
S. Jayakumar
Radiation Biology and Health Sciences Division, Bhabha Atomic Research Centre, Mumbai 400085, India
Sarvjeet Kaur
Radiation Biology and Health Sciences Division, Bhabha Atomic Research Centre, Mumbai 400085, India
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 6 | Page No.: 463-473







ABSTRACT
Bacillus thuringiensis (Bt) has been used as biopesticide sprays due to its insecticidal specificity but precipitation loss is a limitation. Bt isolates, naturally occurring on crop phylloplanes, have better on-plant persistence. Bt isolates have been isolated and characterized from phylloplanes of leguminous crops. Bt isolates, which showed presence of highly conserved 16S-23S rRNA internal transcribed spacer region, were screened by PCR for cry gene families. The cry1 gene family was found to be most abundant, followed by cry2 gene family, while none of isolates showed presence of cry3, 4, 7 and 8 gene families. Bt isolates were further screened for presence of specific genes of cry1 gene family. Four isolates-SK-222, SK-223, SK-229 and SK-232, were found to have cry1Aa, cry1Ab, cry1Ac and cry1D genes. Isolate SK-222 was found to contain maximum types of genes followed by SK-223, SK-229 and SK-232. Protein profiles of isolates by SDS-PAGE showed 130 kDa band corresponding to cry1 protein. Isolate SK-223 was most toxic followed by SK-222, SK-229 and SK-232 towards diamondback moth (Plutella xylostella) by leaf dip bioassay. Isolates SK-222 and SK-223 were significantly more toxic than Bt subsp. kurstaki (HD1). These isolates have potential of development into biopesticides. Full length cry 1Ab gene was amplified by PCR with specially designed primers from isolate SK-222, cloned and sequenced (GenBank accession No. DQ023297). Sequence analysis showed it to be identical to cry1Ab18 gene. (http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/, GenBank accession No. AY319967) isolated previously from a Bt isolate recovered from soil of Ladakh region in this laboratory.
PDF Abstract XML References Citation
Received: March 15, 2013;
Accepted: March 29, 2013;
Published: July 12, 2013
How to cite this article
S. Jayakumar and Sarvjeet Kaur, 2013. Occurrence of cry Genes in Bacillus thuringiensis (Bt) Isolates Recovered from Phylloplanes of Crops Growing in the New Delhi Region of India and Toxicity Towards Diamond-back Moth (Plutella xylostella). Journal of Biological Sciences, 13: 463-473.
DOI: 10.3923/jbs.2013.463.473
URL: https://scialert.net/abstract/?doi=jbs.2013.463.473
DOI: 10.3923/jbs.2013.463.473
URL: https://scialert.net/abstract/?doi=jbs.2013.463.473
INTRODUCTION
Insect infestation is a major limiting factor in enhancement of agricultural productivity. Excessive use of chemical insecticides has led to emergence of resistance, long residual action and toxicity to a wide spectrum of organisms. Bacillus thuringiensis (Bt) produces cry proteins that have specific insecticidal activity (Anwar et al., 1999, Ali et al., 2010). Bt has been used as a microbial biopesticide because of advantages of specific toxicity against target insects, lack of polluting residues and safety to non-target organisms such as mammals, birds, amphibians and reptiles and low cost of development and registration. It accounts for 95% of the 1% market share of biopesticides in the total pesticide market (Flexner and Belnavis, 1999). However, the use of Bt microbial biopesticide formulations has been rather limited due to the problems of narrow host range, low persistence on plants and inability of foliar application to reach the insects feeding inside the plants, notwithstanding several biotechnological approaches for the development of improved Bt biopesticides (Kaur, 2000).
The problems associated with field application of Bt biopesticides have been overcome by Bt transgenic crops (Khan et al., 2001). However, many major pulse crops of India have proved recalcitrant to genetic transformation (Chandra and Pental, 2003). In addition, the deployment of transgenic food crops requires adequate regulatory controls as well as public acceptance (Shelton et al., 2002, Kaur, 2007).
Bt strains have been isolated worldwide from diverse habitats, including soil (Kaur and Singh 2000a; Wang et al., 2003), stored grains (Meadows et al., 1992) phyllosplanes (Hansen et al., 1998; Kaur and Singh, 2000b; Jara et al., 2006) and other miscellaneous habitats (Uribe et al., 2003; Apaydin et al., 2005; Stobdan et al., 2005; Martinez and Caballero, 2003). Over 700 cry genes grouped into more than 60 families with insecticidal spectrum extending over several invertebrate orders have been isolated from Bt strains worldwide (Crickmore et al., 1998; Beard et al., 2001, Wang et al., 2003; Stobdan et al., 2004; Swiecicka et al., 2008; Van Frankenhuyzen, 2009; Noguera and Ibarra, 2010). The identification of cry genes by Polymerase Chain Reaction (PCR) has proven to be a very useful method for characterization of cry genes present in Bt strains due to its rapidity and reproducibility (Porcar and Juarez-Perez, 2003; Beron et al., 2005; Kaur, 2006).
Native Bt isolates from diverse habitats in India have been isolated and screened for the presence of different cry genes in our laboratory (Kaur and Singh 2000a, b, Stobdan et al., 2005; Shankar et al., 2010; Katara et al., 2012; Meena et al., 2012). Bt isolates, which naturally occur in the phylloplane of crops and have high insecticidal activity towards target insect pests can be developed as effective biopesticides. In addition, native Bt isolates can be screened for the presence of novel cry genes in order to fulfil the requirement of specific cry proteins for specific pests and to overcome the resistance developed by insects against existing cry genes. Therefore, we have characterized Bt isolates, from phylloplane of leguminous plants.
MATERIALS AND METHODS
Materials
Bacterial isolates and strains: BT isolates recovered from leguminous phylloplanes in the corresponding author Dr. S. Kaur’s laboratory were used in this study. Bt strains and the recombinant ECE strain (E. coli clones with cry gene) used as reference in this study were obtained from Dr. D.R. Ziegler, Director, Bacillus Genetic Stock Center, Ohio State University, Columbus, OH, USA.
Growth media for bacterial strains: Luria Bertani Agar (LA) and Luria Bertani Broth (LB) were used for the growth of E. coli and Bt strains.
Methods
Isolation of genomic DNA: Genomic DNA was isolated using method modified from Kalman et al. (1993).
Five milliliter starter culture was added to 50 mL LB medium and incubated with shaking at 200 rpm (at 30°C for 12-16 h). Cells were harvested by centrifuging at 8000 rpm for 10 min (JA 20 rotor, Beckman). The pellet was washed in 5 mL Solution I (100 mM Tris-HCl (pH 8.0), 1 mM EDTA and 100 mM NaCl) and resuspended in 10 mL of Solution II (25 mM Tris-HCl (pH 8.0) 25 mM EDTA, 25% Sucrose and Lysozyme 1 mg mL-1) and then incubated at 37°C for 1 h. Then the cells were solubilized by addition of 10% SDS to final concentration of 2% and incubated at 50°C for 15 min. Thereafter, NaCl was added to a final concentration of 1 M and incubated (50°C for 5 min), centrifuged at 10,000 rpm for 15 min and the supernatant was taken out and precipitated with ethanol. Then the pellet was dissolved in TE buffer (10 mM Tris-HCl and 1 mM EDTA).
Isolation of plasmid DNA: A modified alkaline lysis method (Birnboim and Doly, 1979) was used to isolate the plasmid DNA. A 100 mL of Bt cell culture in LB medium was prepared and the cells were pelleted by centrifugation (at 8000 rpm for 15 min). The pellet was resuspended in 2.5 mL of ice-cold solution I (50 mM glucose; 25 mM Tris-HCl (pH 8.0); 10 mM EDTA and Lysozyme was added just before use @ 10 mg mL-1) and then 5 mL of freshly prepared solution II (1 M NaCl, 1% SDS) was added and mixed gently by inverting 4-6 times and incubated at room temperature for 10 min. Thereafter, 3 mL of ice-cold solution III (5 M potassium acetate, 11.5% glacial acetic acid) was added, mixed gently and incubated on ice for 20 min. The suspension was centrifuged at 12,000 rpm for 25 min and the supernatant was separated, wherefrom the plasmid DNA was purified by the ion exchange column and solutions provided by Qiagen plasmid purification kit (Qiagen midi kit, Germany).
PCR analysis of 16S-23S rDNA spacer region: The primer pair designed as per Jensen et al. (1993) was used for amplification of the highly conserved 16S-23S rDNA Internal Spacer Region (ISR) sequence to characterize Bt isolates. The forward primer GI (5’GAAGTCGTA ACAAGG3’) corresponds to a highly conserved region on 16S immediately upstream to the spacer region and the reverse primer (LI (5’ CAAGGCATCCACCGT 3’) corresponds to 23S region immediately next to the internal spacer region. Reaction mixture consisting of genomic DNA 50 ng; primers GI and LI 1 μM each, MgCl2 (25 mM) 5 μL, dNTPs (2 mM) 5 μL; PCR buffer (10X) 5 μL; 1.0 U of Taq DNA polymerase (MBI Fermentas) and volume made up to 50 μL by sterile distilled water, was amplified in a thermal cycler (Gene Amp). The programme for reaction was in 5 steps: Step 1: 94°C for 2 min; Step 2: 94°C for 1 min; Step 3: 50°C for 1; Step 4: 72°C for 2 min; Step 5: 72°C for 10 min. The steps 2, 3 and 4 were repeated 30 times and the PCR products were analyzed in 1.5% agarose gel.
PCR analysis of cry gene families: PCR analysis was performed to identify the cry genes. For the detection of cry1, cry2, cry3, cry4, cry7 and 8 gene families, primers designed as per Ben-Dov et al. (1997) were used. Screening for the presence of cry1-type genes was done by 2 sets of primers designed as per Ceron et al. (1994) and Juarez-Perez et al. (1997). PCR was carried out in thermal cycler (Gene Amp) in 50 μL of reaction mixture containing 50 ng plasmid DNA, PCR buffer (10X) 5 μL; MgCl2 (25 mM) 5 μL; dNTPs (2 mM) 5 μL; Forward and reverse primers 1 μM each and Taq DNA polymerase 1.0 U and sterile distilled water. The programme for reaction was: Step 1: 94°C for 2 min; Step 2: 94°C for1 min; Step 3: 45°C for 1; Step 4: 72°C for 2 min; Step 5: 72°C for 10 min. The steps 2, 3 and 4 were repeated 30 times and the PCR product are analyzed in 1.5% agarose gel.
PCR amplification of full length cry genes: For amplifying full length cry genes Primers designed as per Stobdan et al. (2004) were used. The 50 μL reaction mixture was prepared {1 μg plasmid DNA, PCR buffer (10X) 5 μL; MgCl2 (25 mM) 5 μL; dNTPs (2 mM) 7 μL; Forward and reverse primers 1 μM each and Taq DNA polymerase 1.0 U and sterile distilled water} and amplification was achieved through 5 steps {Step 1: 94°C for 3 min ; Step 2: 94°C for 1 min; Step 3: 45°C for 1; Step 4: 72°C for 4 min; Step 5: 72°C for 20 min, in this Steps 2, 3 and 4 were repeated 35 times} in a thermal cycler (Gene Amp).
Preparation of spore crystal complex of Bt isolates by acetone precipitation: The method used was modified from that of Dulmage et al. (1970). The culture was grown in 250 mL LB medium at 30°C for 72 h in the shaker. Then pH of the culture was adjusted to 7.0. and cells were harvested by centrifugation (8000 rpm; 15 min). Then the cells were resuspended in 0.1 volume 6% lactose solution and stirred on a magnetic stirrer for 10 min. After this 5 volumes of ice cold acetone was added to the flask and stirred for 30 min. Then the suspension was filtered through Whatman No. 1 filter paper with suction by suction pump. The residue was collected and once again acetone extracted and dried in desiccator overnight. The powder was weighed and stored at 4°C for further use.
Sodium dodecyl sulfate-poly acrylamide gel electrophoresis (SDS-PAGE): Minigel casting apparatus (Bangalore Genei) was used. In the bottom 12% resolving gel was casted by mixing 2 mL of solution A (30 % (w/v) acrylamide and 0.8% (w/v) bis-acrylamide), 1.25 mL Solution B (1.5 M Tris-HCl (pH 8.8) and 0.4% SDS), 1.75 mL distilled water, 25 μL Ammonium per sulfate and 10 μL TEMED. On top of this, stacking gel was casted by mixing 0.33 mL Solution A, 0.50 mL Solution C (0.5 M Tris-HCl (pH 6.8) and 0.4% SDS), 1.1 mL double distilled water, 25 μL Ammonium persulphate and 10 μL TEMED. Endotoxin from Bt isolates was prepared by taking 3 loopful of culture from 72 h grown culture into a tube containing 1 mL of ice cold 0.5 M NaCl. Then the tube was centrifuged at 10,000 rpm for 5 min in microfuge and cells were pelleted. These cells were once washed with sterile distilled water. Then the cell pellet was resuspended in 100 μL of sample buffer (60 mM Tris buffer (pH 6.8), 25% glycerol, 2% SDS, 1% Bromophenol blue) and boiled for 5 minutes and then the sample was spun for 30-60 second in centrifuge. Thirty μL of this was loaded on to the gel. Gels were stained with Coomassie Brilliant Blue R-250.
Quantification of Bt toxins: Quantification of toxin protein present in samples was performed by elution of stained bands from SDS-PAGE as described by Ball (1986). Protein band was cut and added along with 1 mL of extracting solution (3% SDS in 50% isopropanol) and incubated at 37°C for 24 h after covering with paraffin in glass test tube. After this the tubes were cooled at room temperature and supernatant was pipetted out with Pasteur pipette in 1 mL cuvettes and absorbance was taken at 595 nm. BSA standard curve was used to quantify the endotoxin. To prepare BSA standard curve, 4 different known concentrations of BSA were run on the gel and eluted as mentioned above and absorbance at 595 nm was taken. Amount of endotoxin was expressed as 100 mg of acetone powder.
Insect bioassay for Bt toxins: The insecticidal activity of Bt isolates towards diamondback moth (Plutella xylostella) was evaluated by leaf dip method. Cabbage leaf discs of 4.5 cm diameter were washed in 0.1% Triton X-100 and air dried. These leaf discs were dipped in spore crystal complex containing 100 ppm of total protein for 10 sec and then air dried for 20 min. These discs were placed in plastic containers having blotting paper in it. Then six days old ten larvae of P. xylostella were transferred to the treated leaf disc in each container. Mortality rates were observed after 48 hours. These rates were corrected according to Abbott (1925) and then statistically analyzed by IRRISTAT software. Each treatment was replicated 4 times and Bt subsp. kurstaki (HD1) was used as positive control and leaf disc treated with 0.1% Triton X-100 alone was used as control.
RESULTS
Genomic DNA was isolated from 27 putative native Bt isolates recovered from phylloplane. PCR was performed to amplify a highly conserved Internal Transcribed Spacer (ITS) region between 16 S and 23 S rRNA genes for molecular characterization of phylloplane Bt isolates. The expected 234 bp PCR product was seen in 10 out of 27 isolates. A few additional bands of about 400 bp were also observed with lighter intensity. These 10 isolates (SK-208, SK-213, SK-214, SK-217, SK-219, SK-220, SK-222, SK-223, SK-229 and SK-232) were taken for further analysis. The Bt strains used as reference also amplified the expected 234 bp band (Fig. 1). From these 10 Bt isolates, plasmid DNA was isolated for screening for the presence of different types of cry genes.
PCR analysis of Bt isolates for the presence of different cry gene families: Presence of cry gene families in 10 Bt isolates was examined by PCR with primer sets designed as per Ben-Dov et al. (1997). The expected band of 277 bp corresponding to the C-terminal region of cry1 gene family was observed in 4 isolates namely SK-222, SK-223, SK-229 and SK-232 (Fig. 2a). Bt subsp. israelensis (4Q5) was used as a negative control and did not show any amplification as expected. PCR product of 701 bp corresponding to cry2 gene family was observed in only one isolate, SK-222 (Fig. 2b). The reference strains Bt subsp. galleriae (4G6) and Bt subsp. aizawai (4J4), used as positive control, also showed the expected band. PCR amplification of cry3, cry4 and cry7 and 8 gene families was not observed in any of the isolates as well as in the reference strains (data not shown).
PCR analysis of Bt isolates for the presence of specific genes of cry1 gene family: Four Bt isolates namely SK-222, SK-223, SK-229 and SK-232 , which were positive for the presence of cry1 gene family, were selected for further screening using 2 different sets of primers designed by Ceron et al. (1994) and Juarez-Perez et al. (1997) for detection of specific genes of cry1 gene family.
PCR screening with primer sets designed as per Ceron et al. (1994). Amplification of the expected 246 bp band, corresponding to cry1Aa and cry1Ad genes was observed in all the isolates SK-222, SK-223, SK-229 and SK-232. ECE52 strain was used as positive reference strain and also showed the expected band (Fig. 3a). The expected band of 171 bp corresponding to cry1Ad gene was observed in isolate SK-222 and in the reference strain Bt subsp. aizawai (4J4) (Fig. 3b). The presence of cry1Ab and cry1Ac genes was detected in all isolates along with the reference strain Bt subsp. finitmus (4B2) used as positive control. Using another set of primers corresponding to cry1Ac gene the expected band of 180 bp was seen in all the isolates (Fig. 3c). All the isolates also showed the presence of cry1B gene as indicated by the presence of expected band of 367 bp (Fig. 3d). The positive reference strains used were Bt subsp. thuringiensis (1715) and ECE128. Both the strains showed the expected band.
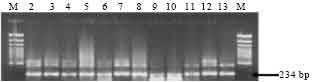 | |
| Fig. 1: | PCR amplification of 16S-23S rRNA spacer region of Bt isolates. M--100 bp Marker, Lane 2: SK-208, Lane 3: SK-213, Lane 4: SK-214, Lane 5: SK-217, Lane 6: SK-219, Lane 7: SK-220, Lane 8: SK-222, Lane 9: SK-223, Lane 10: SK-229, Lane 11: SK-232, Lane 12: 4T1, Lane 13: 4B2 |
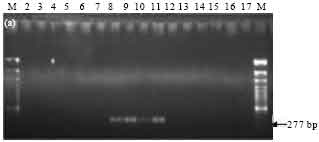 | |
| Fig. 2a: | PCR amplification of cry1 type genes of Bt isolates using primers designed as per Ben-Dov et al. (1997). M-100 bp marker, Lane 2: SK-208, Lane 3: SK-213, Lane 4: SK-214, Lane 5: SK-217, Lane 6: SK-219, Lane 7: SK-220, Lane 8: SK-222, Lane 9: SK-223, Lane 10: SK-229, Lane 11: SK-232, Lane 12: 4G6, Lane 13: 4J4, Lane 14: 4K1, Lane 15: HD1, Lane 16: 4Q5 |
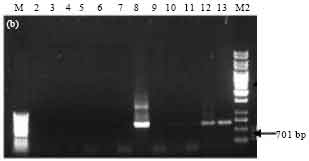 | |
| Fig. 2b: | PCR amplification of cry2 type genes of Bt isolates using primers designed as per Ben-Dov et al. (1997). M-50 bp marker, Lane 2: SK-208, Lane 3: SK-213, Lane 4: SK-214, Lane 5: SK-217, Lane 6: SK-219, Lane 7: SK-220, Lane 8: SK-222, Lane 9: SK-223, Lane 10: SK-229, Lane 11: SK-232, Lane 12: 4G6, Lane 13: 4J4, M2: 1 kb ladder |
Screening with primers specific for cry1C and cry1D genes did not give the expected band sizes of 130 and 290 bp, respectively (data not shown).
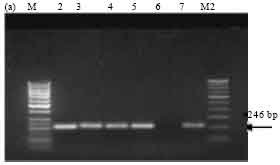 | |
| Fig. 3a: | PCR amplification of cry1Ab, Ac type genes of Bt isolates using primers designed as per Ceron et al. (1994). M-50 bp marker, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: ECE52, Lane 7: 4B2, M2: 100 bp ladder |
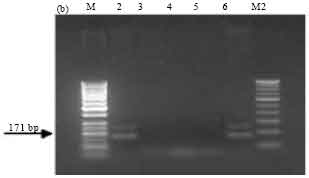 | |
| Fig. 3b: | PCR amplification of cry1Ad type genes of Bt isolates using primers designed as per Ceron et al. (1994). M-50 bp marker, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: 4J4, M2: 100 bp ladder |
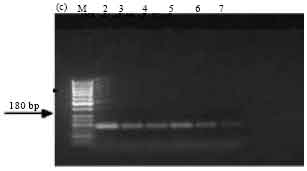 | |
| Fig. 3c: | PCR amplification of cry1Ac type genes of Bt isolates using primers designed as per Ceron et al. (1994). M-50 bp marker, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: ECE52, Lane 7: 4F3, M2: 100 bp ladder |
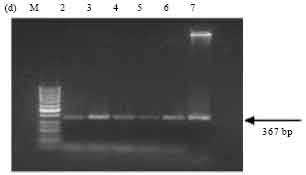 | |
| Fig. 3d: | PCR amplification of cry1B type genes of Bt isolates using primers designed as per Ceron et al. (1994). M-50 bp marker, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: 1715, Lane 7: ECE128 |
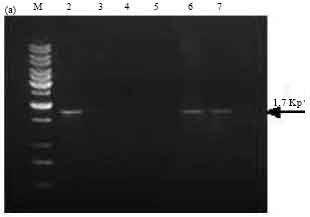 | |
| Fig. 4a: | PCR amplification of cry1A type genes of Bt isolates using primers designed as per Juarez-Perez et al. (1997). M: 1 kb Ladder, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: 4J4, Lane 7: 4G6 |
PCR screening of Bt isolates with primer sets designed as per Juarez-Perez et al. (1997). Amplification of the expected 1.7 kb band corresponding to cry1A was observed in isolate SK-222 and reference strains Bt subsp. aizawai (4J4) and Bt subsp. galleriae (4G6) (Fig. 4a). Isolate SK-222 gave the amplicon size 1.1 kb with primers corresponding to cry1Aa gene (Fig. 4b). It also gave positive signal using the primers corresponding to cry1Ab and cry1Ac by amplifying the expected PCR product of 1.3 and 0.84 kb size, respectively. Other isolates did not yield the PCR product using these primers except isolate SK-223, which showed the presence of cry1Ac gene (Fig. 4c, d). Amplification of cry1Ae gene was observed in two Bt isolates namely SK-222 and SK-223 using specific primers (Fig. 4e).
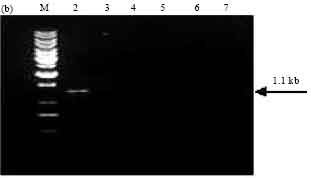 | |
| Fig. 4b: | PCR amplification of cry1Aa type genes of Bt isolates using primers designed as per Juarez-Perez et al. (1997). M: 1 kb Ladder, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: HD1, Lane 7: 4G6 |
 | |
| Fig. 4c: | PCR amplification of cry1Ab type genes of Bt isolates using primers designed as per Juarez-Perez et al. (1997). M: 1 kb Ladder, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: HD1, Lane 7: 1715 |
Amplification of the expected 1.3 kb band using primers specific for cry1B gene was observed only in isolate SK-229. Other isolates failed to give the particular band (Fig. 5a). Presence of cry1D gene was detected in all isolates except SK-223 including the reference strain Bt subsp. aizawai (4J4) with the expected band size of 1.1 kb (Fig. 5b). The amplification of cry1C, cry1E, cry1F and cry1G genes was not in any of the isolates with corresponding primers (data not shown).
PCR amplification of full length cry1 genes: PCR amplification of full length cry1Aa, cry1Ab, cry1Ac and cry1D genes in the four cry1 positive isolates was carried out using primers corresponding to the full length cry1Ab,c gene as described in the materials and methods.
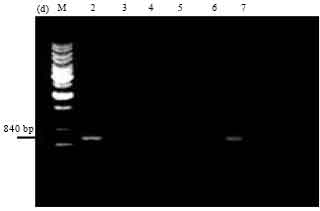 | |
| Fig. 4d: | PCR amplification of cry1Ac type genes of Bt isolates using primers designed as per Juarez-Perez et al. (1997). M: 1 kb Ladder, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: HD1, Lane 7: 4F3 |
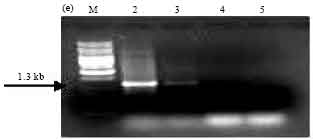 | |
| Fig. 4e: | PCR amplification of cry1Ae type genes of Bt isolates using primers designed as per Juarez-Perez et al. (1997). M: 1 kb Ladder, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232 |
Bt isolate SK-222 alone gave the expected amplicon size of 3.4 kb. Other isolates did not give any band while reference strains namely Bt subsp. kenyae (4F3) and ECE52 gave the full length gene products (Fig. 6).
SDS-PAGE analysis of phylloplane isolates: In the SDS-PAGE analysis, isolate SK-222 showed the 3 prominent protein bands with one with the size of ~130 kDa corresponding to that of the cry1 δ-endotoxin. It also gave bands with the size around 65 kDa and another around 27 kDa, corresponding to cry2 and Cyt protein respectively. Isolate SK-223 gave ~130 kDa band along with 27 kDa.
 | |
| Fig. 5a: | PCR amplification of cry1B type genes of Bt isolates using primers designed as per Juarez-Perez et al. (1997). M: 1 kb Ladder, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: 4A6, Lane 7: 4K1, M2: Lambda DNA digested with Hind III and EcoRI |
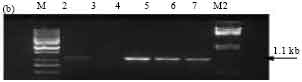 | |
| Fig. 5b: | PCR amplification of cry1D type genes of Bt isolates using primers designed as per Juarez-Perez et al. (1997). M: 1 kb Ladder, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: 4J4, M2: Lambda DNA digested with Hind III and EcoRI |
 | |
| Fig. 6: | PCR amplification of full length cry1Ac gene of Bt isolates. M: 1 kb Ladder, Lane 2: SK-222, Lane 3: SK-223, Lane 4: SK-229, Lane 5: SK-232, Lane 6: 4F3, Lane 7: ECE52 |
Other isolates namely SK-229 and SK-232 gave only the 27 kDa band while the 130 kDa band was not visible. The reference strain ECE52, which has expressible cloned gene, also showed the 130 kDa band (Fig. 7).
Quantification of δ-endotoxin using SDS-PAGE: For the quantification of endotoxin, the SDS-PAGE was repeated along with the 4 known concentrations of BSA.
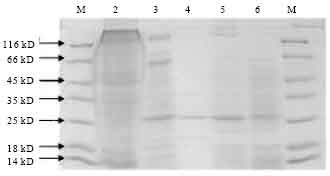 | |
| Fig. 7: | SDS-PAGE of phylloplane isolates. M: Protein molecular weight marker (medium weight), Lane 2: ECE52, Lane 3: SK-222, Lane 4: SK-229, Lane 5: SK-223, Lane 6: SK-232 |
| Table 1: | Analysis of variance (ANOVA) of table of treatment means for 48 h after application |
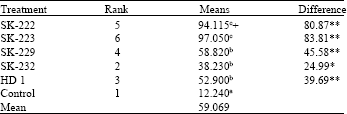 | |
| a:Data from 4 replications of 10 larvae each., **Significant at 1% level, *Significant at 5% level, +: Means followed by a common letter are not significantly different at 5 % level | |
The bands were eluted and endotoxin was quantified.
Insect bioassay: Insect bioassay was conducted with 6 days old larvae of Diamondback moth (Plutella xylostella) by leaf dip method. Each treatment was replicated four times with ten larvae in each replication. Maximum mortality was observed in SK-223 after 48 h of treatment closely followed by SK-222, SK-229 and SK-232. Among the treatments, the toxicity of SK-222 and SK-223 was significantly higher than that of SK-229, SK-232 and Bt subsp. kurstaki (HD1) at 5% level (Table 1).
DISCUSSION
Bt-based biopesticides have been in use for the past five decades. Bt transgenic plants expressing cry proteins have also been developed and commercialized. Although a large number of cry genes are known, due to development of resistance in pests towards the frequently used cry1Aa, cry1Ab and cry1Ac genes, as also to broaden the insecticidal spectrum to cover more number of target insect pests, search for novel types of insecticidal cry genes has been an ongoing effort worldwide. Bt isolates have been recovered from the phylloplanes of leguminous crops in the New Delhi region of India and have been characterized with respect to their growth characteristics, antibiotic resistance and crystal morphology (Kaur and Singh, 2000b). The present investigation was undertaken with the objective of molecular characterization of these Bt isolates in terms of presence of different cry gene families and specific types of cry genes, protein profiles and insecticidal activity.
The 16 S-23 S internal transcribed rDNA spacer region has been used as a taxonomic tool for characterization of bacteria due to high conservation of this sequence (Jensen et al., 1993). For Bt strains, this sequence is amplified as a 234 bp band using specific primers (Hansen and Hendriksen, 2001). This expected band of 234 bp was observed in 10 Bt isolates and these were selected for further PCR screening for the presence of cry genes. An additional band of ca. 400 bp was also seen in some Bt isolates which could be due to polymorphism among these isolates.
The cry genes are generally located on mega-plasmids of size >30 kDa (Gonzalez et al., 1981). The plasmid profile of all the 10 Bt isolates indicated the presence of a high molecular weight plasmid and presence of additional plasmids in some Bt isolates. The presence of cry1, 2, 3, 4, 7 and 8 gene families was investigated in Bt isolates by using 5 pairs of oligonucleotide primers corresponding to highly conserved regions in these cry gene families as designed by Ben-Dov et al. (1997). The cry1 gene family was found to be present in 4 isolates namely SK-222, SK-223, SK-229 and SK-232 as indicated by PCR amplification of 277 bp band. However, this band was not seen in the Bt strains used as reference. This suggests that the primers designed by Ben-Dov et al. (1997) did not anneal to the cry1 type genes present in these reference strains. The cry2 gene family was found to be present only in one isolate namely SK-222 as indicated by the presence of expected PCR product of 701 bp. This band was also seen in the Bt subsp aizawai (4J4) and Bt subsp. galleriae (4G6) used as positive references. The cry3, cry4 and cry7 and 8 gene families were found to be absent in all the Bt isolates as the expected PCR product was not observed. The cry1 gene family has been reported to be the most abundant in Bt isolates from other collections as well (Wang et al., 2003). Presence of both cry1 and cry2 gene families in the Bt isolate SK-222 seems promising, since based on cry gene profile this isolate is predicted to have toxicity both towards lepidopteran and dipteran insects.
Four Bt isolates namely SK-222, SK-223, SK-229 and SK-232, which were found to be positive for the presence of cry1 gene family by PCR amplification, were further screened for the presence of cry1 type genes using the set of primers designed as per Ceron et al. (1994). The cry1Aa, cry1Ad, cry1Ab, cry1Ac and cry1B were found to be present in all the four isolates, while cry1C and cry1D genes were found to be absent. Using a separate set of primers for cry1Ad gene, this gene was found to be present only in the isolate SK-222. With the primer sets designed as per Juarez-Perez et al. (1997). cry1Aa, cry1Ab, cry1Ae and cry1Ac genes were found to be present in only one isolate namely SK-222. The cry1Ae was also found in isolate SK-223. Only one isolate, SK-229 showed the presence of cry1B, whereas, PCR amplification with primers designed as per Ceron et al. (1994) indicated the presence of cry1B gene in all the isolates. The presence of cry1C was not observed in any of the isolates as was observed also using primers designed as per Juarez-Perez et al. (1997). Presence of cry1D gene was seen in 3 isolates, using the primer pair designed as per Juarez-Perez et al. (1997), where as this gene was indicated to be absent using the primer pair as designed by Ceron et al. (1994). Presence of cry1, D, E, F and G genes were not seen in any of the four isolates using primers designed as per Juarez-Perez et al. (1997). In the PCR strategy of Juarez-Perez et al. (1997), a common reverse primer and specific forward primers are used for the identification of cry1 genes. The expected PCR products obtained are also relatively longer as compared with the expected PCR products obtained using primers as designed by Ceron et al. (1994). As these two primer sets correspond to different regions of homology among cry1 type genes, detection of cry1 type genes is dependent on the extent of conservation of the specified homology regions in the cry1 type genes present in different Bt isolates. Use of more than 1 set of primers is thus desirable for the PCR identification of cry1 type genes in Bt isolates.
PCR amplification of full length cry1Ab and cry1Ac genes was carried out using a set of primers specially designed in our lab (Stobdan et al., 2004). Amplification of full length cry1Ab and cry1Ac genes was observed in isolate SK-222. Amplification of full length cry1Ac gene was also observed in Bt subsp. kenyae (4F3) and ECE52 used as reference.
A band of ca. 130 kDa corresponding to cry1 protein was seen in isolates SK-222 and SK-223. SK-222 also exhibited the presence of ~65 kDa band corresponding to cry2 protein. These results corroborate the presence of cry1 and cry2 genes in SK-222 and of cry1 gene in SK-223 as determined by PCR analysis for the presence of cry genes.
Insecticidal activity of four cry1 positive Bt isolates towards P. xylostella was investigated.
| Table 2: | Characterization of Bt isolates from leguminous phylloplanes |
 | |
| **cry protein-As observed by SDS-PAGE, *Insecticidal activity-Average of corrected mortality percentage after 48 hours of treatment from 4 replications of 10 larvae each | |
Isolate SK-223 was found to be highly toxic with mortality of 97.05%, closely followed by SK-222 with the mortality 95%. SK-229 and SK-232 have relatively lower mortality rates of 58.25 and 31.25%, respectively. SK-223 and SK-222 have given significantly higher mortality than Bt subsp. kurstaki (HD1). It is interesting to note that in SK-223 amplification of full length cry1Ab and cry1Ac gene was not observed. This indicates the presence of other highly toxic cry gene in this isolate, which can be confirmed by further investigation. Full length cry1Ab-type gene was amplified from isolate SK-222, cloned into pGEM-T vector and sequenced (GenBank accession No. DQ023297). Sequence analysis showed it to be identical to cry1Ab18 gene (http://www.lifesci.sussex.ac.uk/home/Neil_ Crickmore/Bt/, GenBank accession No. AY319967) isolated previously from a Bt isolate recovered from soil of Ladakh region in our laboratory.
Since a significant number of pests have been reported to have developed resistance to the available cry proteins, novel genes which have increased insecticidal activity and broader spectrum extending to several agronomically important insect pests as well as which have different receptor sites or mode of action in the target insects midgut, are needed for better resistance management (Kaur, 2012). Molecular characterization of Bt isolates based on the PCR analysis, SDS-PAGE and insect bioassay are depicted in Table 2. Isolates SK-222 and SK-223 are promising Bt isolates from leguminous phylloplanes which can be very useful for crop protection either as developed into biopesticides as well as for gene isolation.
ACKNOWLEDGMENTS
Authors thank Rakesh Narula and Sunita Srivastva for excellent technical assistance and Dr. G.T. Gujar, Division of Entomology, IARI, New Delhi, for providing insect culture facilities. Part of this work was submitted as M.Sc. thesis of JK to P.G. School, IARI.
REFERENCES
- Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Ali, B.A., H.H. Salem, X.M. Wang, T.H. Huang, Q.D. Xie and X.Y. Zhang, 2010. Effect of Bacillus thuringiensis var. Israelensis endotoxin on the intermediate snail host of Schistosoma japonicum. Curr. Res. Bacteriol., 3: 37-41.
CrossRefDirect Link - Anwar, R., S. Karim and S. Riazuddin, 1999. Entomocidal activity of Bacillus Thuringiensis transgenic rice plants against rice leaffolder, Cnaphalocrocis Medinalis (Lepidoptera: Pyralidae). Pak. J. Biol. Sci., 2: 1472-1477.
CrossRefDirect Link - Apaydin, O., A.F. Yenidunya, S. Harsa and H. Gunes, 2005. Isolation and characterization of Bacillus thuringiensis strains from different grain habitats in Turkey. World J. Microbiol. Biotechnol., 21: 285-292.
Direct Link - Ball, E.H., 1986. Quantification of proteins by elution of coomassie brilliant blue R-250 from stained bunds after Sodium Dodecyl Sulfate-Polyacrylamide gel electrophoresis. Anal. Bochem., 155: 23-27.
PubMed - Beard, C.E., C. Ranasinghe and R.J. Akhurst, 2001. Screening for novel cry genes by hybridization. Lett. Applied Microbiol., 33: 241-245.
Direct Link - Ben-Dov, E., A. Zaritsky, E. Dahan, Z. Barak and R. Sinai et al., 1997. Extended screening by PCR for seven cry-group genes from field-collected strains of Bacillus thuringiensis. Applied Environ. Microbiol., 63: 4883-4890.
Direct Link - Beron, C.M., L. Curatti and G.L. Salerno, 2005. New strategy for identification of novel cry-type genes from Bacillus thuringiensis strains. Applied Environ. Microbiol., 71: 761-765.
Direct Link - Birnboim, H.C. and J. Doly, 1979. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res., 7: 1513-1523.
CrossRefPubMedDirect Link - Ceron, J., Covarrubias, L., Quintero, R., Ortiz and A. Ortiz et al., 1994. PCR analysis of cry1 insecticidal family genes from Bacillus thuringiensis. Appl. Environ. Microbiol., 60: 353-356.
Direct Link - Chandra, A. and D. Pental., 2003. Regeneration and genetic transformation of grain legumes: An overview. Curr. Sci., 84: 381-387.
Direct Link - Crickmore, N., D.R. Zeigler, J. Feitelson, E. Schnepf and J. Van Rie et al., 1998. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev., 62: 807-813.
Direct Link - Dulmage, H.T., J.A. Correa and A.J. Martinez, 1970. Coprecipitation with lactose as means of recovering the spore crystal complex of Bacillus thuringiensis. J. Inverteb. Pathol., 15: 15-20.
PubMed - Van Frankenhuyzen, K., 2009. Insecticidal activity of Bacillus thuringiensis crystal proteins. J. Invert. Pathol., 101: 1-16.
PubMed - Gonzalez, J.M., H.T. Dulmage and B.C. Carlton, 1981. Correlation between specific plasmids and d-endotoxin production in Bacillus thuringiensis. Plasmid, 5: 351-365.
PubMedDirect Link - Hansen, B.M. and N.B. Hendriksen, 2001. Detection of enterotoxic Bacillus cereus and Bacillus thuringiensis strains by PCR analysis. Applied Environ. Microbiol., 67: 185-189.
PubMed - Hansen, B.M., P.H. Damgaard, J. Eilenberg and J.C. Pedersen, 1998. Molecular and phenotypic characterization of Bacillus thuringiensis isolated from leaves and insects. J. Invertebrate Pathol., 71: 106-114.
CrossRef - Jara, S., P. Maduell and S. Orduz, 2006. Diversity of Bacillus thuringiensis strains in the maize and bean phylloplane and their respective soils in Colombia. J. Appl Microbiol., 101: 117-124.
PubMed - Jensen, M.A., J.A. Webster and N. Straus, 1993. Rapid identification of bacteria on the basis of polymerase chain reaction-amplified ribosomal DNA spacer polymorphisms. Appllied Envion. Microbiol., 59: 945-952.
Direct Link - Juarez-Perez, V.M., M.D. Ferrandis and R. Frutos, 1997. PCR-based approach for detection of novel Bacillus thuringiensis cry genes. Applied Environ. Microbiol., 63: 2997-3002.
Direct Link - Kalman, S., K.L. Kiehne, J.L. Libs and T. Yamamoto, 1993. Cloning of a novel cryIC type gene from a strain of Bacillus thuringiensis subsp. Galleriae. Applied Environ. Microbiol., 59: 1131-1137.
Direct Link - Katara, J., R. Deshmukh, N.K. Singh and S. Kaur, 2012. Molecular typing of native Bacillus thuringiensis isolates from diverse habitats in India using REP-PCR and ERIC-PCR analysis. J. Gen. Applied Microbiol., 58: 83-94.
CrossRef - Kaur, S., 2000. Molecular approaches towards development of novel Bacillus thuringiensis biopesticides. World J. Microbiol. Biotechnol., 16: 781-793.
CrossRefDirect Link - Kaur, S., 2006. Molecular approaches for identification and construction of novel insecticidal genes for crop protection. World J. MIcrobiol. Biotechnol., 22: 233-253.
CrossRefDirect Link - Kaur, S. and A. Singh, 2000. Natural occurrence of Bacillus thuringiensis in leguminous phylloplanes in the New Delhi region of India. World J. Microbiol. Biotech., 16: 679-682.
CrossRefDirect Link - Khan, M.A., R. Makhdoom, T. Husnain, M.Z. Saleem, K. Malik, Z. Latif, I. Altosaar and S. Riazuddin, 2001. Expression of Bt gene in a dicot plant under promoter derived from a monocot plant. Pak. J. Biol. Sci., 4: 1518-1522.
CrossRefDirect Link - Martinez, C. and P. Caballero, 2002. Contents of Cry genes and insecticidal toxicity of Bacillus thuringiensis strains from terrestrial and aquatic habitats. J. Applied Microbiol., 92: 745-752.
PubMed - Meadows, M.P., D.J. Ellis, J. Butt, P. Jarrett and H.D. Burges, 1992. Distribution, frequency and diversity of Bacillus thuringiensis in an animal feed mill. Applied Environ. Microbiol., 58: 1344-1350.
Direct Link - Meena, R.K., G.G.K. Kumari, A. Govind, T. Gujar and S. Kaur, 2012. Screening of Bacillus thuringiensis Isolates Recovered from Diverse Habitats in India for the Presence of cry1A-type Genes and Cloning of a cry1Ac33 Gene Toxic to Helicoverpa armigera (American Bollworm) Asian J. Biotechnol., 4: 53-69.
CrossRef - Noguera, P.A. and J.E. Ibarra, 2010. Detection of new cry genes of Bacillus thuringiensis by use of a novel PCR primer system. Applied Environ. Microbiol., 76: 6150-6155.
PubMed - Porcar, M. and V. Juarez-Perez, 2003. PCR based identification of Bacillus thuringiensis pesticidal crystal genes. FEMS Microbiol. Rev., 26: 419-432.
Direct Link - Stobdan, T., S. Kaur and A. Singh, 2004. Cloning and nucleotide sequence of a novel Cry gene from Bacillus thuringiensis. Biotechnol. Lett., 26: 1153-1156.
PubMed - Swiecicka, I., D.K. Bideshi and B.A. Federici, 2008. Novel isolate of Bacillus thuringiensis subsp. thuringiensis that produces a quasicuboidal crystal of Cry1Ab21 toxic to larvae of Trichoplusia ni. Applied Environ. Microbiol., 74: 923-930.
Direct Link - Uribe, D., W. Martinez and J. Ceron, 2003. Distribution and diversity of cry genes in native strains of Bacillus thuringiensis obtained from different ecosystems from Colombia. J. Invertebr. Pathol., 82: 119-127.
PubMed - Wang, J., A. Boets, J. van Rie and G. Ren, 2003. Characterization of cry1, cry2 and cry9 genes in Bacillus thuringiensis isolates from China. J. Inverteb. Pathol., 82: 63-71.
CrossRefDirect Link