth. Asian J. Biol. Sci., 4: 291-299.
CrossRefDirect Link - Rippka, R., J. Deruelles, J.B. Waterbury, M. Herdman and R.Y. Stanier, 1979. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology, 111: 1-61.
CrossRefDirect Link - Weller, D.M., J.M. Raaijmakers, B.B.M. Gardener and L.S. Thomashow, 2002. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathol., 40: 309-348.
CrossRefPubMedDirect Link