
Thnaian Althnaian
Department of Anatomy, College of Veterinary Medicine and Animal Resources, King Faisal University, Al-Ahsa, Saudi Arabia
Journal of Biological Sciences
Year: 2016 | Volume: 16 | Issue: 3 | Page No.: 65-71







ABSTRACT
Because of AFB1 effects on antioxidant enzyme activities and gene expression in kidneys have not been completely elucidated, the current study was designed to investigate the histopathological picture and activities and gene expression of antioxidant enzymes in kidneys of rats. Therefore, 20 rats were randomly divided into two equal groups. Rats in the first group served as control whereas rats in the second group received single i.p., dose of AFB1 (3 mg kg–1 b. wt.). Kidney damage and oxidative stress were observed in AFB1-intoxicated rats as indicated by a significant elevation in thiobarbituric acid reactive substances (TBARS), reduction of reduced glutathione (GSH) concentration, reduction in the activities of antioxidant enzymes namely catalase (CAT), total superoxide dismutase (SOD), glutathione peroxidase (GPX) and glutathione-s-transferase (GST) and down-regulation of gene expression of these antioxidant enzymes compare to control. Kidney sections of rats intoxicated with AFB1 showed swelling and vacuolar cytoplasm of distal tubules and desquamated distal tubular cells. In conclusion, AFB1 may exert its toxicity to kidney tissues by induction of lipid peroxidation and inactivation and down-regulation of gene expression of antioxidant enzymes.
PDF Abstract XML References Citation
Received: December 04, 2015;
Accepted: January 21, 2016;
Published: March 15, 2016
How to cite this article
Thnaian Althnaian, 2016. Gene Expression and Activities of Antioxidant Enzymes in Kidneys of Rats Intoxicated with Aflatoxin B1. Journal of Biological Sciences, 16: 65-71.
DOI: 10.3923/jbs.2016.65.71
URL: https://scialert.net/abstract/?doi=jbs.2016.65.71
DOI: 10.3923/jbs.2016.65.71
URL: https://scialert.net/abstract/?doi=jbs.2016.65.71
INTRODUCTION
Kidney has an important role in excretions of drugs and body waste products by highly specialized cells Choi et al. (2011). Fungal contaminated food may affect the urinary system. Fungal metabolism (Aspergillus flavus and Aspergillus parasiticus) produces aflatoxins (Goto et al., 1996). The AFB1 is the most toxic type (Busby and Wogan, 1984) in human and animals (Roebuck and Maxuitenko, 1994). The concentration of the absorbed AFB1 was higher in gonads, liver, kidneys, spleen, thymus, endocrine gland, lung and brain (Marvan et al., 1983). Cytochrome P450 (CYP450) acts on AFB1 and produces AFB1-8, 9-epoxide (Smela et al., 2001) which is responsible for induction of oxidative stress of tissues (Antunes et al., 2002; Atessahin et al., 2003; El-Demerdash et al., 2004; El-Bahr, 2015), depletion of antioxidants (El-Bahr, 2015), formation of DNA adducts and tumor initiation (Choy, 1993). The histopathological studies on kidney tissues indicated hydropic and vacuolar degeneration, parenchymatous tubular degeneration of the distal tubules, epithelial swelling, desquamation of epithelial tubular cells and granular appearance of cytoplasm in aflatoxin treated rats (Devendran and Balasubramanian, 2011). In addition, kidney necrosis and increased ratio of the area of bowman’s capsules to the area of its glomerulus have been reported earlier (Al-Habib et al., 2007). Recently, new assumption of the mechanism of action of AFB1 has been reported (El-Bahr, 2015). This assumption stated that, toxicity of AFB1 may due to significant down-regulation of gene expression of hepatic antioxidant enzymes in rats intoxicated with AFB1 (El-Bahr, 2015). However, this assumption has not been verified in kidney tissues of AFB1 intoxicated rats. Therefore, the current study aimed to investigate the effect of AFB1 on the activities and regulation of gene expression of antioxidant enzymes in kidney of rats.
MATERIALS AND METHODS
Chemicals: The AFB1, Ethylene Glycol Tetraacetic Acid (EGTA), ethylenediaminetetraacetic acid (EDTA), sucrose, tris, butanol, mannitol, metaphosphoric acid, hydrogen peroxide (H2O2) and dimethyl sulfoxide (DMSO) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Radioimmunoprecipitation assay (RIPA) buffer was provided by Cayman Chemical Company, USA. All other chemicals were of analytical grade.
Animals, experimental design and sampling: Twenty rats (100±5 g) were obtained from the laboratory animal house of the College of Veterinary Medicine and Animal Resources, King Faisal University, Saudi Arabia. They were maintained according to the national guidelines and protocols, which were approved by the University Animal Ethics Committee. They were housed in clean and disinfected plastic cages. Commercial basal diet and water were provided ad libitum. Rats were subjected to a natural photoperiod of 12 h light: Dark cycle throughout the experimental period which lasted for 5 weeks (35 days). The rats were divided into two groups (10 rats per group). Rats in the first group were injected with a single i.p., dose of 1% DMSO at the first day of the experiment and served as control whereas, rats in the second group were administered with a single i.p., dose of AFB1 (3 mg kg–1 b.wt.) (Kamdem et al., 1983) dissolved in 1% DMSO at the first day of the experiment. After 5 weeks (day 35), the rats were anaesthetised with diethyl ether. The kidney tissues were removed and divided into three portions. The first portion of the kidney tissue was cut into small pieces and immersed in neutral-buffered formalin for 24 h for histopathological examination. The second portion of the kidney tissue was immediately frozen at -30°C for biochemical analysis of antioxidant enzyme activities and the last portion of the kidney tissue was immediately frozen in liquid nitrogen and stored at -80°C for molecular analysis of antioxidant enzyme gene expression.
Histopathological assessment of kidney damage: Kidney tissues were cut into small pieces and immersed in neutral-buffered formalin for 24 h. The fixed tissues were processed routinely, embedded in paraffin, sectioned, deparaffinised and rehydrated using standard techniques (Bancroft and Gamble, 2002). The extent of AFB1-induced necrosis was evaluated by assessing the morphological changes in the kidney sections stained with hematoxylin and eosin (H and E) using standard techniques.
Determination of kidney antioxidant enzymes, thiobarbituric acid reactive substances (TBARS) and reduced glutathione (GSH) levels: One gram of kidney tissue was homogenised in 5 mL of cold 20 mM HEPES buffer, pH 7.2, containing 1 mM EGTA, 210 mM mannitol and 70 mM sucrose. After centrifugation )1500 g, 5 min) at 4°C, the supernatant was removed and stored frozen at -80°C until further analysis of total superoxide dismutase (SOD). One gram of kidney tissue was homogenised in 5 mL of cold 50 mM potassium phosphate buffer, pH 7, containing 1 mM EDTA. After centrifugation (10,000 g, 15 min) at 4°C, the supernatant was removed and stored frozen at -80°C until further analysis of catalase (CAT), glutathione peroxidase (GPX), glutathione-s transferase (GST) and reduced glutathione (GSH). The extent of lipid peroxidation in terms of TBARS formation was measured by mixing 1 g of kidney tissue with RIPA buffer (catalogue #10010263, Cayman Chemical Company, USA). After homogenisation, sonication and centrifugation (1600 g, 10 min), the supernatant was removed and stored frozen at -80°C until further analysis. The ELISA kits of Cayman Chemical Company, USA were used for determination of the activities of CAT (nmol/min/g tissue, catalogue #707002), GPX (nmol/min/g tissue, catalogue #703102), total SOD (U/g tissue, catalogue #706002) and GST (nmol/min/g tissue, catalogue #703302) and the concentrations of GSH (μM, catalogue #703002) and TBARS (μM, catalogue #10009055) by using an ELISA reader (Absorbance Microplate Reader ELx 800TM BioTek®, USA). The results were calculated according to the manufacturer’s instruction.
Total RNA isolation and real-time RT-PCR of kidney antioxidant enzymes: Kidney tissues (approximately 1 g of tissue per sample) were immediately added to 1 mL of TRIzol reagent (Invitrogen, Carlsbad, CA) and homogenised using a homogeniser (Tissue Ruptor, Qiagen GmbH, Germany). One millilitre of the tissue homogenate was transferred to a microfuge tube and total RNA was extracted by the addition of 0.2 mL chloroform. Next, the samples were vigorously vortexed for 15 sec and incubated at room temperature for 3 min. After centrifugation )12,000 g, 15 min) at 4°C, the aqueous phase containing RNA was transferred to new tubes. The RNA was precipitated by mixing the aqueous phase with 0.5 mL isopropyl alcohol and incubated at room temperature for 10 min. After centrifugation at 12,000 g for 10 min at 4°C, RNA pellets were washed by mixing and vortexing with 1 mL of 75% ethanol. After centrifugation (7,500 g, 5 min) at 4°C, RNA pellets were resuspended in nuclease-free water (Life Technologies. USA). The purity of RNA at 260/280 OD ratio and the RNA integrity were evaluated using a Multi-Mode Microplate reader (SYNERGY Mx, BIO-TEK. Winooski, Vermont, USA). Only high purity samples (OD260/280 >1.8) were subjected to further manipulation. cDNA was obtained from RNA samples using a Reverse Transcription System Kit (Promega, Madison, USA) and Bio-Rad Thermal Cycler (T100TM, Foster city, California, USA). Briefly, total RNA was activated at 70°C for 10 min and 20 μL reaction mixtures were prepared with 4 μL MgCl2, 2 μL of reverse transcription 10x letter buffer, 2 μL of dNTP mixture (10 mM), 0.5 μL of random primers, 0.75 μL of AMV reverse transcriptase enzyme, 1 ng RNA and nuclease-free water to a final volume of 20 μL. Next, the reaction was incubated at 42°C for 60 min, followed by incubation at 94°C for 5 min. The cDNA was diluted up to 100 μL with nuclease-free water for PCR amplification. Real-time RT-PCR was performed using the QuantiFastTM SYBR Green PCR Master Mix Kit (QIAGEN, Hilden; Germany). The 25 μL reaction for each examined gene was prepared from 12.5 μL of master mix, 2 μL forward primer (10 pmol), 2 μL reverse primer (10 pmol), 2 μL cDNA of the sample and 6.5 μL of nuclease-free water. The cycling parameters were 50°C for 2 min, 95°C for 15 min, 40 cycles of 95°C for 10 sec, followed by 55°C for 30 sec and 72°C for 10 sec with a final melting temperature at 95°C for 20 sec. For each gene examined, duplicate samples from each cDNA were analysed using real-time RT-PCR and the Bio-Rad CFX Manager 3.0 Software of the C1000 Touch thermal cycler-CFX96 Real-time PCR (BIO-RAD, Foster city, California, USA). The β-actin mRNA fragment was used as a housekeeping gene to normalise the expression data. The primer sequences are described in Table 1.
Statistical analysis: All data are presented as the mean±standard error of the mean using one way analysis of variance (ANOVA). All tests were performed using a statistical analysis system program (SAS., 2002). The relative gene expression of target genes compared to the β-actin reference gene was calculated using the Bio-Rad CFX Manager 3.0 Software of the C1000 Touch thermal cycler-CFX96 Real-time PCR (BIO-RAD, Foster City, California, USA).
RESULTS
Morphological and histopathological assessment of kidney damage: No morphological gross changes were detected in kidney of the control group. However, kidneys of the rats intoxicated with AFB1 showed congestion, hemorrhage, swelling and yellowish-white and depressed areas.
| Table 1: | Details giving primer sequences for the genes amplified |
 | |
| SOD: Superoxide dismutase, CAT: Catalase, GPX: Glutathione peroxidase, GST: Glutathione-s-transferase | |
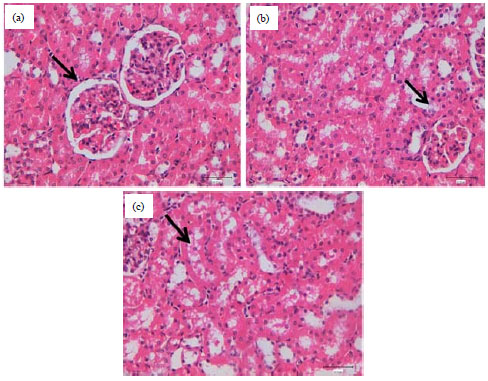 | |
| Fig. 1(a-c): | Histopathological assessment of renal damage in (a) Control kidney showing normal kidney surrounding by proximal and distal tubules (arrow), HE. Bar = 40.00 μm, (b) Kidney of AFB1 treated rats showing swelling and vacuolar cytoplasm of distal tubules (arrows), HE. Bar = 40.00 μm and (c) Kidney of AFB1 treated rats showing desquamated distal tubular cells (arrow), HE. Bar = 40.00 μm |
| Table 2: | Effect of AFB1 for five weeks on levels of TBARS (μM) and GSH (μM) and activities of CAT (nmol/min/g tissue), total SOD (U g–1 tissue), GPX (nmol/min/g tissue) and GST (nmol/min/g tissue) in rats kidneys tissues |
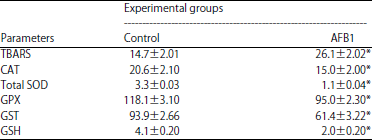 | |
Values are expressed as Mean±SEM, n = 10 for each group, significance was calculated at p<0.05, TBARS: Thiobarbituric acid reactive substances, CAT: Catalase, SOD: Superoxide dismutase, GPX: Glutathione peroxidase, GST: Glutathione-s-transferase, GSH: Reduced glutathione, *Significant as compared to control animals, **Significant as compared to AFB1 treated animals | |
Histopathological assessment of kidney damage showed in Fig. 1. Kidney of control rats showed normal tissues surrounding by proximal and distal tubules (Fig. 1a) whereas, kidney of AFB1 treated rats showed swelling and vacuolar cytoplasm of distal tubules (Fig. 1b) and desquamated distal tubular cells (Fig. 1c).
Effect of treatments on testicular lipid peroxidation: Data summarized in Table 2 showed that, the TBARS level was significantly (p<0.05) increased in the kidney tissues of AFB1-intoxicated rats compare to the control.
Effect of treatments on GSH and testicular antioxidant enzyme activities: The GSH concentration and antioxidant enzyme activities (CAT, total SOD, GPX and GST) are shown in Table 2. The concentration of GSH was significantly reduced (p<0.05) in kidney tissues of AFB1-intoxicated rats compare to the control. All of the antioxidant enzyme activities were significantly reduced (p<0.05) in kidney tissues of AFB1-intoxicated rats compare to the control.
Effect of treatments on the gene expression of antioxidant enzymes: The mRNA expression of total SOD, CAT, GPX and GST was measured using real-time RT-PCR (Fig. 2). These results showed that, the expression of all of the antioxidant enzymes was significantly reduced (p<0.05) in kidney tissues of AFB1-intoxicated rats compare to the control.
DISCUSSION
The morphological changes observed in the current study agree with previous report in rats (Al-Ghasham et al., 2008). The histopathological findings reported in the current study were parallel to the current biochemical results and indicated degenerative changes in the kidney of rats intoxicated with AFB1. Similar histopathological changes of aflatoxin-induced kidney damage in rats have been previously reported (Al-Habib et al., 2007; Devendran and Balasubramanian, 2011; Fetaih et al., 2015; El-Mahalaway, 2015).
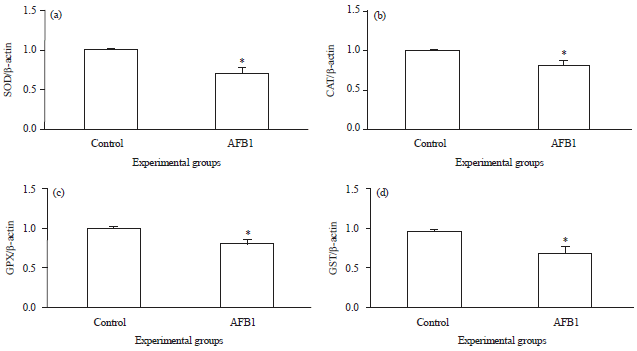 | |
| Fig. 2(a-d): | Real time RT-PCR analysis of, (a) SOD, (b) CAT, (c) GPX and (d) GST in kidneys tissues of control and aflatoxin B1 (AFB1) treated rats. Values are expressed as Mean±SEM, *Values are significantly different (p<0.5) compared to control |
The significant increase in TBARS in the kidney tissues of AFB1-intoxicated rats indicated oxidative stress. This finding was consistent with previous studies in the kidney of rats (Mohamed and Metwally, 2009; Abdulmajeed, 2011; El-Mahalaway, 2015; Hassan et al., 2015) and liver (El-Bahr, 2015; El-Bahr et al., 2015) intoxicated with AFB1. The significant increase in kidney TBARS in AFB1-intoxicated rats was potentially attributed to the AFB1 metabolised by the cellular cytochrome P450 enzyme system to form the reactive intermediate, AFB1-8, 9-epoxide, which in turn reacted with biomolecules such as lipids, resulting in lipid peroxidation and cellular damage (Stresser et al., 1994). In addition, the current study reported a significant decrease in both the GSH content and the activities of antioxidant enzymes (total SOD, GPX, CAT and GST) in AFB1-intoxicated rats compared to the control, which induced an elevation of TBARS levels.
Glutathione plays an important role in the antioxidant system of the body. It maintains the normal structure and function of the cells via a redox and detoxification reaction. In the present study, a significant decrease in GSH values was observed in kidney tissues of AFB1-intoxicated rats. The depletion of hepatic GSH in these animals may be attributed to the conjugation of GSH with electrophilic metabolites of AFB1, a reaction that is catalysed by GST. Similar results were reported in kidney (Mohamed and Metwally, 2009; Abdulmajeed, 2011; Devendran and Balasubramanian, 2011; Deabes et al., 2012; Hassan et al., 2015) and liver tissues (El-Bahr, 2015; El-Bahr et al., 2015) of rats intoxicated with AFB1. Oxidative stress is defined as the imbalance between pro-oxidants and antioxidants in biological systems. Thus, the significant increase in lipid peroxidation could be due to a significant reduction in the activities of enzymatic antioxidants, such as CAT, total SOD, GPX and GST as well as non-enzymatic antioxidants such as GSH, in the kidney tissues of AFB1-intoxicated rats compared to the control. Superoxide radicals are converted to H2O2 by SOD. Furthermore, H2O2 is converted to molecular oxygen and H2O by either CAT or GPX. Moreover, GPX can reduce lipid peroxides and other organic hydroperoxides that are highly cytotoxic products. Thus, SOD, CAT, GPX and GST constitute the principal components of the antioxidant system and their deficiencies result in oxidative stress (Ahmad et al ., 2012). Moreover, a significant reduction in these enzyme activities in the kidneys of AFB1-intoxicated rats could be responsible for the observed increased lipid peroxidation as reflected in the high level of TBARS combined with the low level of GSH in these animals compared to the control during aflatoxicosis. Significant reductions in GPX, SOD, CAT and GST activities have been reported in the kidneys of aflatoxin-fed rats (Deabes et al ., 2012; Antonyak et al ., 2015) and mice (Gupta and Sharma, 2011). In addition, the same significant reductions in GPX, SOD, CAT and GST activities have been reported in the livers of aflatoxin-fed rats (El-Bahr, 2015; El-Bahr et al ., 2015). At the gene expression level, the current findings indicated that AFB1 induced a down-regulation of all antioxidant enzyme gene expression in AFB1-intoxicated rats. The publications regarding the gene expression levels of antioxidant enzymes in kidneys of rats intoxicated with AFB1 are lack. To the best of the author knowledge, the current study may the first study to address the expression of these enzymes in kidneys of rats intoxicated with AFB1. However, the gene expression of the antioxidants enzymes in liver of rats intoxicated with AFB1 has been documented. In this study, Yarru et al. (2009) reported that dietary aflatoxin induced a down-regulation of SOD gene expression in livers of broiler chicks fed with aflatoxin. This down-regulation caused an accumulation of superoxide anions within the mitochondria and impaired its proper function (Marczuk-Krynicka et al., 2003). Additionally, the most recent report (El-Bahr, 2015) indicated down-regulation of gene expression of CAT, GST, GPX and SOD in livers of rats intoxicated with AFB1. The conjugation of reactive xenobiotic metabolites with GSH as previously described above is an important step in the detoxification mechanism. This conjugation process is mediated by GST. An overload of xenobiotics (AFB1) depleted the GSH via the conjugation process. Down-regulation of GST gene expression in AFB1-intoxicated rats compared to the control as demonstrated in the current study (Fig. 2c) could limit the ability of hepatic tissues to conjugate the reactive metabolites.
CONCLUSION
The present study concluded that, AFB1-induced kidney damage in rats. AFB1 may exert its toxicity to kidney tissues by induction of lipid peroxidation and inactivation and down-regulation of gene expression of antioxidant enzymes.
ACKNOWLEDGMENT
The author thanks the Deanship of Scientific Research in King Faisal University, Saudi Arabia for their support of this study.
REFERENCES
- Abdulmajeed, N.A., 2011. Therapeutic ability of some plant extracts on aflatoxin B1 induced renal and cardiac damage. Arabian J. Chem., 4: 1-10.
CrossRefDirect Link - Ahmad, H., J. Tian, J. Wang, M.A. Khan, Y. Wang, L. Zhang and T. Wang, 2012. Effects of dietary sodium selenite and selenium yeast on antioxidant enzyme activities and oxidative stability of chicken breast meat. J. Agric. Food Chem., 60: 7111-7120.
CrossRefDirect Link - Al-Ghasham, A., H.S. Ata, S. El-Deep, A.R. Meki and S. Shehada, 2008. Study of protective effect of date and Nigella sativa on Aflatoxin B1 toxicity. Int. J. Health Sci., 2: 26-44.
PubMedDirect Link - Al-Habib, M.F.M., A.A. Jaffar and H.H. Abdul-Ameer, 2007. Aflatoxin B1-induced kidney damage in rats. J. Fac. Med. Baghdad, 49: 147-150.
Direct Link - Antonyak, H., C. Olijnyk, N. Koval, R. Fedyakov, M. Dosviadchynska, I. Panchuk and B. Chekh, 2015. Effects of aflatoxin B1 on lipid peroxidation and activities of antioxidant enzymes in rat organs and erythrocytes. Visnyk Lviv Univ.: Ser. Biol., 69: 41-48.
Direct Link - Antunes, N.L., M.M. Souweidane, E. Li, M.K. Rosenblum and P.G. Steinherz, 2002. Methotrexate leukoencephalopathy presenting as Kluver-Bucy syndrome and uncinate seizures. Pediatr. Neurol., 26: 305-308.
CrossRefPubMedDirect Link - Atessahin, A., I. Karahan, S. Yilmaz, A.O. Ceribasi and I. Princci, 2003. The effect of manganese chloride on gentamicin-induced nephrotoxicity in rats. Pharmacol. Res., 48: 637-642.
CrossRefPubMedDirect Link - Choi, J.J., B.S. Moffett, E.J. McDade and D.L. Palazzi, 2011. Altered gentamicin serum concentrations in obese pediatric patients. Pediatric Infect. Dis. J., 30: 347-349.
CrossRefDirect Link - Choy, W.N., 1993. A review of the dose-response induction of DNA adducts by aflatoxin B1 and its implications to quantitative cancer-risk assessment. Mutat. Res./Rev. Genet. Toxicol., 296: 181-198.
CrossRefDirect Link - Deabes, M.M., H.R. Darwish, K.B. Abdel-Aziz, I.M. Farag, S.A. Nada and N.S. Tawfek, 2012. Protective effects of Lactobacillus rhamnosus GG on aflatoxins-induced toxicities in male albino mice. Environ. Anal. Toxicol., 2: 132-140.
CrossRefDirect Link - Devendran, G. and U. Balasubramanian, 2011. Biochemical and histopathological analysis of aflatoxin induced toxicity in liver and kidney of rat. Asian J. Plant Sci. Res., 1: 61-69.
Direct Link - El‐Bahr, S.M., 2015. Effect of curcumin on hepatic antioxidant enzymes activities and gene expressions in rats intoxicated with aflatoxin B1. Phytother. Res., 29: 134-140.
CrossRefDirect Link - El-Bahr, S.M., M.A. Embaby, A.A. Al-Azraqi, A.M. Abdelghany, Y.A. Hussein, F.A. Hizab and T.A. Althnaian, 2015. Effect of curcumin on aflatoxin B1-induced toxicity in rats: A biochemical and histopathological study. Int. J. Biochem. Res. Rev., 5: 63-72.
CrossRefDirect Link - El-Demerdash, F.M., M.I. Yousef, F.S. Kedwany and H.H. Baghdadi, 2004. Cadmium-induced changes in lipid peroxidation, blood hematology, biochemical parameters and semen quality of male rats: Protective role of vitamin E and β-carotene. Food Chem. Toxicol., 42: 1563-1571.
CrossRefPubMedDirect Link - El-Mahalaway, A.M., 2015. Protective effect of curcumin against experimentally induced aflatoxicosis on the renal cortex of adult male albino rats: A histological and immunohisochemical study. Int. J. Clin. Exp. Pathol., 8: 6019-6030.
Direct Link - Fetaih, H.A., A.A. Dessouki and A.S. Tahan, 2015. Toxopathological effect of AFB1 on pregnant rats. Global Anim. Sci. J., 3: 145-153.
Direct Link - Goto, T., D.T. Wicklow and Y. Ito, 1996. Aflatoxin and cyclopiazonic acid production by a sclerotium-producing Aspergillus tamarii strain. Applied Environ. Microbiol., 62: 4036-4038.
Direct Link - Gupta, R. and V. Sharma, 2011. Ameliorative effects of Tinospora cordifolia root extract on histopathological and biochemical changes induced by aflatoxin-B1 in mice kidney. Toxicol. Int., 18: 94-98.
CrossRefDirect Link - Hassan, A.M., S.H. Abdel-Aziem, A.A. El-Nekeety and M.A. Abdel-Wahhab, 2015. Panax ginseng extract modulates oxidative stress, DNA fragmentation and up-regulate gene expression in rats sub chronically treated with aflatoxin B1 and fumonisin B1. Cytotechnology, 67: 861-871.
CrossRefDirect Link - Kamdem, L., J. Magdalou, G. Siest, M. Ban and D. Zissu, 1983. Induced hepatotoxicity in female rats by aflatoxin B1 and ethynylestradiol interaction. Toxicol. Applied Pharmacol., 67: 26-40.
CrossRefDirect Link - Marczuk-Krynicka, D., T. Hryniewiecki, J. Piatek and J. Paluszak, 2003. The effect of brief food withdrawal on the level of free radicals and other parameters of oxidative status in the liver. Med. Sci. Monit., 9: 131-135.
PubMedDirect Link - Mohamed, A.M. and N.S. Metwally, 2009. Antiaflatoxigenic activities of some plant aqueous extracts against aflatoxin-b1 induced renal and cardiac damage. J. Pharmacol. Toxicol., 4: 1-16.
CrossRefDirect Link - Smela, M.E., S.S. Currier, E.A. Bailey and J.M. Essigmann, 2001. The chemistry and biology of aflatoxin B1: From mutational spectrometry to carcinogenesis. Carcinogenesis, 22: 535-545.
CrossRefPubMedDirect Link - Stresser, D.M., G.S. Bailey and D.E. Williams, 1994. Indole-3-carbinol and beta-naphthoflavone induction of aflatoxin B1 metabolism and cytochromes P-450 associated with bioactivation and detoxication of aflatoxin B1 in the rat. Drug Metab. Dispos., 22: 383-391.
PubMedDirect Link - Yarru, L.P., R.S. Settivari, N.K.S. Gowda, E. Antoniou, D.R. Ledoux and G.E. Rottinghaus, 2009. Effects of turmeric (Curcuma longa) on the expression of hepatic genes associated with biotransformation, antioxidant and immune systems in broiler chicks fed aflatoxin. Poult. Sci., 88: 2620-2627.
CrossRefPubMedDirect Link