
Rajendra K. Meena
National Research Centre on Plant Biotechnology, Indian Agricultural Research Institute Campus, New Delhi 110012, India
Gunda Gouthami Krishna Kumari
National Research Centre on Plant Biotechnology, Indian Agricultural Research Institute Campus, New Delhi 110012, India
Alpana Govind
Division of Entomology, Indian Agricultural Research Institute, 110012, New Delhi
T. Gujar
Division of Entomology, Indian Agricultural Research Institute, 110012, New Delhi
Sarvjeet Kaur
National Research Centre on Plant Biotechnology, Indian Agricultural Research Institute Campus, New Delhi 110012, India
Asian Journal of Biotechnology
Year: 2012 | Volume: 4 | Issue: 2 | Page No.: 53-69







ABSTRACT
Helicoverpa armigera (American bollworm) is a severe pest of many economically important crops such as cotton, pigeon pea, chickpea and tomato in the Indian subcontinent. Insecticidal cry genes from the bacterium Bacillus thuringiensis (Bt) have been used for developing transgenic crops. Transgenic cotton expressing cry1Ac gene has shown good level of protection from H. armigera. However, there is a threat of eventual development of resistance in insects upon large-scale cultivation of transgenic crops. Therefore, prospecting of Bt strains for new types of cry genes is desirable. This study was undertaken for screening of native Bt isolates for the presence of cry1Aa,b,c-type genes with the objective of cloning, sequence analysis, expression and evaluation of toxicity. Sixty three native Bt isolates recovered from different soil and grain samples from diverse agricultural and non-agricultural locations in India, along with 10 known Bt strains used as reference, were screened for the presence of full length cry1A-type genes by Polymerase Chain Recation (PCR) using a set of primer. The full length gene was obtained in the native Bt isolate SK-711 recovered from Red gram field, Lam, Guntur Andhra Pradesh and in 3 Bt strains. The gene from the native Bt isolate was cloned into an E. coli expression vector. The sequence of the cloned gene (GenBank accession No. GQ866913) was analyzed by comparison with previously known cry1A-type genes and was assigned the name cry1Ac33 by the Bacillus thuringiensis Nomenclature Committee. The gene was expressed in E. coli and evaluated for toxicity towards H. armigera. The cry1Ac33 gene, cloned from a native Bt isolate, has been found to be more toxic towards H. armigera than the holotype cry1Ac1 used as a control, based on LC50 toxicity analysis.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: February 10, 2012;
Published: March 12, 2012
How to cite this article
Rajendra K. Meena, Gunda Gouthami Krishna Kumari, Alpana Govind, T. Gujar and Sarvjeet Kaur, 2012. Screening of Bacillus thuringiensis Isolates Recovered from Diverse Habitats in India for the Presence of cry1A-type Genes and Cloning of a cry1Ac33 Gene Toxic to Helicoverpa armigera (American Bollworm). Asian Journal of Biotechnology, 4: 53-69.
DOI: 10.3923/ajbkr.2012.53.69
URL: https://scialert.net/abstract/?doi=ajbkr.2012.53.69
DOI: 10.3923/ajbkr.2012.53.69
URL: https://scialert.net/abstract/?doi=ajbkr.2012.53.69
INTRODUCTION
Bacillus thuringiensis (Bt) is an aerobic, gram-positive, spore-forming bacterium producing crystal proteins (Cry) which are selectively toxic to target insects (Feitelson et al., 1992; (Karim et al., 1999; Garcia-Robles et al., 2001; Amjad et al., 2001; Zia-ur-Rehman et al., 2002; Zaied et al., 2003; De Escudero et al., 2006; Halima et al., 2006; Ali et al., 2010). Cry proteins act by insertion into the microvillar brush-border membranes in the midgut of susceptible insects, leading to disruption of osmotic balance, lysis of epithelial cells and eventually death of insect (Schnepf et al., 1998). Insect resistant Bt transgenic crops were first developed using native cry1Ac gene (Barton et al., 1987; Anwar et al., 1999; Khan et al., 2001). The total acreage of transgenic crops has been steadily increasing with countries including 10 developed and 19 developing countries having adopted commercial cultivation of transgenic crops on 140 million hectares in 2010 (Gatehouse, 2008; James, 2010). The use of Bt crops has resulted in increased yields and significant reductions of insecticide application, thus providing economic and environmental benefits (Brookes and Barfoot, 2008; Carpenter, 2010).
The bollworm, Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) is a polyphagous pest of worldwide occurrence that inflicts an annual crop damage in India worth US$ 1 billion (Subramanian and Mohunkumar, 2006). It is a major pest of several economically important crops such as cotton, pigeonpea, chickpea, tomato, sorghum, okra and blackgram. Transgenic cotton expressing cry1Ac gene, released for commercial cultivation in India and China, has shown good level of protection from H. armigera (Wu et al., 2003; Choudhary and Gaur, 2010).
However, there is a threat of eventual development of resistance in H. armigera exposed to Cry1Ac protein season-long in all plant parts upon large-scale cultivation of Bt cotton (Shelton et al., 2002; Kaur, 2004). Development of second-generation Bt transgenic crops requires new insecticidal genes for stacking or pyramiding of genes wherein more than one insecticidal genes are used in combination as a resistance management strategy (Zhao et al., 2003; Kaur, 2006). Two cry genes (out of cry1Ab, cry1Ac, cry2A, cry1C) pyramided rice lines and their hybrids exhibited excellent efficacy against stem borers and leaf folders in field evaluation (Yang et al., 2011). The rapidity of development of resistance necessitates isolation of new genes for transgenic development. A significant exponential increase in the frequency of alleles conferring cry2Ab resistance in Australian field populations of H. punctigera since the adoption of a second generation Bt cotton expressing Cry2Ab protein was observed (Downes et al., 2010). Therefore, the isolation of novel cry-type genes from Bt isolates is a useful approach to tackle the development of resistance against currently deployed limited number of cry-type gene(s).
Bt isolates are being screened the world over in search of new types of insecticidal genes. Bt strains have been isolated worldwide from diverse habitats, including soil (Bernhard et al., 1997; Kaur and Singh, 2000a; Wang et al., 2003), stored grains (Meadows et al., 1992) phyllospheres (Hansen et al., 1998; Kaur and Singh, 2000b; Jara et al., 2006) and other miscellaneous habitats (Uribe et al., 2003; Apaydin et al., 2005; Stobdan et al., 2005; Martinez and Caballero, 2002). Over 200 cry genes grouped into 40 families with insecticidal spectrum extending over several invertebrate orders have been isolated from Bt strains worldwide (Crickmore et al., 1998; Wang et al., 2003; Stobdan et al., 2004; Swiecicka et al., 2008). The identification of cry genes by Polymerase Chain Reaction (PCR) has proven to be a very useful method for characterization of cry genes present in Bt strains due to its rapidity and reproducibility (Porcar and Juarez-Perez, 2003; Beron et al., 2005; Kaur, 2006).
Native Bt isolates from diverse habitats in India have been isolated and screened for the presence of different cry genes in our laboratory (Kaur and Singh, 2000a, b; Stobdan et al., 2005). A novel cry1Ab type gene has been cloned by PCR amplification with specifically designed primers from a native Bt isolate from cold desert soil of Ladakh region (GenBank Accession No. AY 319967) and named as cry1Ab18 by the Bacillus thuringiensis Nomenclature Committee (http://www.biols.susx.ac.uk/Home/Neil_Crickmore/Bt/) (Stobdan et al., 2004). A novel cry gene (GenBank accession number DQ023296) was cloned from a native Bt isolate (SK-729) recovered from stored cotton seeds and designated as cry1Ac18 by the Bacillus thuringiensis Nomenclature Committee (Kaur and Allam, 2006). In this study, sixty three native Bt isolates obtained from different soil and grain samples from diverse locations in India have been screened for the presence of cry1Aa,b,c-type genes with specially designed primers and the amplified gene has been cloned, sequenced and expressed in E. coli for analysis of toxicity towards H. armigera.
MATERIALS AND METHODS
Materials
Bacterial isolates and strains: Sixty three native Bt isolates recovered from different soil and grain samples from diverse agricultural and non-agricultural locations in India in the corresponding author Dr. S. Kaur’s laboratory were used in this study (Table 1). Bt strains and the recombinant strain ECE 53 (E. coli clone with cry1Ac1 gene) used as reference in this study were kindly provided by Dr. D.R. Ziegler, Director, Bacillus Genetic Stock Center, Ohio State University, Columbus, OH, USA to Dr. S. Kaur (Table 2).
| Table 1: | Native Bt isolates recovered from diverse habitats in India used in this study |
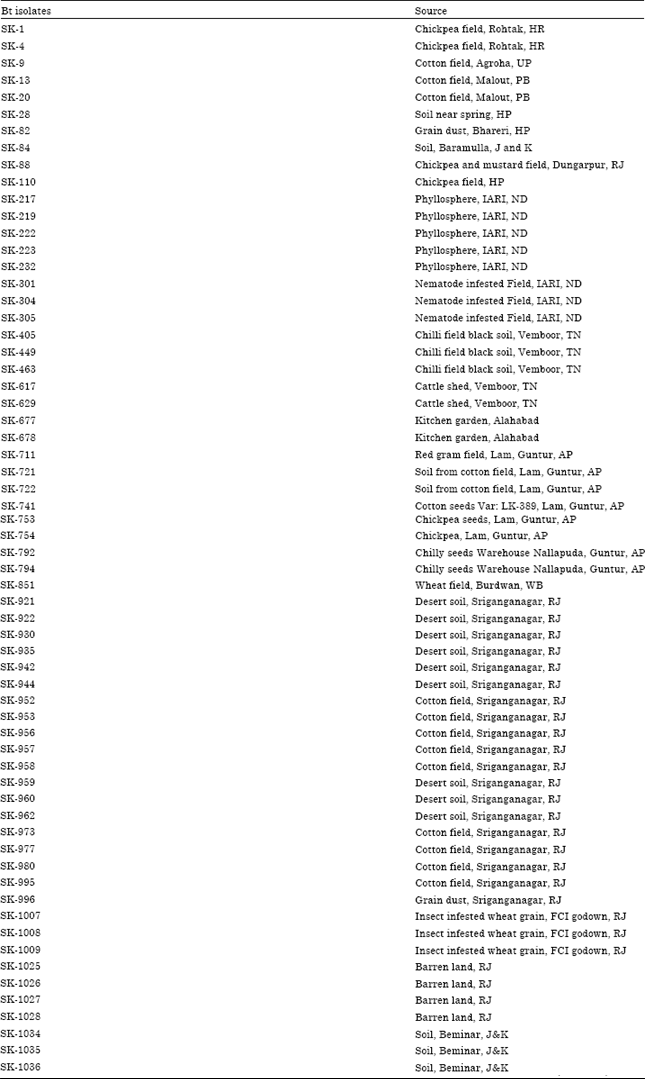 | |
| AP: Andhra Pradesh, RJ: Rajasthan, WB: West Bengal, ND: New Delhi, J&K: Jammu and Kashmir, HP: Himachal Pradesh, TN: Tamil Nadu, PB: Punjab, HR: Haryana | |
| Table 2: | Bt strains used as reference in this study |
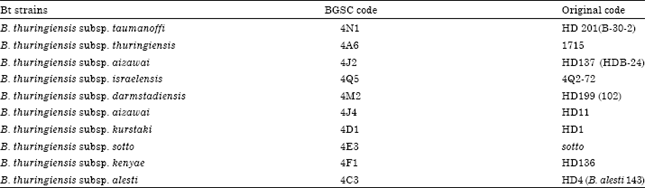 | |
| Table 3: | Characteristics of specific primers designed as per Beard et al. (2001) for amplification of full length cry1A-type genes |
 | |
Growth media for bacterial strains: Luria Bertani Agar (LA) and Luria Bertani Broth (LB) were used for the growth of E. coli and Bt strains.
Oligonucleotide PCR primers: A set of primers designed as per Beard et al. (2001) was used for screening of Bt isolates for the presence of full length cry1Aa,b,c-type genes (Table 3).
Methods
Genomic DNA extraction: Genomic DNA was extracted from Bt isolates by the method modified from Kalman et al. (1993). A single colony from a freshly streaked plate was incubated into 5 mL Luria Bertani (LB) medium containing penicillin (10 μg mL-1) and incubated at 30°C overnight with shaking at 150 rpm. This starter culture was added to 50 mL LB medium and incubated at 300C with shaking at 150 rpm to an optical density of 0.8 at 600 nm. The cells were harvested by centrifuging at 7000 rpm for 10 min (Beckman JA 20 rotor) at 4°C. The cell pellet was washed in 5 mL TES buffer (10 mM tris-HCl, 1 mM EDTA, 100 mM NaCl, pH 8.0) and suspended in 5 mL of resuspension solution (25% sucrose, 25 mM Tris-HCl, 25 mM EDTA, pH 8.0) containing 1 mg mL-1 Lysozyme. The cell suspension was incubated at 37°C for 1 h. 10% SDS was added to the suspension to a final concentration of 2% and the suspension was incubated at 500C for 15 min and then at 4°C overnight. The suspension was centrifuged at 10,000 rpm (JA 20 rotor, Beckman) for 15 min and the supernatant was carefully taken out. DNA in the supernatant was precipitated with 2 volumes of ethanol. DNA was resuspended in 10 mL TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0) containing 1 M NaCl, 10 μg mL-1 RNase and 0.6 mg mL-1 Proteinase K and incubated at 30°C for 30 min. The mixture was extracted with phenol-chloroform (1:1) and DNA was precipitated with ethanol. DNA pellet was washed once with 70% ethanol, air-dried and dissolved in 300 μL of TE buffer.
Plasmid DNA extraction: Plasmid DNA was isolated from Bt isolates and reference strains by using Qiagen Plasmid Midi kit (Qiagen, Germany). A single colony from a freshly streaked plate was inoculated into 10 mL LB medium containing penicillin (10 μg mL-1) and incubated at 30°C with shaking at 150 rpm. Overnight grown cultures were pelleted by centrifugation at 7000 rpm (Beckman JA20 rotor) for 10 min at 4°C. Method as described in the supplier’s manual was followed, with the modification of preheating of elution buffer to 50°C for isolation of large plasmids. DNA pellet was dissolved in 200 μL TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0). Plasmid DNA was size fractionated on 0.8% agarose gel along with 1 kb DNA ladder (MBI Fermentas, Germany) used as molecular weight marker. DNA bands were observed under UV in the gel documentation system (Alpha gel imager).
PCR amplification of cry1Aa,b,c-type genes: PCR was carried out in a reaction mixture of 25 μL containing DNA template 50 ng; PCR buffer with (NH4)2SO4 and MgCl2 (10X) 2.5 μL; deoxy ribonucleotide triphosphate (dNTPs) (2 mM) 2.5 μL; primers (1 μM) 1 μL each, Taq DNA polymerase 1.0 U and sterile distilled water in thermal cycler (BioRad). Amplification was carried out for 30 cycles of denaturation at 94°C for 1 min, primer annealing at 42°C for 1 min and extension at 72°C for 4 min. Denaturation in the first cycle was performed for 2 min and extension was performed for 15 min in the last cycle. PCR products were visualized on 0.8% agarose gel using 1 kb DNA ladder marker. PCR product was analyzed on 0.8% agarose gel by electrophoresis using 1 kb DNA ladder marker (MBI Fermentas).
Cloning of full-length cry1Aa,b,c-type gene: The 3.6 kb PCR product amplified with primers designed as per Beard et al. (2001) from native Bt isolate SK-711 was eluted from the gel using QIA Mini Elute gel extraction kit (Qiagen) and cloned into pQE-30 UA expression cloning vector (Qiagen, Germany) as per manufacturer’s instructions. This vector is specially designed for direct cloning of PCR product as it provided with “U” overhangs at each 3’ end, which hybridizes with, “A” overhang at each 3’ end of the PCR product amplified using Taq DNA polymerase and is also suitable for expression in E. coli. The 3.6 kb gel purified PCR product of native Bt isolate SK-711 was ligated with vector and the recombinant vector was transformed into competent Escherichia coli strain XL-1 Blue following the manufacturer’s protocol with some modifications. Recombinant clones were analyzed for the presence of insert DNA by restriction and PCR amplification of plasmid DNA of the clones. Plasmid DNA was isolated from the clones using Plasmid Mini kit (Qiagen) and digested with restriction enzymes BamHI to linearize the plasmid and with BamHI and Sal I (Fermentas) to take out the insert at 370C for 4 h. Restricted DNA was analyzed on 0.8% agarose gel by electrophoresis using 1 kb DNA ladder marker. PCR analysis of plasmid DNA of positive clones was carried out using primer pair cry1AabcF and cry1AabcR as described.
Nucleotide sequencing and analysis: Sequence determination of the clone pRKM-711 carrying 3.5 kb cry gene in the pQE-30 UA expression cloning vector was performed by service provider. Homology search was done using BLAST programme of NCBI (National Centre for Biotechnology Information) web site: http://www.ncbi.nlm.nih.gov and Multalin version 5.4.1 for multiple alignments.
Expression of cloned cry1Ac33 gene into E. coli: The recombinant clone pRKM-711 was grown in LB containing ampicillin (100 μg mL-1) at 37°C with shaking at 150 rpm and was induced to express cry1Ac33 gene with IPTG (isopropyl-β-D-galactopyranoside) at a final concentration of 1 mM for 4 h. The cells were harvested by centrifuging at 7000 rpm for 10 min (Beckman JA 20 rotor) at 4°C. SDS-PAGE analysis of protein purified from induced recombinant clone was carried out by a method modified from Lee et al. (1992) and analysed on 12% SDS-PAGE.
Solubilised toxin preparation from recombinant clone carrying cry gene in expression vector: The solubilised protein was prepared from the recombinant E. coli clone carrying cry1Ac33 gene in pQE30 UA expression vector. The recombinant clone was induced to express cry gene with IPTG at a final concentration of 1 mM for 4 h. The cells were harvested by centrifugation and after sonication, the pellet was washed thrice with wash solution 1 (0.5 M NaCl and 2% triton X-100), thrice with wash solution 2 (0.5 M NaCl) and finally thrice with double distilled water. The toxin was solubilised by incubating pellet in 2 mL of solubilisation buffer (50 mM sodium carbonate containing dithiothreitol 10 mM, pH 10.5).
| Table 4: | Protein concentration of solubilized protein preparation of recombinant E. coli clones |
 | |
| *Protein content is expressed as μg of total protein per μL of soluble protein solution | |
This toxin was used for insect bioassays after protein quantification by InvitrogenTM Qubit flourometer (Table 4). The solubilised protein preparation of the recombinant strain ECE 53 (E. coli clone with cry1Ac1 gene) was also prepared for use as the control in insect bioassays.
H. armigera rearing for evaluation of toxicity of Cry protein: Susceptibility of H. armigera collected from cotton and chickpea field from IARI location was tested by exposing neonates of this insect to different concentration of toxin mixed with artificial diet. The larvae were allowed to grow until pupation on the artificial diet. The artificial diet prepared was essentially gram based diet. Pupae were collected and kept in cage for adult emergence. The adults were paired and then kept in a cage and were given 10% honey solution for egg laying which was done on Markin cloth. The eggs, thus collected, were separated day wise and allowed to hatch. The neonates were then transferred to the artificial diet. The insects were reared at 27°C and 60-70% RH. The pupae were separated and then kept in a jar at 27°C until adult emergence. The adults were then allowed to mate and lay eggs.
Insect bioassay: Bioassays were carried out on H. armigera by diet incorporation method using 3 replicates having 10 insects taken with appropriate control. An aliquot of toxin solution was mixed with 10 g diet at different concentrations. The estimates of toxins were based up on their protein content. The treated diet was then divided into three replicates and fed to neonates separately. The insects were allowed to feed freely. The mortality of larvae was observed each day until 4 and 7 days. The mortality data were used for estimation of LC50 in terms of μg g-1 diet as per maximum likelihood programme. The solubilised protein preparation of recombinant strain ECE 53 (E. coli clone with cry1Ac1 gene) was used as control in bioassays.
RESULTS
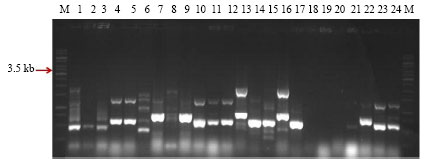
Screening of Bt isolates for the presence of full length cry1Aa,b,c-type genes: Sixty three native Bt isolates and 10 Bt reference strains were subjected to PCR amplification using specially designed primer set as per Beard et al. (2001) for the amplification of the full length cry1Aa,b,c-type genes. The prominent band of expected size of 3.6 kb was observed in native Bt isolate SK-711. Three Bt reference strains namely, B. thuringiensis subsp. taumanoffi (4N1), B. thuringiensis subsp. aizawai (4J2 and 4J4) were also found to be positive. These Bt strains are known to have full length cry1A-type genes. B. thuringiensis subsp. israelensis (4Q5) was used as a negative control as it doesn’t have full length cry1A-type genes. In addition, to band of expected size, some bands of small size were also observed in some isolates (Fig. 1a-c). Most of the remaining Bt isolates and Bt strains showed the variant bands of smaller size than expected and some didn’t showed any band.
Cloning of PCR amplified full length cry1A-type gene from native Bt isolate: PCR product of native Bt isolate SK-711 amplified using the set of primers designed as per Beard et al. (2002) was purified from the agarose gel matrix using Qiagen Mini Elute gel extraction kit (Fig. 2).
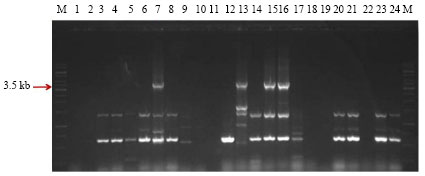 | |
| Fig. 1a: | PCR amplification of native Bt isolates and Bt strains used as reference with primers designed as per Beard et al. (2001). M: 1kb DNA Ladder; Lane 1: SK-930, Lane 2: SK-629, Lane 3: SK-722, Lane 4: SK-922, Lane 5: SK-851, Lane 6: SK-958, Lane 7: SK-711, Lane 8: SK-20, Lane 9: SK-305, Lane 10: SK-222, Lane 11: SK-1, Lane 12: SK-13, Lane 13: 4N1, Lane 14: 4A6, Lane 15: 4J2, Lane 16: 4J4, M: 1kb DNA Ladder, Lane 17: 4Q5, Lane 18: 4C3,Lane 19: 4M2, Lane 20: SK-977, Lane 21: SK-980, Lane 22: SK-995, Lane 23: SK-935 and Lane 24: SK-959 |
 | |
| Fig. 1b: | PCR amplification of native Bt isolates with primers designed as per Beard et al. (2001). M: 1kb DNA Ladder, Lane 1: SK-88, Lane 2: SK-4, Lane 3: SK-28, Lane 4: SK-232, Lane 5: SK-463, Lane 6: SK-957, Lane 7: SK-678, Lane 8: SK-754, Lane 9: SK-677, Lane 10: SK-921, Lane 11: SK-960, Lane 12: SK-942, Lane 13: SK-944, Lane 14: SK-304, Lane 15: SK-996, Lane 16: SK-9 M: 1kb DNA Ladder, Lane 17: SK-219, Lane 18: SK-617, Lane 19: SK-741, Lane 20: SK-953, Lane 21: SK-753, Lane 22: SK-449, Lane 23: SK-82 and Lane 24: SK-110 |
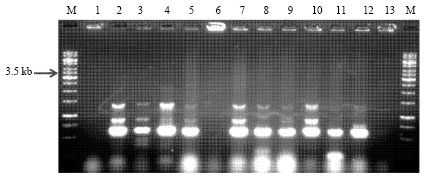 | |
| Fig. 1c: | PCR amplification of native Bt isolates using primers designed as per Beard et al. (2001). M: 1kb DNA Ladder, Lane 1: SK-792, Lane 2: SK-952, Lane 3: SK-962, Lane 4: SK-1007, Lane 5: SK-1008, Lane 6: SK-1009, Lane 7: SK-1025, Lane 8: SK-1026, Lane 9: SK-1027, Lane 10: SK-1028, Lane 11: SK-1034, Lane 12: SK-1036 and M: 1kb DNA Ladder |
It was cloned into pQE-30 UA expression cloning vector, which is 3.5 kb in size. Seven colonies were observed on LA plates containing ampicillin (100 μg mL-1). Molecular confirmation of recombinant clones was carried out by restriction analysis and PCR amplification. Plasmid DNA of each colony was isolated and examined for their size by agarose gel electrophoresis (Fig. 3). One recombinant clone (711-D) showed a band of about 8.0 kb while the rest showed a band of about 2.5 kb. Recombinant plasmid DNA of clone 711-D was digested with BamH1 restriction enzyme and as expected, a single band of size 7.1 kb corresponding to linearized vector was observed.
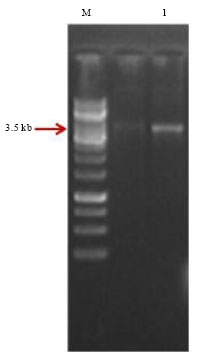 | |
| Fig. 2: | Gel purification of PCR product of native Bt isolate SK-711. M: 1 kb DNA Ladder, Lane 1: Purified PCR product |
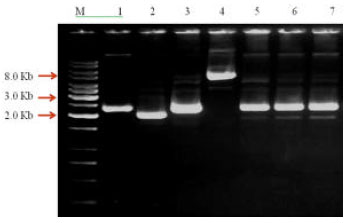 | |
| Fig. 3: | Gel electrophoresis of uncut recombinant plasmid DNA. M: 1 kb DNA Ladder, Lane 1: 711-A, Lane 2: 711-B, Lane 3: 711-C, Lane 4: 711-D, Lane 5: 711-E, Lane 6: 711-F, Lane 7: 711-G |
The recombinant clone was further confirmed by double digestion using BamH1 and Sal1 restriction enzymes for excising the insert of size 3.6 kb. Two bands of size 3.5 and 3.6 kb corresponding to vector and insert, respectively were observed (Fig. 4). PCR amplification of recombinant plasmid DNA of clone 711-D and of plasmid DNA of native Bt isolate SK-711 was carried out, using a set of primers designed as per Beard et al. (2001) to confirm the presence of desired insert in recombinant vector DNA. All the reaction conditions were kept same as in the previous PCR reaction through which Bt isolates were screened by this set of primers. The prominent band of 3.6 kb as expected was observed with both the native Bt isolate SK-711 as well as the recombinant clone 711-D (Fig. 5). The recombinant clone was named as pRKM-711.
Sequence determination and analysis: The sequence of the cry gene cloned in the recombinant clone pRKM 711 was determined by primer walking. Homology search and analysis was done by using BLAST programme of NCBI (National Centre for Biotechnology Information) web site: http://www.ncbi.nlm.nih.gov.). The sequence of the cloned gene was deposited in the NCBI GenBank (accession No. GQ866913). The sequence was analyzed by comparison with previously known cry1A-type genes and was assigned the name cry1Ac33 by the Bacillus thuringiensis Nomenclature Committee (http://www.biols.susx.ac.uk/Home/Neil_Crickmore/Bt/. The sequence was further analyzed for comparison with all the previously reported cry1Ac-type genes downloaded from the site http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/. using Multalin (version 5.4.1) web-based tool for multiple alignments. The gene sequence showed high level of homology with previously reported cry1Ac genes.
Expression of recombinant clone pRKM-711 in E. coli: The gene was expressed in E. coli under IPTG induction. The expected band of 130 kDa was observed in SDS-PAGE with Coomassie blue staining (Fig. 6).
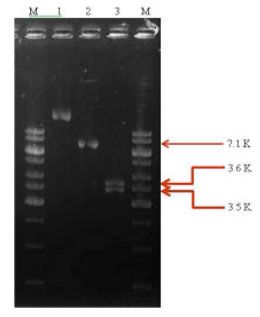 | |
| Fig. 4: | Restriction analysis of recombinant clone pRKM-711. M: 1 kb DNA Ladder, Lane 1: Uncut plasmid, Lane 2: Single digestion with BamH1, Lane 3: Double digestion with BamH1+Sal1, M: 1 kb DNA Ladder |
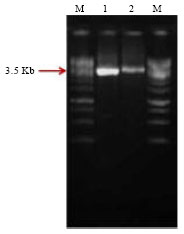 | |
| Fig. 5: | Confirmation of recombinant clone pRKM-711 by PCR amplification by PCR amplification with primers designed as per Beard et al. (2001). M: 1 kb DNA Ladder, Lane 1: PCR product with pRKM-711 as template, Lane 2: PCR product with native Bt isolate SK 711 as template, M: 1 kb DNA Ladder |
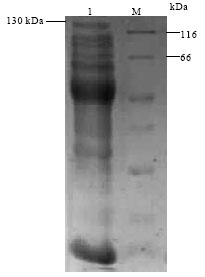 | |
| Fig. 6: | SDS-PAGE analysis of recombinant clone pRKM-711 with a cry1Ac-type gene in pQE-30 UA vector expressed in E. coli. Lane 1: Recombinant clone pRKM-711; M: Protein Marker. |
Evaluation for toxicity towards H. armigera: The solubilised protein preparation was evaluated for toxicity towards H. armigera (Table 4). It was observed that the cry1Ac33 gene was more toxic towards H. armigera than the holotype cry1Ac1 used as a control, based on LC50 toxicity analysis (Table 5, 6).
DISCUSSION
Several molecular techniques have been employed for detection of the cry genes out of which the identification of Bt cry genes by Polymerase Chain Reaction (PCR) has proven to be a very useful and rapid method for strain characterization and selection (Porcar and Juarez-Perez, 2003; Kaur, 2006).
| Table 5: | Evaluation of toxicity of solubilized liquid Cry1Ac33 protein (code: NRCPB RKM) against H. armigera* |
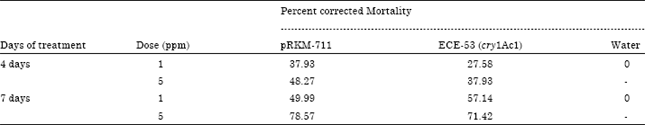 | |
| * 30 insects were used in three replicates for each assay | |
| Table 6: | Toxicity of Cry toxins to the neonates of H. armigera |
 | |
PCR and its variants have been widely used for detection of cry genes in collection of native Bt isolates since last decade, worldwide (Bravo et al., 1998; Beard et al., 2001; Choi et al., 2007; Thaphan et al., 2008; Thammasittirong and Attathom, 2008; Zhu et al., 2009). The genes of cry1 family are specifically toxic to insects of order Lepidoptera which include some major devastating pests of economically important crops and are responsible for most of the crop losses.
PCR screening of native Bt isolates for the presence of full length cry1Aa,b,c-type genes: Sixty three native Bt isolates and 10 Bt strains used as reference were screened using specially designed primers for the amplification of full length cry1Aa,b,c-type gene(s). Cry1A toxins are very important because of their high toxicity to lepidopteran pests and widespread distribution among Bt strains (Uribe et al., 2003; Wang et al., 2003; Stobdan et al., 2004; Beron et al., 2005; Kaur, 2006).
The cry1Aa, cry1Ab and cry1Ac genes are the most frequently found in Bt strains and native isolates (Ben-Dov et al., 1997; Bravo et al., 1998; Uribe et al., 2003; Stobdan et al., 2004; Armengol et al., 2007; Ramalakshmi and Udayasuriyan, 2010; Shankar et al., 2010). In order to obtain whole open reading frame of cry1A-type gene(s) for practical utility, amplification and cloning of full length cry1A-type genes was attempted by screening of 63 native Bt isolates with 10 reference Bt strains using specially designed primer sets as per Beard et al. (2001) in PCR amplification. These primer sets are specifically designed for the amplification of full length cry1A-type genes through the analysis of conserved regions by multiple alignments of all nucleotide sequences of cry1A-type genes available in databases. The prominent band of expected size 3.6 kb was observed in the native Bt isolate SK-711 recovered from Red gram field, Lam, Guntur Andhra Pradesh. In addition, to this native Bt isolate, 3 Bt strains namely, B. thuringiensis subsp. taumanoffi (4N1), B. thuringiensis subsp. aizawai (4J2 and 4J4) used as reference, were also found to be positive with the band of expected size along with some variant bands of small size. These three Bt strains are known to harbour cry1A-type genes on their plasmids (http://bacillus.biosci.ohio-state.edu/). The appearance of unexpected bands may imply that the primers may have recognized related cry genes sequences. Some of these bands may also be due to favoured amplification of shorter region of homology. It is also possible that some of the Bt isolates may harbour the cry1A-type genes in the truncated form such that primer recognition region is not present. Such a situation may seem conceivable due to frequent recombination events in cry genes helped by flanking transposons (Lereclus and Arantes, 1992).
Cloning of full length gene amplified by primer set designed as per Beard et al. (2001): The fragment of 3.6 kb amplified from native Bt isolate SK-711 by using primer set designed as per Beard et al. (2001) was cloned into pQE-30 UA expression cloning vector and transformed into competent Escherichia coli strain XL-1 Blue cells. All the colonies grown on LA plates containing ampicillin were selected as the vector lacks any other selection marker apart from the ampicillin resistance for the selection of colonies having recombinant vector. One recombinant clone (pRKM-711-D) was selected out of seven on the basis of plasmid DNA profile and was further confirmed by restriction analysis as well as by PCR amplification of recombinant plasmid DNA using the same set of primer designed as per Beard et al. (2001) as used for amplification of plasmid DNA of Bt isolate SK-711.
Sequence determination and analysis of cloned full length gene: Sequence similarity of the cloned cry1Ac33 gene with previously reported cry1Ac genes was observed. The nucleotide sequence of the cloned gene was almost 99% identical to all other cry1Ac genes except some differences of gap region and substitution of a particular nucleotide at specific positions. The nucleotide sequence corresponding to C-terminal of the Cry protein showed more than 50% homology with all the previously reported cry1Ac-type genes. The C-terminus portion of the protoxin is cleaved during proteolysis in the midgut of insects by proteases. The cry1A-type genes typically range from 3.1 to 3.6 kb. Variation in toxicity and specificity due to minor amino acid substitutions exist among different Cry1A toxins (Tounsi et al., 1999). Sequences of 62 cry1A genes, classified into cry1Aa to cry1Ai types, have been published (Crickmore et al., 1998).
The sequence of the cloned cry1Ac33 gene was also found to be almost 99% identical to the previously isolated cry1Ac18 gene (http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/, GenBank accession No. DQ023296) from the native Bt isolate SK-729 recovered from cotton seed, var.: LK-861, Lam, Guntur Andhra Pradesh in our laboratory (Kaur and Allam, 2006). The sequence was also 95% identical to the previously isolated cry1Ab18 gene (http://www.lifesci.sussex.ac.uk/ home/Neil_Crickmore/Bt/, GenBank accession No. AY319967) from the Bt isolate recovered from Ladakh and from native Bt isolate SK-222 (GenBank accession No. DQ023297) recovered from Pea phyllosphere in our laboratory (Stobdan et al., 2004). These genes had been cloned from native Bt isolates using two sets of specially designed ORF primers corresponding to full-length cry1Aa,b,c-type genes (Stobdan et al., 2004). In this study, the set of primers designed as per Beard et al. (2001) was employed.
Expression in E. coli and evaluation for toxicity towards H. armigera: The development of resistance in target insects towards Cry toxins could diminish the long-term efficacy of Bt crops (Tabashnik, 2008). To delay the onset of resistance in insects towards the known genes, one or more different types of cry genes can be used in gene pyramiding strategy. Therefore, the search for the cry genes by screening of native Bt isolates is of much interest for deployment of these genes in transgenic crops.
The cry1Aa, cry1Ab and cry1Ac genes have been reported to be toxic to lepidopteran pests, particularly, H. armigera, a severe and polyphagous pest of several economically important crops such as cotton, pigeon pea, chickpea and tomato (Zafar et al., 2000; Khalique, 2003; Gupta et al., 2007). Of all the cry genes, the cry1Ac genes are most toxic to H. armigera (Padidam, 1992; Kumar et al., 2004).
The cry1Ac33 gene was expressed in E. coli and evaluated for toxicity towards H. armigera and has been found to be more toxic towards H. armigera than the holotype cry1Ac1 used as a control, based on LC50 toxicity analysis. The differences in nucleotide and derived amino acid sequence of cry1Ac33 gene from that of cry1Ac1 gene may be involved in observed higher toxicity of this gene. To tackle the imminent problem of development of resistance in target insect pests towards cry genes currently deployed in transgenic plants, novel insecticidal genes are required (Kaur, 2007). This gene can, thus, be useful for deployment for insect control.
ACKNOWLEDGMENTS
Authors acknowledge R.K. Narula for technical assistance and Department of Biotechnology for financial assistance. This work was part of M.Sc. Thesis of RKM.
REFERENCES
- Ali, B.A., H.H. Salem, X.M. Wang, T.H. Huang, Q.D. Xie and X.Y. Zhang, 2010. Effect of Bacillus thuringiensis var. Israelensis endotoxin on the intermediate snail host of Schistosoma japonicum. Curr. Res. Bacteriol., 3: 37-41.
CrossRefDirect Link - Amjad, M., M. Afzal and K. Mehmood, 2001. New synthetic and bio-insecticides against maize stem borer, Chilo partellus (Swinhoe) on golden maize. J. Biological Sci., 1: 38-39.
CrossRefDirect Link - Anwar, R., S. Karim and S. Riazuddin, 1999. Entomocidal activity of Bacillus Thuringiensis transgenic rice plants against rice leaffolder, Cnaphalocrocis Medinalis (Lepidoptera: Pyralidae). Pak. J. Biol. Sci., 2: 1472-1477.
CrossRefDirect Link - Apaydin, O., A.F. Yenidunya, S. Harsa and H. Gunes, 2005. Isolation and characterization of Bacillus thuringiensis strains from different grain habitats in Turkey. World J. Microbiol. Biotechnol., 21: 285-292.
Direct Link - Armengol, G., M.C. Escobar, M.E. Maldonado and S. Orduz, 2007. Diversity of Colombian strains of Bacillus thuringiensis with insecticidal activity against dipteran and lepidopteran insects. J. Applied Microbiol., 102: 77-88.
PubMed - Barton, K.A., H.R. Whiteley and N.S. Yang, 1987. Bacillus thuringiensis δ-endotoxin in transgenic Nicotiana tabacum provides resistance to lepidopteran insects. Plant Physiol., 85: 1103-1109.
Direct Link - Beard, C.E., C. Ranasinghe and R.J. Akhurst, 2001. Screening for novel cry genes by hybridization. Lett. Applied Microbiol., 33: 241-245.
Direct Link - Ben-Dov, E., A. Zaritsky, E. Dahan, Z. Barak and R. Sinai et al., 1997. Extended screening by PCR for seven cry-group genes from field-collected strains of Bacillus thuringiensis. Applied Environ. Microbiol., 63: 4883-4890.
Direct Link - Bernhard, K., P. Jarrett, M. Meadows, J. Butt and D.J. Ellis et al, 1997. Natural isolates of Bacillus thuringiensis: Worldwide distribution, characterization and activity against insect pests. J. Inverteb. Pathol., 70: 59-68.
CrossRefDirect Link - Beron, C.M., L. Curatti and G.L. Salerno, 2005. New strategy for identification of novel cry-type genes from Bacillus thuringiensis strains. Applied Environ. Microbiol., 71: 761-765.
Direct Link - Bravo, A., S. Sarabia, L. Lopez, H. Ontiveros and C. Abarca et al, 1998. Characterization of Cry genes in a mexican Bacillus thuringiensis strain collection. Applied Environ. Microbiol., 64: 4965-4972.
Direct Link - Brookes, G. and P. Barfoot, 2008. Global impact of biotech crops: Socio-economic and environmental effects, 1996-2006. AgBio Forum, 11: 21-38.
Direct Link - Carpenter, J.E., 2010. Peer-reviewed surveys indicate positive impact of commercialized GM crops. Nat. Biotechnol., 28: 319-321.
CrossRefDirect Link - Choi, J.Y., M.S. Li, H.J. Shim, J.Y. Roh and S.D. Woo, et al., 2007. Isolation and characterization of strain of Bacillus thuringiensis subsp: Kenyae containing two novel Cry1-type toxin genes. J. Microbiol Biotechnol., 17: 1498-1503.
PubMed - Choudhary, B. and K. Gaur, 2010. Bt Cotton in India: A Country Profile. The International Service for the Acquisitions of Agri-biotech Applications (ISAAA), Ithaca, NY., USA., ISBN: 978-1-892456-46-X, Pages: 32.
Direct Link - Crickmore, N., D.R. Zeigler, J. Feitelson, E. Schnepf and J. Van Rie et al., 1998. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev., 62: 807-813.
Direct Link - De Escudero, R., A. Estela, M. Porcar, C. Martinez and J.A. Oguiza et al., 2006. Molecular and insecticidal characterization of a Cry1I protein toxic to insects of the families Noctuidae. Tortricidae, Plutellidae and Chrysomelidae. Applied Environ. Microbiol., 72: 4796-4804.
PubMed - Downes, S., T. Parker and R. Mahon, 2010. Incipient resistance of Helicoverpa punctigera to the cry2Ab Bt toxin in bollgard II® cotton. PLoS One, Vol. 5.
CrossRefDirect Link - Garcia-Robles, I., J. Sanchez, A. Gruppe, A.C. Martinez-Ramirez, C. Rausell, M.D. Real and A. Bravo, 2001. Mode of action of Bacillus thuringiensis PS86Q3 strain in hymenopteran forest pests. Insect Biochem. Mol. Biol., 31: 849-856.
CrossRefDirect Link - Gatehouse, J.A., 2008. Biotechnological prospects for engineering insect-resistant plants. Plant Physiol., 146: 881-887.
Direct Link - Gupta, R.K., J.C. Raina, R.K. Arora and K. Bali, 2007. Selection and field effectiveness of nucleopolyhedrovirus isolates against Helicoverpa armigera (Hubner). Int. J. Virol., 3: 45-59.
CrossRefDirect Link - Hansen, B.M., P.H. Damgaard, J. Eilenberg and J.C. Pedersen, 1998. Molecular and phenotypic characterization of Bacillus thuringiensis isolated from leaves and insects. J. Invertebrate Pathol., 71: 106-114.
CrossRef - Jara, S., P. Maduell and S. Orduz, 2006. Diversity of Bacillus thuringiensis strains in the maize and bean phylloplane and their respective soils in Colombia. J. Appl Microbiol., 101: 117-124.
PubMed - Karim, S., M. Murtaza and S. Riazuddin, 1999. Field evaluation of bacillus thuringiensis, insect growth regulators, chemical pesticide against Helicoverpa armigera (Huber) (Lepidoptera: Noctuidae) and their compatibility for integrated pest management. Pak. J. Biol. Sci., 2: 320-326.
CrossRefDirect Link - Kaur, S., 2006. Molecular approaches for identification and construction of novel insecticidal genes for crop protection. World J. MIcrobiol. Biotechnol., 22: 233-253.
CrossRefDirect Link - Kaur, S. and A. Singh, 2000. Natural occurrence of Bacillus thuringiensis in leguminous phylloplanes in the New Delhi region of India. World J. Microbiol. Biotech., 16: 679-682.
CrossRefDirect Link - Khalique, F. and A. Khalique, 2003. Impact of Bacillus thuringiensis Subsp. kurstaki on biology of Helicoverpa armigera. Pak. J. Biol. Sci., 6: 615-621.
CrossRefDirect Link - Khan, M.A., R. Makhdoom, T. Husnain, M.Z. Saleem, K. Malik, Z. Latif, I. Altosaar and S. Riazuddin, 2001. Expression of Bt gene in a dicot plant under promoter derived from a monocot plant. Pak. J. Biol. Sci., 4: 1518-1522.
CrossRefDirect Link - Kumar, S., V. Udaisuriyan, P. Sangeetha and M. Bharathi, 2004. Analysis of Cry2A proteins encoded by genes cloned from indigenous isolates of Bacillus thuringiensis for toxicity against Helicoverpa armigera. Curr. Sci., 86: 566-570.
Direct Link - Lee, M.K., R.E. Milne, A.Z. Ge and D.H. Dean, 1992. Location of a Bombyx mori receptor binding region on a Bacillus thuringiensis δ-endotoxin. J. Biol. Chem., 267: 3115-3121.
Direct Link - Lereclus, D. and O. Arantes, 1992. SpbA locus ensures the segregational stability of pTH1030, a novel type of gram-positive replicon. Mol. Microbiol., 6: 35-46.
PubMed - Martinez, C. and P. Caballero, 2002. Contents of Cry genes and insecticidal toxicity of Bacillus thuringiensis strains from terrestrial and aquatic habitats. J. Applied Microbiol., 92: 745-752.
PubMed - Meadows, M.P., D.J. Ellis, J. Butt, P. Jarrett and H.D. Burges, 1992. Distribution, frequency and diversity of Bacillus thuringiensis in an animal feed mill. Applied Environ. Microbiol., 58: 1344-1350.
Direct Link - Porcar, M. and V. Juarez-Perez, 2003. PCR based identification of Bacillus thuringiensis pesticidal crystal genes. FEMS Microbiol. Rev., 26: 419-432.
Direct Link - Ramalakshmi, A. and V. Udayasuriyan, 2010. Diversity of Bacillus thuringiensis isolated from Western Ghats of Tamil Nadu State, India. Curr. Microbiol., 61: 13-18.
PubMed - Halima, H.S., A.A. Bahy, H.H. Tian and D.X. Qing, 2006. Molecular characterization of novel Bacillus thuringiensis isolate with molluscicidal activity against the intermediate host of schistosomes. Biotechnology, 5: 413-420.
CrossRefDirect Link - Schnepf, E., N. Crickmore, J. van Rie, D. Lereclus and J. Baum et al., 1998. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev., 62: 775-806.
Direct Link - Stobdan, T., S. Kaur and A. Singh, 2004. Cloning and nucleotide sequence of a novel Cry gene from Bacillus thuringiensis. Biotechnol. Lett., 26: 1153-1156.
PubMed - Subramanian, S. and S. Mohankumar, 2006. Genetic variability of the bollworm, Helicoverpa armigera, occurring on different host plants. J. Insect Sci., Vol. 6, No. 1.
CrossRefDirect Link - Swiecicka, I., D.K. Bideshi and B.A. Federici, 2008. Novel isolate of Bacillus thuringiensis subsp. thuringiensis that produces a quasicuboidal crystal of Cry1Ab21 toxic to larvae of Trichoplusia ni. Applied Environ. Microbiol., 74: 923-930.
Direct Link - Tabashnik, B.E., 2008. Delaying insect resistance to transgenic crops. Proc. Natl. Acad. Sci., 105: 19029-19030.
CrossRefDirect Link - Thammasittirong, A. and T. Attathom, 2008. PCR-based method for the detection of cry genes in local isolates of Bacillus thuringiensis from Thailand. J. Invertebr. Pathol., 98: 121-126.
PubMed - Thaphan, P., S. Keawsompong and J. Chanpaisaeng, 2008. Isolation, toxicity and detection of cry gene in Bacillus thuringiensis isolates in Krabi province, Thailand. Songklanakarin. J. Sci. Technol., 30: 597-601.
Direct Link - Uribe, D., W. Martinez and J. Ceron, 2003. Distribution and diversity of cry genes in native strains of Bacillus thuringiensis obtained from different ecosystems from Colombia. J. Invertebr. Pathol., 82: 119-127.
PubMed - Wang, J., A. Boets, J. van Rie and G. Ren, 2003. Characterization of cry1, cry2 and cry9 genes in Bacillus thuringiensis isolates from China. J. Inverteb. Pathol., 82: 63-71.
CrossRefDirect Link - Wu, K., Y. Guo, N. Lv, J.T. Greenplate and R. Deaton, 2003. Efficacy of transgenic cotton containing a Cry1Ac gene from Bacillus thuringiensis against Helicoverpa armigera (Lepidoptera: Noctuidae) in Northern China. J. Eco. Entomol., 96: 1322-1328.
CrossRefDirect Link - Yang, Z., H. Chen, W. Tang, H. Hua and Y. Lin, 2011. Development and characterisation of transgenic rice expressing two Bacillus thuringiensis genes. Pest Manage. Sci., 67: 414-422.
CrossRefDirect Link - Zaied, K.A., Z.M. El-Diasty, S.K. Kawther and M.E. Ismail, 2003. Sensitivity of lepidopterous larvae to recombinant products of Bacillus thuringiensis as a bioinsecticide agent. Pak. J. Biol. Sci., 6: 273-313.
CrossRefDirect Link - Zafar, A.U., S. Karim, I.A. Nasir and S. Riazuddin, 2000. Shelf life and field evaluation of CAMB Bacillus thuringiensis biopesticide against Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) on tomato. Pak. J. Biol. Sci., 3: 804-807.
CrossRefDirect Link - Zia-ur-Rehman, M., A.U. Zafar, I.A. Nasir and S. Riazuddin, 2002. Comparative study of Bacillus thuringiensis biopesticides against cotton bollworms. Asian J. Plant Sci., 1: 574-576.
CrossRefDirect Link - Zhao, J.Z., J. Cao, Y. Li, H.L. Collins, R.T. Roush, E.D. Earle and A.M. Shelton, 2003. Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nat. Biotechnol., 21: 1493-1497.
PubMed - Zhu, J., F. Tan, J. Tang, Y. Li, A. Zheng and P. Li, 2009. Characterization of insecticidal crystal proteincry gene of Bacillus thuringiensis from soil of Sichuan Basin, China and cloning of novel haplotypes Cry gene. Annals Microbiol., 59: 1-8.
CrossRefDirect Link - Kalman, S., K.L. Kiehne, J.L. Libs and T. Yamamoto, 1993. Cloning of a novel cryIC type gene from a strain of Bacillus thuringiensis subsp. Galleriae. Applied Environ. Microbiol., 59: 1131-1137.
Direct Link - Tounsi, S., A. J'Mal, N. Zouari and S. Jaoua, 1999. Cloning and nucleotide sequence of a novel cry1Aα-type gene from Bacillus thuringiensis subsp. Kurstaki. Biotechnol. Lett., 21: 771-775.
Direct Link