
Gh. Hassan Dar
Division of Plant Pathology, S.K. University of Agricultural Sciences and Technology of Kashmir, Shalimar, Srinagar, Jammu and Kashmir-191 121, India
M.A. Beig
Division of Plant Pathology, S.K. University of Agricultural Sciences and Technology of Kashmir, Shalimar, Srinagar, Jammu and Kashmir-191 121, India
F.A. Ahanger
Division of Plant Pathology, S.K. University of Agricultural Sciences and Technology of Kashmir, Shalimar, Srinagar, Jammu and Kashmir-191 121, India
Nadeem A. Ganai
Division of Plant Pathology, S.K. University of Agricultural Sciences and Technology of Kashmir, Shalimar, Srinagar, Jammu and Kashmir-191 121, India
M. Ashraf Ahangar
Division of Plant Pathology, S.K. University of Agricultural Sciences and Technology of Kashmir, Shalimar, Srinagar, Jammu and Kashmir-191 121, India
Asian Journal of Plant Pathology
Year: 2011 | Volume: 5 | Issue: 2 | Page No.: 62-74







ABSTRACT
The present study was aimed to identify root rot pathogens of blue pine (Pinus wallichiana) in Kashmir and develop appropriate eco-friendly disease management strategy. During nursery surveys, Fusarium oxysporum, Rhizoctonia solani and Macrophomina phaseolina were found root rot incitants with isolation frequency of 47.3, 29.7 and 13.0%, respectively. Locally isolated antagonists inflicted 33.0-73.3 and 29.5-70.8% mycelial growth inhibition in F. oxysporum and R. solani, respectively, with Trichoderma harzianum and T. viride proving most effective. The mycorrhizal fungi, Pisolithus tinctorius and Laccaria laccata, significantly inhibited the growth of R. solani and F. oxysporum by 46.2 and 45.4 and 44.7 and 43.7%, respectively. Bioagents significantly improved seedling biomass and root/shoot length. Mycorrhizal plants showed 5-13 fold higher rhizosphere phosphatase activity than non-mycorrhizal ones. Four effective fungal bioagents, inoculated individually and in combination with pathogen under nursery conditions, significantly improved seedling biomass and height with maximum gain by P. tinctorius and L. laccata. Rhizoctonia infection decreased biomass and seedling height by 32.6 and 35.4%, whereas bioagents mitigated the pathogenic effect. The bioagents in R. solani/F. oxysporum-infected soil significantly improved seedling biomass and height over pathogen treatments alone. P. tinctorius and L. laccata exhibited 44.2 and 39.1% root colonization in comparison to 19.5-24.2% in presence of pathogens. The study revealed that bioagents, especially mycorrhizae, effectively mitigate root rot in blue pine and can be efficiently exploited in integrated disease management module.
PDF Abstract XML References Citation
Received: March 24, 2011;
Accepted: May 18, 2011;
Published: July 12, 2011
How to cite this article
Gh. Hassan Dar, M.A. Beig, F.A. Ahanger, Nadeem A. Ganai and M. Ashraf Ahangar, 2011. Management of Root Rot Caused by Rhizoctonia solani and Fusarium oxysporum in Blue Pine (Pinus wallichiana) Through use of Fungal Antagonists. Asian Journal of Plant Pathology, 5: 62-74.
URL: https://scialert.net/abstract/?doi=ajppaj.2011.62.74
URL: https://scialert.net/abstract/?doi=ajppaj.2011.62.74
INTRODUCTION
The Himalayan mountainous ranges harbour four out of the six indigenous pine species of Indian subcontinent, viz., Pinus wallichiana, P. roxburghii, P. gerardiana and P. kesiya. Of these, blue pine (Pinus wallichiana Jack.) has its native habitat spread over Eastern Afghanistan, South-eastern Tibet and China to North Burma and is mostly found in Himalayas at an elevation of 1500 to 3500 m m.a.s.l. The tree is widely distributed in Jammu and Kashmir, Himachal Pradesh and Gharwal hills in the West, spreading towards Bhutan with sporadic distribution in Arunachal Pradesh in the east. Of the total 20,230 km2 area under forests in Jammu and Kashmir State, conifers cover 40.87% area out of which blue pines occupy 9.73% area (Beig et al., 2008a).
Blue pine, locally known as Kail pine, faces several constraints in its successful regeneration in field. The plants are often exposed to persistent pathogenic attacks, particularly those inciting root rot and wilt diseases, at primary stages of plant establishment. Root rot, wilt and die-back on container-grown conifers such as spruce and pine is major problem since 1990’s in Europe (Lilja et al., 2010). The root rot fungi which pose serious threat to forest nurseries include the species of Fusarium, Phytophthora, Pythium, Rhizoctonia, Macrophomina and Cylindrocladium (Huang and Kuhlman, 1990; Asiegbu et al., 1999; Wafaa and Haggag, 2002). These pathogens often invade terminal unsuberized roots of young seedlings and cause late damping off or root rot/wilt thereby kill the host. The fungi penetrate into root epidermal cell wall, grow intercellularly, decompose cell wall constituents and persist by metabolising cell contents. Root rot is world-wide a serious problem in pine seedlings and serious losses due to this disease have been reported from Ontario, Canada, USA and many European countries (Greifenghagen et al., 1991; Lilja and Rikala, 2000). In view of highly devastating nature of root rot pathogens, effective disease management is essential to raise healthy pine seedlings for successful implementation of reforestation and afforestation programmes.
Several approaches involving fungicide use and cultural measures have been adopted by the nursery growers to reduce the root rot incidence; yet the disease continues to assume serious threat to conifers (Shah et al., 1999). The use of biocontrol agents is presently gaining momentum as a supplement to chemical treatment in integrated disease management module. The effective use of antagonistic bacteria, actinomycetes and fungi as biocontrol agents against several soil-borne pathogens have been demonstrated in several field and horticultural crops (Yobo et al., 2010). The free-living rhizosphere micro-organisms such as Trichoderma, Gliocladium, Penicillium, Pseudomonas, Bacillus, Azotobacter, Azospirillum, etc., favourably influence the plant growth directly or indirectly; whereas mycotrophy benefits the host plant by enhancing the root capacity to absorb nutrients, extracting the nutrients beyond depletion zones in soil and protecting the root from pathogenic invasions (Eziashi et al., 2006; Gacitua et al., 2009; Lu and Huang, 2010; Heydari and Pessarakli, 2010). The fungal antagonists may compete for an ecological niche by consuming available nutrients and by secreting a spectrum of biochemicals effective against various fungal pathogens. These biochemicals may include cell wall-degrading enzymes, siderophores, chelating iron, a wide variety of volatile and non-volatile antibiotics, etc. (Susi et al., 2011). Widyastuti et al. (2003) and Benitez et al. (2004) speculated that indirect and direct defensive mechanisms of Trichoderma species act coordinately. Accordingly the success in biocontrol process depends on the antagonisitic strain involved, the antagonized fungus, the crop plant and the environmental conditions, including nutrient availability, pH, temperature and iron concentration. Root drench of potted Pinus radiata with Trichoderma atroviride isolate R32 not only enhanced seedling root biomass and stem diameter but also induced systemic resistance to Diplodia pinea and reduced dieback incidence by 20% compared to untreated controls (Reglinski et al., 2011).
Symbionts, such as mycorrhizal fungi and free living organisms form integral components of pine rhizosphere, an area exhibiting all kinds of antagonistic, parasitic and growth promoting interactions (Taylor and Alexander, 2005). Mycorrhiza reportedly aid young seedlings in their early survival and establishment through intricate and complex system of hyphal networks thereby not only ensures the sustained nutrient supply but also provides protection against invading pathogens (Podila and Douds, 2001; Jha et al., 2008). The culture filtrate of mycorrhiza, Suillus cdlinius Hebeloma mesophaeum and Paxillus sp., reportedly exhibit antagonistic effect on the mycelial growth and spore germination of Fusarium oxysporum and Pythium vexans and the antifungal activity has been attributed to oxalic acid production (Quarraqi et al., 2005; Yamaji et al., 2005). Inoculation of Pisolithus tinctorius in Pinus densiflora significantly increased dry matter and stem diameter when compared to non-inoculated seedlings (Choi et al., 2005). Paxillus involutus effectively controlled root rot caused by Fusarium oxysporum and Fusarium moniliforme in red pine (Pal and Gardener, 2006). Efforts are underway to improve the quality of forest nursery seedlings through inoculation of suitable mycorrhizal strain in association with other compatible bioagents.
The perusal of literature reveals that some work has been done on the management of root rot diseases in conifers other than blue pine (Enebak et al., 1990). From India, meagre information about the causal pathogens responsible, symptomology and management of diseases is available for conifers (Kaushik et al., 2002; Bisht et al., 2003). In view of the importance of root rot disease in blue pine and the damage inflicted by it, there is urgent need to develop a suitable disease management strategy which may include biological control as an important component. Therefore, the present investigation was undertaken to assess the bio-efficacy of fungal antagonists in the management of root rot disease on blue pine under temperate conditions of Kashmir.
MATERIALS AND METHODS
The root rot pathogens were isolated from diseased roots, collected during the survey of blue pine (Pinus wallichiana) nurseries in years 2007 and 2008. Three to four surface-sterilized diseased root bits of 3-5 mm size were aseptically transferred to Potato Dextrose Agar (PDA) medium and plates incubated at 25±2oC for mycelial growth. The cultures were purified by hyphal tip method (Dasgupta, 1988). Various cultural and morphological characteristics of isolated fungi were recorded by visual and microscopic examinations. Morphological characteristics of isolated fungi were compared with standard descriptions given by Nelson et al. (1983) and Sneh et al. (1998). The fungal/bacterial antagonists and ectomycorrhiza were isolated from the rhizosphere of blue pine trees by dilution plate method on PDA and Modified Melin Norkan’s agar (MMN) media, respectively (Marx, 1969; Rangeshwaran and Prasad, 2000). The cultures were purified by single spore/hyphal tip method. The identification of isolated fungi was done on the basis of cultural and morphological characteristics (Arx, 1981). The isolated ectomycorrhiza were identified on the basis of culturo-morphological characteristics and identified with the help of standard descriptions (Godbout and Fortin, 1985; Lakhanpal, 1988).
In vitro assessment of biocontrol agents: The antagonistic activity of isolated rhizosphere fungi/bacterium and mycorrhizae against root rot pathogens was assessed by dual culture technique using PDA and MMN agar media, respectively (Dhingra and Sinclair, 1985). Treatments were replicated four times in a completely randomized design and incubated at 25±2°C for 8 days. Radial growth was recorded on 8th day of incubation and mycelial inhibition calculated as per Vincent (1947).
The morphology of hyphae in interaction zone was observed under light (10x) microscope. Clear and characteristically parasitized hyphae were examined under high magnification power (40x). Based on the growth and mycoparasitic nature, biocontrol agents were grouped into various categories as per the scale given byMunshi and Dar (2004). The antifungal activity of bacterial antagonist was assessed by dual culture technique according to the method given by Dennis and Webster (1971), using PDA medium. Control plates inoculated with pathogen alone were also maintained. The treatment plates, replicated four times, were incubated at 28±2°C in completely randomized design. Mycelial growth of pathogen and inhibition zone was measured after 72 h of incubation.
In vivo evaluation of antagonists: The efficacy of antagonist under in vivo conditions was assessed as per the method described earlier (Ahangar et al., 2011). The treatments were replicated 10 times and arranged in a completely randomized block design in a glasshouse at 26±3°C. The seedlings were gently uprooted after 3 months to assess fresh plant biomass and mycorrhizal development. The entire root systems were examined under a stereomicroscope to count the number of mycorrhizal short roots (Daughtridge et al., 1986). The mycorrhizal colonization was assessed by counting the total number of mycorrhizal short roots formed by inoculated fungi against the total number of short roots observed. Root and shoot length of seedlings was measured and alkaline acid phosphatase activity of rhizosphere soil was estimated as per the method of Tabatabai and Bremner (1969). The amount of ρ-nitrophenol (PNP) released was calculated in reference to the standard curve prepared by using different concentrations of PNP. The enzyme activity was expressed in terms of μM PNP released g-1 soil h-1.
Nursery studies on pathogen-antagonist interaction: The most effective five fungal antagonists, including two ectomycorrhiza, were evaluated individually and in combination with major pathogens. The pathogenic inoculums of non-mycorrhizal fungus were incorporated @ 10 g kg-1 mixture to ensure sick soil formation. Biocontrol agents, multiplied on wheat bran, were added @ 10 g kg-1 potting mixture (with an inoculum load of 1x109 cfu g-1) as per treatment. Ectomycorrhiza were multiplied in vermiculite-peat moss based carrier added to sick soil @ 15 mL bag-1. The antagonist additions to sick soil were made 15 days before seed sowing. Non-sick soil and sick soils without antagonist inoculation served as controls. The surface sterilized and stratified healthy seeds of blue pine were sown 3 per bag of sterilized potting mixture of one kg in a 1.5 kg capacity plastics bag. After germination seedlings were thinned out to one per bag. Each treatment replicated 10 times was arranged in a CRD in a greenhouse at 25±3°C and irrigated with sterile water as and when required. No fertilizers or protective chemicals were applied throughout the study. Ectomycorrhizal infection in roots was estimated and roots examined under a stereomicroscope to count the number of mycorrhizal short roots as suggested by Beckjord et al. (1984).
The fresh plant biomass and seedling height were estimated 90 and 180 days after seedling emergence. Root colour intensity and root rot index was measured on the basis of root area affected according to the root rot index rating scale described by Purkayastha et al. (1981). The data were subject to analysis of variance and means compared using Duncan’s new multiple range test (Gomez and Gomez, 1984). Arcsine and square transformations were employed wherever applicable. The data analysis was carried out using Statistical Packages for Social Sciences (SPSS ver. 11.5 Chicago USA for windows).
RESULTS AND DISCUSSION
Root rot pathogens of pines: During preliminary survey the roots of blue pine seedling showing root rot and wilt symptoms, collected from forest nurseries in Anantnag and Baramulla districts of Jammu and Kashmir, were found infected by three root rot causing fungi viz., Fusarium oxysporum f.sp. pini Schlecht. Synd. and Hans., Rhizoctonia solani Kuhn. and Macrophomina phaseolina (Tassi.) Goid. Besides these, some saprophytic fungi were isolated from the affected root portions. Fusarium oxysporum was isolated from all the surveyed nurseries with an overall isolation frequency of 47.3% which was followed by Rhizoctonia solani and Macrophomina phaseolina with overall isolation frequencies of 29.7 and 13.0%, respectively and the rest 10% were the species of Mucor, Rhizophus, Penicillium, Aspergillus and Trichoderma. These observations are in line with Pinto et al. (2006) who recorded 44.6% isolation frequency of F. oxysporum from colonized roots of Pinus sylvestris seedlings and 0.3% isolation frequency of R. solani from the diseased roots of Picea abies seedlings from Uppsala Sweden. Lilja et al. (1995) and Stepniewska-Jarosz et al. (2006) frequently isolated the above pathogens from diseased conifer roots, cone scales and seeds of Scots pine from Florida and Poland forest nurseries, respectively. Fusarium species, especially F. oxysporum, reportedly is highly potential root rot pathogens in many forest nurseries (Enebak et al., 1990).
Antagonistic isolates: Three non-mycorrhizal fungal antagonists Trichoderma viride, T. harzianum and Gliocladium roseum and four mycorrhizal fungi Pisolithus tinctorius, Laccaria laccata, Boletus spp. and Suillus granulatus were isolated from blue pine rhizosphere. Their identification was made on the basis of morpho-cultural characteristics. L. laccata and S. granulatus have previously been isolated from the basidiomata collected from Pinus patula plantations by Reddy and Natarajan (1997) from Nilgiri hills, Tamil Nadu, India and Yamada et al. (2001) from Ibaraki, Japan. Dar et al. (2009) have reported the presence of Pisolithus tinctorius, Laccaria laccata, Boletus spp. and Suillus granulatus from Gulmarg, Bandipora and Baramulla conifer forests of Kashmir with high rhizosphere phosphatase activity and root colonizing potential.
Pathogenic growth inhibition by antagonists: In vitro evaluation of antagonists against F. oxysporum and R. solani in dual culture revealed that all the tested biocontrol agents significantly inhibited the mycelial growth of pathogens (Table 1). The mycelial inhibition on 8th day ranged from 33.0 to 73.3% and 29.5 to 70.8% for F. oxysporum and R. solani, respectively, with T. harzianum proving more effective against both the pathogens, followed by T. viride (70.5 and 64.8%) and Gliocladium roseum (50.0 and 54.7%). Trichoderma species proved highly antagonistic and exhibited strong mycoparasitic activity.
| Table 1: | In vitro effect of various antagonists on mycelial growth of Fusarium oxysporum and Rhizoctonia solani in dual culture |
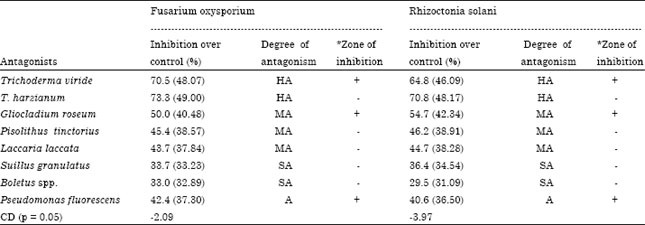 | |
| HA: Highly antagonistic, MA: Moderately antagonistic, A: Antagonistic, SA: Slow antagonistic, * +: Inhibition zone present, -: inhibition zone absent | |
They completely overgrew the host mycelia once in contact with pathogens and formed hyphal coils on pathogenic colonies. G. roseum and Pseudomonas fluorescens grew slowly and developed zone of inhibition against the pathogen. Amongst the mycorrhizal fungi Pisolithus tinctorius and Laccaria laccata inflicted significantly higher mycelial growth inhibition of 45.4 and 46.2 and 43.7 and 44.7% in F. oxysporum and R. solani, respectively. Further, L. laccata and P. tintorius were observed to be moderate antagonists as they caused rupture and twisting of pathogenic hypha followed by their gradual desiccation, protoplasm shrinkage and ultimately cell lysis. Boletus spp. and Suillus granulatus inflicted only 33.0 and 29.5 and 33.7 and 36.4% inhibition in mycelial growth of F. oxysporum and R. solani, respectively, so were categorized as slow antagonists. The growth inhibitive effects of antagonists are in agreement with Rudresh et al. (2005) who observed 72.1 and 77.0% growth inhibition in R. solani and F. oxysporum, respectively, by T. harzianum and T. viride which also exhibited strong mycoparasitic activity and completely overgrew the host mycelia once in contact with them. Dubey (1998) observed hyphal coil or hook or appresoria formation by T. harzianum on fungal colony of Rhizoctonia solani. Trichoderma species reportedly produce chitinase and β1-3, glucanase enzymes which degrade cell wall and cause hyphal lysis of pathogens (Wu et al., 1986).
The formation of inhibition zone by T. viride, G. roseum and P. fluorescens suggests the involvement of strong antibiosis mechanism, possibly due to the production of volatile metabolites and diffusible chemicals produced by antagonist. Munshi and Dar (2004) noticed inhibition zone formation by Gliocladium sp. against Fusarium pallidoroseum. The mycoparasitic activity of Laccaria laccata against R. solani and Fusarium sp. has earlier been suggested by Zhao and Kuo (1988).
Plant growth improvement by antagonists: Preliminary in vivo evaluation of mycorrhizal and non-mycorrhizal fungal bioagents in improving pine seedling growth was assessed in pot culture experiments. All the test biocontrol agents significantly improved root and shoot length of pine seedlings and yielded higher biomass than uninoculated control, as observed 90 days after seedling emergence (Table 2). L. laccata and P. tinctorius depicted significantly higher fresh plant biomass of 0.99 and 0.97 g plant-1, respectively, followed by T. harzianum (0.85 g plant-1) and T. viride (0.83 g plant-1) in comparison to un-inoculated control (0.62 g plant-1).
| Table 2: | In vivo effect of fungal antagonists, including ectomycorrhiza, on plant growth, root colonization and rhizosphere acid phosphatase activity in blue pine seedlings |
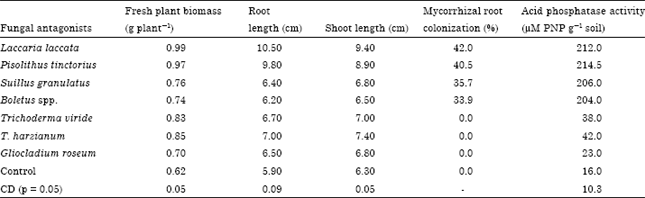 | |
| Observations taken 3 months after inoculation | |
Similar trend was noticed in case of root and shoot length with significantly higher length of 10.5 and 9.4 cm, respectively, in L. laccata in comparison to the respective values of 5.9 and 6.3 cm in un-inoculated control. This was followed P. tinctorius, T. harzianum and T. viride. The findings are in agreement with Villeneuve et al. (1991) who observed 40% growth increase in Douglas-fir seedlings due to the inoculation of L. laccata and Reddy and Natarajan (1997) who reported 38.0 and 186.5% increase in seedling height and shoot dry weight of Pinus patula due to L. laccata inoculation after 8 months. It appears that mycorrhizal fungi imparted protection, induced resistance or released antimicrobial compounds to favour plant growth. Fungal sheath around the roots seemed to have restricted the fungal advancement into mycorrhizal cortex. Protection of conifer seedlings against Fusarium spp. due to L. laccata has been attributed to the possible production of antifungal phenol compounds by the host in presence of mycorrhizal species (Chakravarty et al., 1991). Farquhar and Peterson (1990) showed that Pinus resinosa seedlings inoculated with Paxillus involutus had induced resistance to F. oxysporum.
Mycorrhizal colonization and rhizosphere phosphatase activity: The mycorrhizal root colonization and rhizosphere phosphatase activity was higher in mycorrhiza inoculated treatments. The tested ectomycorrhizal fungi colonized 33.9-42.0% roots in 90 days, developed symbiotic association with pine seedling roots and improved mycorrhizal short root formation (Table 2). Significantly high mycorrhizal root colonization was observed in pine inoculated with L. laccata (42.0%) with better rhizosphere phosphatase activity of 212.0 μM PNP g-1 soil. However, P. tinctorius was at par with L. laccata with root colonization of 40.5% and rhizosphere phosphatase activity of 214.5 μM PNP g-1 soil. In case of soils inoculated with non-mycorrhizal bioagents and untreated control the phosphatase activity was significantly very low ranging from 16.0 to 42.0 μM PNP g-1 soil. Mycorrhizal plants exhibited 5-13 fold higher rhizosphere phosphatase activity than non-mycorrhizal ones. Acid phosphatase is solely of extracellular origin and is involved in the mineralization of organic phosphates. The phosphatase enzymes solubilize insoluble forms of phosphorus and other nutrients not readily available to plant roots. Dunabeitia et al. (2004) and Beig et al. (2008b) observed greater acid phosphatase activity in the rhizosphere of mycorrhizal plants than non-mycorrhizal ones. T. harzianum has also the ability to solubilize many plant nutrients including rock phosphate from their solid phase compounds by their enzymatic activity (Altomare et al, 1999). The synergistic effect of biocontrol agents in combined inoculation resulted higher phosphatase activity in the rhizosphere of blue pine seedlings.
Plant growth improvement in presence of pathogens: Under field nursery conditions four effective fungal biocontrol agents including 2 effective ectomycorrhiza were inoculated individually and in combination with root rot pathogen (F. oxysporum or R. solani). All the test bioagents significantly improved plant growth in terms of biomass and seedling height in blue pine whereas presence of root-rot pathogens reduced the overall plant growth and biomass (Table 3). The bioagent inoculated plants had higher biomass of 0.51-0.60 and 0.98-1.15 g plant-1 on 90 and 180th days growth, respectively, in comparison to 0.46 and 0.84 g plant-1 in uninoculated control. Maximum gain was observed in case of P. tinctorius followed by L. laccata. After 90 days growth, the use of biocontrol agents individually, significantly improved seedling height (5.9-6.5 cm) over uninoculated control (4.8 cm), with maximum gain in blue pine seedlings inoculated with P. tinctorius, followed by L. laccata, T. viride and T. harzianum. With the advancement in growth period, the shoot height depicted almost similar trend with inoculated plants having height of 9.2-10.6 cm as compared to 7.5 cm in unioculated control after 180 days growth.
| Table 3: | In vivo interaction of fungal antagonists with wilt pathogens, Rhizoctonia solani (R) and Fusarium oxysporum (F) on blue pine seedlings (pooled data of 2 years) |
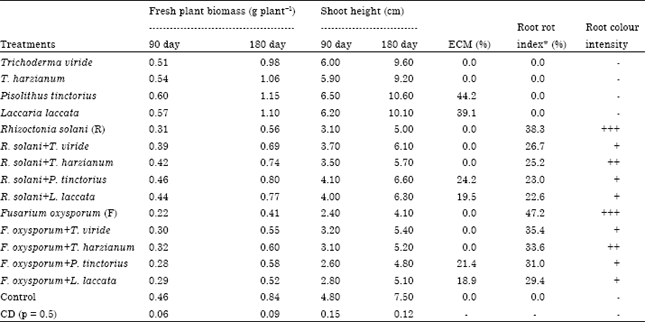 | |
| * Root rot scale: No root rot = 0; <10 percent root area affected = 0.10; 11-25 percent root area affected = 0.25; 26-50 percent root area affected = 0.50; 51-75 percent root area affected = 0.75; 76 percent root area affected = 1.00 | |
Rhizoctonia-infection resulted in less plant biomass (0.31 g plant-1) and seedling height (3.1 cm) after 90 days growth, with almost similar trend noticed after 180 days. However, in presence of bioagents the effect of R. solani was significantly reduced and seedling biomass and shoot height in pine was 0.39-0.46 g plant-1 and 3.5-4.1 cm, respectively, on 90th day of growth. The seedlings biomass and height was 0.69-0.80 g plant-1 and 5.7-6.6 cm, respectively, in Rhizoctonia + bioagent treatments as compared to 0.56 g plant-1 and 5.0 cm in R. solani treatment alone on 180th day of growth with maximum increase in R. solani + P. tinctorius followed by R. solani + L. laccata treatment. The bioagents in presence of root rot pathogen Fusarium oxysporum also promoted plant growth and mitigated the influence of pathogen (Table 3). Fusarium oxysporum infected blue pine seedlings had less biomass and shoot height of 0.22 g plant-1 and 2.4 cm, respectively, after 90 days growth and 0.41 g plant-1 and 4.1 cm after 180th days growth in comparison to unicoculated control. The bioagents in F. oxysporum-infected soils remarkably had higher plant biomass and seedling height ranging from 0.28 to 0.32 g plant-1 and 2.6 to 3.2 cm, respectively, on 90th day of growth and from 0.52-0.60 g plant-1 and 4.8-5.4 cm, respectively, on 180th day of growth with maximum improvement in biomass in F. oxysporum+T. harzianum, followed by F. oxysporum+P. tinctorius and maximum increase in plant height in F. oxysporum+T. viridi treatment followed by F. oxysporum+T. harzianum. The overall improvement in plant biomass and seedling height by bioagents may be attributed to the growth promoting and protective effects of biocontrol agents. Werner et al. (2002) observed that mycorrhizal Pinus sylvestris seedlings inoculated with Trichoderma virens produced significantly higher plant growth and biomass of needles, trunks and root than uninoculated plants. T. harzianum showed stimulatory effect on seedling growth and biomass of pine. Trichoderma species reportedly produce hormone like metabolites and release nutrients from soil or organic matter thereby facilitate better plant growth (Windham et al., 1986; Yobo et al. 2010).
Disease control by antagonists: The antagonists remarkably decreased root rot disease index and improved root colour intensity in pathogen-infected treatments (Table 3). High root rot index was observed in pine seedlings raised in either R. solani or F. oxysporum infested soil as compared to control (non-sick soil). The bioagents significantly reduced root rot from 38.3% in R solani treatment to 22.6-26.7% with maximum decrease in L. laccata followed by P. tinctorius, T. harzianum and T. viridi. In case of F. oxysporum infection, the bioagents decreased root rot from 47.2% in F. oxysporum treatment to 29.4-35.4% in bioagents, with maximum reduction by L. laccata, followed by P. tinctorius, T. harzianum and T. viridi. The study revealed that, besides improving growth, bioagents especially mycorrhiza protected seedlings against root rot pathogens in blue pine. Inoculation of bioagents in presence of root rot pathogens proved superior over pathogen-inoculated control. Decrease in root rot disease index by antagonistic fungi including mycorrhiza may be attributed to the competition for nutrient base and space, cross protection of roots, alteration in host metabolism and improved nutrient supply. Chakravarty and Adam (1987) reported that growth and development of mycorrhizal species was strongly suppressed by root rot pathogens F. oxysporum and R. solani. Various bioagents reportedly have greater rhizosphere competence and parasitizes the pathogenic fungi (Naik, 2003; Gao et al., 2010). The antagonists in rhizosphere likely compete with the pathogen for host surface and nutrients and inhibit pathogenic growth through antibiosis/mycoparasitism mechanisms (Howell, 2003; Sharma et al., 2010). This seems to have reduced the seedling root decay. These speculations are substantiated by our in vitro bicontrol studies on F. oxysporum and R. solani. The principal mechanisms of Trichoderma spp. for disease control have been presumed to be those primarily acting upon the pathogens and include mycoparasitism, antibiosis and competition for resources and space (Harman, 2006). Thus reduction of root rot in blue pine seedlings incited by F. oxysporum and R. solani may be due to the biological control action of T. harzianum on these pathogens.
Pisolithus tinctorius alone showed 44.2% root colonization in comparison to 39.1% root colonization observed in L. laccatta after 180 days growth. In presence of R. solani the mycorrhizal root colonization was comparatively less, 24.2% in P. tinctorius and 19.5% in L. laccata. Almost similar effect of mycorrhiza was noticed in presence of F. oxysporum. Increased root colonization may be attributed to the competition for space and reduction in root biomass due to root decay by rot causing incitant, thereby leaving less surviving root area for symbiosis. The seedlings with considerable ectomycorrhizal colonization rapidly regenerate new lateral roots, create more new sites for ectomycorrhizae and thereby utilize available nutrients more efficiently than non-mycorrhizal seedlings. Antagonists or mycorrhizae inoculation in Rhizoctonia infected plants lessened the effect of root rot. Mycorrhizal root colonization was less in presence of Rhizoctonia solani and F. oxysporum. This indicates that the disease protection by ectomycorrhiza may involve multiple mechanisms including antibiosis, synthesis of fungistatic compounds by plant roots in response to mycorrhizal infection and physical barrier of fungal mantle around the plant root (Duchesne et al., 1987; Dar et al., 2007). The favourable effect of mycorrhizal fungus on plant growth and health may be attributed to the excretion of growth promoting substances by mycorrhizae or indirectly by alteration in root physiology, uptake of minerals and pattern of exudation into the mycorrhizosphere (Leyval and Berthelin, 1990).
CONCLUSION
The tested fungus biocontrol agents, including mycorrhiza, effectively mitigated root rot disease in blue pine. The bioagents once applied in field multiply in soil and when threshold population is achieved they may reduce the disease incidence thereby ensure the successful establishment of pine seedlings at early stages. The fungal bioagents can be efficiently exploited in integrated disease management module.
REFERENCES
- Ahangar, M.A. G.H. Dar, Z.A. Bhat and N.R. Sofi, 2011. Fungi associated with root rot of Pinus wallichiana seedlings in Kashmir. Plant Pathol. J., 10: 42-45.
CrossRefDirect Link - Altomare, C., W.A. Norvell, T. Bjbrkman and G.E. Harman, 1999. Solubilization of phosphates and micronutrients by the plant-growth-promoting and biocontrol fungus Trichoderma harzianum Rifai 1295-22. Applied Environ. Microbiol., 65: 2926-2933.
Direct Link - Asiegbu, F.O., M. Kacprzak, G. Daniel, M. Johansson, J. Stenlid and M. Manka, 1999. Biochemical interactions of conifer seedling roots with Fusarium spp. Can. J. Microbiol., 45: 923-935.
Direct Link - Beig, M.A., G.H. Dar, N.A. Ganai and N.A. Khan, 2008. Mycorrhizal biodiversity in Kashmir forests and some new records of macrofungi from J&K State. Applied Biol. Res., 10: 26-30.
Direct Link - Benitez, T., A.M. Rincon, M.C. Limon and A.C. Codon, 2004. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol., 7: 249-260.
PubMedDirect Link - Chakravarty, C., R.L. Peterson and B.E. Ellis, 1991. Interaction between the ectomycorrhizal fungus Paxillus involutus, damping-off fungi and Pinus resinosa seedlings. J. Phytopathol., 32: 207-218.
CrossRef - Chakravarty, P. and T. Adam, 1987. Differential influence of ectomycorrhizae on plant growth and disease resistance in Pinus sylvestris seedlings. J. Phytopathol., 120: 104-120.
CrossRef - Choi, D.S., A.M. Quoreshi, V. Maruyama, H.O. Jin and T. Koike, 2005. Effect of ectomycorrhizal infection on growth and photosynthetic characteristics of Pinus densiflora seedlings grown under elevated CO2 concentrations. Photosynthetica, 43: 222-229.
Direct Link - Dar, G.H., M.A. Beig and N.A. Ganai, 2009. Diversity of ectomycorrhizal fungi of various forests in district Baramulla (Jammu and Kashmir). Indian J. For., 32: 137-140.
Direct Link - Dar, G.H., M.A. Beig and N.A. Ganai, 2007. Effect of source and inoculum load of ectomycorrhizae on the growth and biomass of containerized kail pine (Pinus wallichiana) seedlings. Applied Biol. Res., 9: 19-28.
Direct Link - Daughtridge, A.T., R.B. Steven, G.P. Stephen and E.G. Harold, 1986. A rapid staining technique for assessment of ectomycorrhizal infection of oak roots. Can. J. Bot., 64: 1101-1103.
CrossRefDirect Link - Bisht, D., A. Pandey and L.M.S. Palni, 2003. Influence of microbial inoculations on Cedrus deodara in relation to survival, growth promotion and nutrient uptake of seedlings and general soil microflora. J. Sustain. For., 17: 37-54.
CrossRefDirect Link - Dennis, C. and J. Webster, 1971. Antagonistic properties of species-groups of Trichoderma: I. Production of non-volatile antibiotics. Trans. Br. Mycol. Soc., 57: 25-39.
CrossRefDirect Link - Sinclair, J.B. and O.D. Dhingra, 1985. Basic Plant Pathology Methods. CRC Press, Boca Raton, Florida, United States, ISBN: 9780849359217, Pages: 355.
Direct Link - Dunabeitia, M.K., S. Hormilla, I. Jose, K. Txarterina, U. Arteche and J.M. Becerril, 2004. Differential responses of three fungal species to environmental factors and their role in the mycorrhization of Pinus radiata D. Don. Mycorrhiza, 14: 11-18.
CrossRef - Enebak, S.A., M.A. Palmer and R.A. Blanchette, 1990. Managing soil-borne pathogens of white pine in a forest nursery. Plant Dis., 74: 195-198.
Direct Link - Eziashi, E.L., N.U. Uma, A.A. Adekunle and C.E. Airede, 2006. Effect of metabolites produced by Trichoderma species against Ceratocystis paradoxa in culture medium. Afr. J. Biotechnol., 5: 703-706.
Direct Link - Gao, F.K., C.C. Dai and X.Z. Liu, 2010. Mechanisms of fungal endophytes in plant protection against pathogens. Afr. J. Microbiol. Res., 4: 1346-1351.
Direct Link - Gacitua, A.S., F.C. Valiente, P.K. Diaz, C.J. Hernandez, M.M. Uribe, V.E. Sanfuentes, 2009. Identification and biological characterization of isolates with activity inhibitive against Macrophomina phaseolina (Tassi) Goid. Chilean J. Agric. Res., 69: 526-533.
Direct Link - Harman, G.E., 2006. Overview of mechanisms and uses of Trichoderma spp. Phytopathology, 96: 190-194.
CrossRefDirect Link - Howell, C.R., 2003. Mechanism employed by Trichoderma species in the biological control of plant diseases. The history and evolution of current concepts. Plant Dis., 87: 4-10.
Direct Link - Huang, J.W. and E.G. Kuhlman, 1990. Fungi associated with damping off of slash pine seedlings in Georgia. Plant Dis., 74: 27-30.
Direct Link - Heydari, A. and M. Pessarakli, 2010. A review on biological control of fungal plant pathogens using microbial antagonists. J. Biol. Sci., 10: 273-290.
CrossRefDirect Link - Jha, B.N., G.D. Sharma and A.K. Shukla, 2008. Effect of ectomycorrhizal development on growth in pine seedlings. J. Plant Sci., 3: 77-84.
CrossRefDirect Link - Leyval, C. and J. Berthelin, 1990. Influence of acid producing Agrobacterium and Laccaria laccata on pine and beech growth, nutrient uptake and exudation. Agric. Ecosyst. Environ., 28: 313-319.
CrossRef - Lilja, A., M. Poteri, R.L. Petaisto, R. Rikala, T. Kurkela and R. Kasanen, 2010. Fungal diseases in forest nurseries in Finland. Silva Fennica, 44: 525-545.
Direct Link - Lilja, A. and R. Rikala, 2000. Effect of uninucleate Rhizoctonia on the survival of outplanted Scots pine and Norway spruce seedlings. For. Pathol., 30: 109-116.
CrossRef - Lilja, A., A.M. Hallaksela and R. Heinonen, 1995. Fungi colonizing Scots-pine cone scales and seeds and their pathogenicity. Eur. J. For. Pathol., 25: 38-46.
CrossRef - Lu, C. and B. Huang, 2010. Isolation and characterization of Azotobacteria from pine rhizosphere. Afr. J. Microbiol. Res., 4: 1299-1306.
Direct Link - Pinto, P.M., J.A.P. Alonso, V.P. Fernandez and J.J.D. Casero, 2006. Fungi isolated from diseased nursery seedlings in Spain. New Forests, 31: 41-56.
CrossRef - Purkayastha, R.P., U. Menon and B.N. Chakraborty, 1981. Rhizobium-macrophomina interaction affecting phytoalexin production and disease resistance of soybean. Indian J. Exp. Biol., 19: 462-465.
Direct Link - Rangeshwaran, R. and R.D. Prasad, 2000. Isolation and evaluation of rhizospheric bacteria for biological control of chickpea wilt pathogens. J. Biol. Contamin., 14: 9-15.
Direct Link - Reddy, M.S. and K. Natarajan, 1997. Coinoculation efficacy of ectomycorrhizal fungi on Pinus patula seedlings in a nursery. Mycorrhiza, 7: 133-138.
CrossRefDirect Link - Reglinski, T., N. Rodenburg, J.T. Taylor, G.L. Northcott, A.A. Chee, T.M. Spiers and R.A. Hill, 2011. Trichoderma atroviride promotes growth and enhances systemic resistance to Diplodia pinea in radiata pine (Pinus radiata) seedlings. For. Pathol., (In Press).
CrossRef - Sharma, R., R.C. Rajak and A.K. Pandey, 2010. Evidence of antagonistic interactions between rhizosphere and mycorrhizal fungi associated with Dendrocalamus strictus (Bamboo). J. Yeast Fungal Res., 1: 112-117.
Direct Link - Stepniewska-Jarosz, S., M. Manka and F.O. Aslegbu, 2006. Studies on anaestomosis groups of Rhizoctonia solani isolates causing disease in two forest nurseries in Poland. Forest Pathol., 35: 97-109.
CrossRef - Susi, P., G. Aktuganov, J. Himanen and T. Korpela, 2011. Biological control of wood decay against fungal infection. J. Environ. Mgmt., (In Press).
CrossRef - Tabatabai, M.A. and J.M. Bremner, 1969. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem., 1: 301-307.
CrossRefDirect Link - Taylor, A.F.S. and I. Alexander, 2005. The ectomycorrhizal symbiosis: Life in the real world. Mycologist, 19: 102-112.
CrossRef - Villeneuve, N., Le, F. Tacon and D. Bouchard, 1991. Survival of inoculated Laccaria bicolour in competition with native ectomycorrhizal fungi and effects on the growth of outplanted Douglas-fir seedlings. Plant Soil, 135: 95-107.
CrossRef - Vincent, J.M., 1947. Distortion of fungal hyphae in the presence of certain inhibitors. Nature, 159: 850-850.
CrossRefPubMedDirect Link - Haggag, W.M., 2002. Sustainable agriculture management of plant diseases. J. Biological Sci., 2: 280-284.
CrossRefDirect Link - Werner, A., M. Zadworny and K. Ldzikowska, 2002. Interaction between Laccaria laccata and Trichoderma virens in co-culture and the rhizosphere of Pinus sylvestris grown in vitro. Mycorrhiza, 12: 41-47.
CrossRef - Widyastuti, S.M., S. Harjono and D. Yuniarti, 2003. Biological control of sclerotium rolfsii damping-off of tropical pine (Pinus merkusii) with three isolates of Trichoderma spp. J. Biological Sci., 3: 95-102.
CrossRefDirect Link - Windham, M.T., Y. Elad and R. Baker, 1986. A mechanism for increased plant growth induced by Trichoderma spp. Phytopathology, 76: 518-521.
Direct Link - Yamada, A., T. Ogura and M. Ohmasa, 2001. Cultivation of mushrooms of edible ectomycorrhizal fungi associated with Pinus densiflora by in vitro mycorrhizal synthesis-II, morphology of mycorrhizas in open pot soil. Mycorrhiza, 11: 59-66.
CrossRefDirect Link - Yamaji, K., H. Ishimoto, N. Usui and S. Mori, 2005. Organic acids and water soluble phenolics produced by Paxillus sp. together show antifungal activity against Pythium vexans under acidic culture conditions. Mycorrhiza, 15: 17-23.
CrossRef - Yobo, K.S., M.D. Laing and C.H. Hunter, 2010. Application of selected biological control agents in conjunction with tolclofos-methyl for the control of damping-off caused by Rhizoctonia solani. Afr. J. Biotech., 9: 1789-1796.
Direct Link - Farquhar, M.L. and R.L. Peterson, 1990. Early effects of the ectomycorrhizal fungus Paxillus involutus on the root rot organism Fusarium associated with Pinus resinosa. Can. J. Bot., 68: 1589-1596.
Direct Link