
Pawitra Pulbutr
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy, Mahasarakham University, Mahasarakham, 44150, Thailand
Atika Jaruchotikamol
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy, Mahasarakham University, Mahasarakham, 44150, Thailand
Jirawan Chaiyachart
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy, Mahasarakham University, Mahasarakham, 44150, Thailand
Pannee Sonrasee
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy, Mahasarakham University, Mahasarakham, 44150, Thailand
Wannaporn Poungpan
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy, Mahasarakham University, Mahasarakham, 44150, Thailand
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 5 | Page No.: 294-300







ABSTRACT
(-)-Epigallocatechin-3-gallate (EGCG), the major catechins found in green tea, has been known to possess various pharmacological activities. It has been demonstrated that EGCG affects on several adipocyte functions, involving in its weight-controlling action. This study was aimed to investigate the effects of EGCG on adipocyte lipolysis in adipocytes derived from Normal Pellet Diet (NPD)-fed rats and High Fat Diet (HFD)-fed rats. Sixteen male Wistar rats were divided into 2 groups (8 rats per group). The rats in each group were fed with NPD or HFD for 3 weeks. The adipocyte suspensions were prepared by the collagenase digestion method. The concentration of Free Fatty Acid (FFA) was measured after 1 h-incubation as an index of adipocyte lipolysis. EGCG at every concentration tested (1, 5 and 10 μM) significantly stimulated basal lipolysis in adipocytes derived from the NPD-fed rats with the FFA concentrations of 453.67±25.57, 458.87±26.84 and 443.97±19.75 μM mL-1 Packed Cell Volume (PCV)/h, respectively (p<0.05). In adipocytes derived from the HFD-fed rat, EGCG only at the concentration of 5 μM significantly increased basal adipocyte lipolysis with the FFA concentration of 1,091.75±220.56 μM mL-1 PCV h-1 (p<0.05; n = 4). Conversely, EGCG at any of the concentrations tested did not cause a significant change in isoprenaline (0.1 μM)-induced lipolysis in both adipocyte groups. These results indicate a lower lipolytic action of EGCG in adipocytes derived from the HFD-fed rats. Further experiments thus should be done to explore the reason for the difference in the lipolytic response to EGCG between these adipocytes.
PDF Abstract XML References Citation
Received: June 07, 2012;
Accepted: July 24, 2012;
Published: September 06, 2012
How to cite this article
Pawitra Pulbutr, Atika Jaruchotikamol, Jirawan Chaiyachart, Pannee Sonrasee and Wannaporn Poungpan, 2012. Different Lipolytic Response to (-)-Epigallocatechin-3-gallate in Adipocytes Derived
from Normal Diet-fed Rats and High Fat Diet-fed Rats. Journal of Biological Sciences, 12: 294-300.
DOI: 10.3923/jbs.2012.294.300
URL: https://scialert.net/abstract/?doi=jbs.2012.294.300
DOI: 10.3923/jbs.2012.294.300
URL: https://scialert.net/abstract/?doi=jbs.2012.294.300
INTRODUCTION
Obesity which is now considered as a global public health problem, has significant impacts on various aspects, including physical and mental health, quality of life and also economy (Luppino et al., 2010; Cash et al., 2011; Von Lengerke and Krauth, 2011). Although, obesity is a multifactorial disease, over food consumption and lack of physical activity are known as the major causes of the disease (Biro and Wien, 2010). Obesity is an established risk factor for several chronic disorders such as hypertension, dyslipidemia, type 2 diabetes and some forms of cancers (Kopelman, 2000). Thus, prevention of obesity is the plausibly effective way to protect people from aforementioned diseases. Adipocyte lipolysis which is characterized by hydrolysis of triglyceride storage in adipocytes, plays a crucial role in the regulation of fat mass (Arner, 2005). Modulation of adipocyte lipolysis thus provides a potential way to reduce fat mass and alleviate obesity and related disorders.
Since there are some limitations of conventional weight controlling programs, pharmacologically active compounds derived from plants have been highly focused as an alternative means to manage obesity. One of the most promising compounds is green tea-derived (-)-epigallocatechin-3-gallate (EGCG). Green tea which is obtained from the leaves of Camellia sinensis, is a widely consumed oriental beverage. EGCG accounts for approximately 50% of the total amount of catechins found in green tea (Balentine et al., 1997). EGCG has been found to play a key role in various health-related benefits of green tea, including its weight controlling action (Lee et al., 2009a; Thielecke and Boschmann, 2009; Chen et al., 2011). EGCG has been reported to reduce fat mass by controlling various adipocyte functions, including suppression of adipogenesis, inhibition of adipocyte lipogenesis and stimulation of adipocyte lipolysis (Chan et al., 2011; Sung et al., 2010; Lee et al., 2009a; Ogasawara et al., 2011). However, the lipolytic effect of EGCG has not been established in adipocytes derived from animals fed with high fat diet which is a well-known model of diet-induced obesity. This study was aimed to investigate the effects of EGCG on adipocyte lipolysis in isolated adipocytes derived from high fat diet-fed rats and to compare its actions to those found in adipocytes derived from normal diet-fed rats.
MATERIALS AND METHODS
Preparation of adipocytes: Sixteen male Wistar rats weighing 150-170 g were obtained from the National Laboratory Animal Center, Mahidol University, Thailand. The animals were kept at constant temperature (25±1°C) with a 12 h dark-light cycle. After acclimatization period of one week, the rats were randomly divided into two groups. The rats in the first group (n = 8) were fed with Normal Pellet Diet (NPD) (C.P. mice feed food No. 082, Bangkok, Thailand) (NPD-group), whilst the rats in the second group (n = 8) were fed with high fat diet (HFD-group). The fat content in NPD and HFD are approximately 12 and 56% of total energy, respectively. High fat diet was prepared according to the formulation of Srinivasan et al. (2005) with slight modification. The animals were fed ad libitum and freely accessed to water. After three weeks, the rats were sacrificed by CO2 overdose and epididymal fat pads were dissected for preparation of adipocyte suspension. All procedures performed with the animals were approved by the animal research ethic committee, Mahasarakham University, Thailand.
Adipocytes were isolated according to the method of Rodbell (1964) with some modifications. Briefly, the epididymal fat pads were rinsed with Hanks buffer containing 5 mM glucose and 0.5% bovine serum albumin (Sigma) (BSA, with less than 0.005% (w/w) free fatty acid). Hanks physiological solution containing (in mM): 5.6 KCl, 138 NaCl, 4.2 NaHCO3, 1.2 NaH2PO4, 2.6 CaCl2, 1.2 MgCl2, 10 HEPES, pH 7.4 (with NaOH). The tissues were cut into small pieces and then digested with 0.25 mg mL-1 collagenase type II enzyme (Sigma) at 37°C. Isolated adipocytes were filtered through nylon mesh and then rinsed twice with collagenase free Hanks buffer. After the second wash, the adipocytes were resuspended with Hanks solution into 1:1 (v/v) proportion to give a final volume of the cell suspension at about 8-15 mL, depending on the volume of adipocytes. The cell suspension was transferred to a fresh Nalgene conical flask and stored in the water bath at 37°C before performing experiments. All experiments were conducted at the Faculty of Pharmacy, Mahasarakham University, Thailand during 18th November 2011- 25th December 2011.
Adipocyte lipolysis: Adipocytes were incubated for 60 min in microcentrifuge tubes at 37°C with Hanks buffer, pH 7.4, containing 5 mM glucose, 0.5% BSA and various concentrations of (-)-Epigallocatechin-3-gallate (EGCG) (Sigma) at 1, 5 and 10 μM. For stimulated lipolysis study, the experiments were performed in the presence of isoprenaline (Sigma) at the concentration of 0.1 μM. The final volume of the incubations was adjusted to 1 mL. After 1 h incubation, the cell free incubation media were collected and the concentrations of Free Fatty Acid (FFA) were measured by using non-esterified fatty acid assay kit from WAKO Chemicals (Japan).
Statistical analysis: The results are expressed as Mean±SEM. Statistical analysis was performed by using either independent-sample Student’s t-test or one-way analysis of variance (ANOVA) followed by Bonferroni post hoc test. Statistical tests were performed using SPSS software version 16.0 (SPSS Inc., Chicago, Illinois, USA). The data was considered as significantly different when p-value<0.05.
RESULTS
Body weight and epididymal fat pad weight: The body weight of the rats was measured at the beginning of the experiment (day 0) and every week thereafter (day 7, 14, 21 and 28). The body weight of the rats fed with NPD was not different from that of the rats fed with HFD (Table 1). However, the epididymal fat pad weight of the HFD-fed rats (2.03±0.13 g/100 g of body weight, n = 8) was significantly higher than that of the NPD-fed control rats (1.48±0.85 g/100 g of body weight, n = 8) with the p-value of <0.05 (Table 2).
Lipolytic effects of EGCG in adipocytes derived from the NPD-fed rats (NPD-group): At the basal condition, the FFA concentration derived from adipocytes in the NPD group was 323.46±28.32 μM mL-1 PCV h-1 (Fig. 1).
| Table 1: | Body weight of NPD-fed rats and HFD-fed rats |
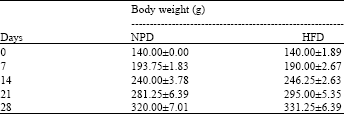 | |
| Values are Mean±SEM, n = 8, There is no statistically significant difference between groups. The statistical analysis was performed by using independent-sample Student’s t-test | |
| Table 2: | Weight of epididymal fat pads of NPD-fed rats and HFD-fed rats |
| *p<0.05 when compared with the NPD group (independent-sample Student’s t-test), Values are Mean±SEM, n = 8 | |
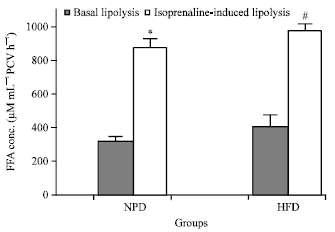 | |
| Fig. 1: | Basal lipolysis and isoprenaline (0.1 μM)-induced lipolysis in adipocytes derived from the NPD-group and the HFD-group. The results are expressed as the Mean±SEM (n = 5). *p<0.05 when compared with basal lipolysis in the NPD group; #p<0.05 when compared with basal lipolysis in the HFD group (independent-sample Student’s t-test) |
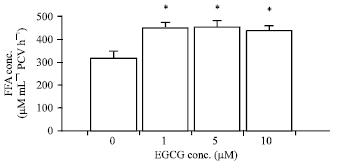 | |
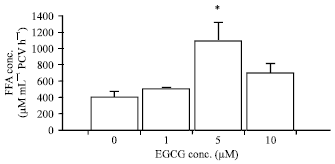
| Fig. 2: | Effects of EGCG on basal adipocyte lipolysis in the NPD-group. The results are expressed as the Mean±SEM (n = 5). *p<0.05 when compared with basal lipolysis (One-way ANOVA followed by Bonferroni post hoc test) |
EGCG at the concentrations of 1, 5 and 10 μM significantly stimulated basal adipocyte lipolysis in the NPD group with the FFA concentrations of 453.67±25.57, 458.87±26.84 and 443.97±19.75 μM mL-1 PCV/ h, respectively (p<0.05; n = 5) (Fig. 2).
When isoprenaline (0.1 μM) was added, the FFA concentration was significantly increased to the level of 877.57±52.69 μM mL-1 PCV h-1 (p<0.05) (Fig. 1). The FFA level in the presence of isoprenaline was calculated as 171.30±16.73% increase of that in the absence of isoprenaline (basal lipolysis). EGCG at any of the concentrations tested (1, 5, 10 μM) did not cause a significant change in isoprenaline-induced lipolysis in adipocytes derived from the NPD-fed rats.
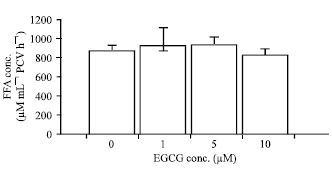 | |
| Fig. 3: | Effects of EGCG on isoprenaline-induced lipolysis in the NPD-group. The results are expressed as the Mean±SEM (n = 5). There is no statistically significant difference between groups. The statistical analysis was performed by using One-way ANOVA followed by Bonferroni post hoc test. Adipocytes were treated with 0.1 μM isoprenaline under the condition shown |
 | |
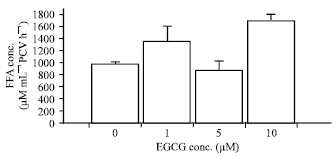
| Fig. 4: | Effects of EGCG on basal adipocyte lipolysis in the HFD-group. The results are expressed as the Mean±SEM (n = 4). *p<0.05 when compared with basal lipolysis (One-way ANOVA followed by Bonferroni post hoc test) |
In the condition of isoprenaline-induced lipolysis, the FFA concentrations in the presence of EGCG at the concentrations of 1, 5 and 10 μM were 929.50±180.09, 936.43±76.63 and 831.85±58.38 μM mL-1 PCV h-1, respectively (n = 5, Fig. 3).
Lipolytic effects of EGCG in adipocytes derived from the HFD-fed rats (HFD-group): The FFA concentration in the basal lipolysis of adipocytes derived from the HFD-fed rats was 407.49±70.42 μM mL-1 PCV h-1. This was not statistical dissimilar to the FFA concentration found in the basal lipolysis of adipocytes derived from the NPD-fed rats. EGCG at the concentration of 5 μM significantly increased the basal lipolysis in adipocytes derived from the HFD group with the FFA concentration of 1,091.75±220.56 μM mL-1 PCV h-1 (p<0.05; n = 4, Fig. 4). The basal adipocyte lipolysis was not significantly affected by the addition of EGCG at the other concentrations tested (1 and 10 μM).
 | |
| Fig. 5: | Effects of EGCG on isoprenaline-induced lipolysis in the HFD-group. The results are expressed as the Mean±SEM (n = 5). There is no statistically significant difference between groups. The statistical analysis was performed by using One-way ANOVA followed by Bonferroni post hoc test. Adipocytes were treated with 0.1 μM isoprenaline under the condition shown |
The FFA levels in the presence of EGCG at the concentrations of 1 and 10 μM were modestly increased to the concentrations of 514.28±9.11 and 706.48±107.89 μM mL-1 PCV h-1, respectively (n = 4). However, these levels did not reach a statistical significance.
Isoprenaline at the concentration of 0.1 μM significantly increased adipocyte lipolysis in adipocytes derived from the HFD group with the FFA concentration of 980.00±38.57 μM mL-1 PCV h-1 (p<0.05) (Fig. 1). In the presence of isoprenaline, the FFA concentration was increased by 140.51±9.46% of the basal lipolysis. EGCG at the concentrations of 1 and 10 μM caused an increase in isoprenaline-induced lipolysis, with the FFA concentrations of 1,356.16±262.97 and 1,699.70±110.94 μM mL-1 PCV h-1, respectively (n = 5). However, these FFA concentrations were not statistically different from that in the presence of isoprenaline alone. EGCG at the concentration of 5 μM did not cause a significant change in the isoprenaline-induced lipolysis, with the FFA concentration of 885.87±151.36 μM mL-1 PCV h-1 (n = 5, Fig. 5).
DISCUSSION
The body weight of the rats fed with HFD was not different from that of the rats fed with NPD. This is similar to the results of Harris (1991) and Collin et al. (2006) as well as our previous finding (Pulbutr et al., 2011). However, the weight of epididymal fat pads in the HFD-fed rats was significantly higher than that in the NPD-fed rats. The similar findings were reported by Naim et al. (1985) and Collin et al. (2006). From their studies, after feeding with high fat diet for 3 weeks, there was a significant increase in the weights of epididymal and visceral fat pads.
From this study, EGCG at every concentration tested (1, 5 and 10 μM) significantly stimulated basal adipocyte lipolysis in adipocytes isolated from the NPD-fed rats. This is in accordance with the results from the study of Lee et al. (2009a) which showed that 10 μM EGCG significantly increased basal adipocyte lipolysis. However, from their study the lipolytic stimulating action of EGCG was not detected when the lower concentration of 1 μM was used. Our experiments were performed by using primary rat adipocytes whilst 3T3-L1 adipocytes were used in the study of Lee et al. (2009a). The difference in adipocyte cell type may be the reason for the discrepancy in the lipolytic response to EGCG found between the studies. In addition, the lipolytic action of EGCG at the lower concentration (2.79 μM) was also demonstrated recently in primary rat adipocytes by Ogasawara et al. (2011).
The mRNA levels of lipolytic genes, including Hormone Sensitive Lipase (HSL) enzyme and Adipose Triglyceride Lipase (ATGL) enzyme were found to be increased in mice after feeding with high fat-diet supplemented with EGCG (0.2 or 0.5% w/w) for 8 weeks (Lee et al. 2009b). Additionally, it was shown from the same study that the mRNA levels of various adipogenic genes, such as PPAR-γ, CCAAT Enhancer-binding Protein-α (C/EBP-α), Sterol Regulatory Element-binding Protein-1c (SREBP-1c), adipocyte fatty acid-binding protein (aP2), Lipoprotein Lipase (LPL) and Fatty Acid Synthase (FAS), were also decreased in the mice fed with EGCG-supplemented high fat diet. Lee et al. (2009a) revealed that when treated with 10 μM EGCG for 24 h, the mRNA level of HSL was dramatically increased in 3T3-L1 adipocytes. The investigators thus concluded that EGCG increased adipocyte lipolysis via induction of HSL mRNA expression. However, in our study, the lipolytic stimulating action was demonstrated even when the adipocytes were treated with EGCG only for 1 h. It remains unanswered whether the lipolytic stimulating action of EGCG found here is also related to the changes in HSL mRNA expression.
It was reported recently that the lipolytic stimulating effect of EGCG was abolished when the adipocytes were pre-incubated with MAPK/ERK kinase 1/2 (MEK1/2) inhibitor (PD98059) (Ogasawara et al., 2011). From their study, the levels of ERK1/2 phosphorylation and MEK1/2 phosphorylation were also increased by EGCG treatment. These indicate that EGCG exerts its lipolytic action via the activation of ERK1/2 pathway which is another essential pathway for adipocyte lipolysis. Phosphorylation of HSL at Ser600 by ERK increases the activity of HSL and consequently stimulates adipocyte lipolysis (Greenberg et al., 2001).
EGCG did not produce its lipolytic stimulating action in adipocytes derived from the NPD-fed rats in the condition of isoprenaline-induced lipolysis. Isoprenaline was found to induce adipocyte lipolysis via both cAMP-dependent pathway and ERK1/2 pathway (Greenberg et al., 2001). It was reported that the lipolytic action of isoprenaline was depleted by approximately 30% when MEK inhibitors (PD98059 or U0126) were added simultaneously. It is possible that isoprenaline and EGCG may stimulate adipocyte lipolysis by using the similar ERK1/2 pathway. Thus, the lipolytic action of EGCG could not be observed in the presence of isoprenaline. Present results also imply that EGCG may not potentiate adipocyte lipolysis in the situations of sympathetic nervous system stimulation such as physical exercise and physical or psychological stress. However, EGCG also possesses other positive effects on lipid metabolism, such as inhibition of adipogenesis, inhibition of lipid accumulation and stimulation of lipid oxidation (Auvichayapat et al., 2008; Chan et al., 2011). These beneficial effects of EGCG may still exert during the sympathetic stimulation and the body weight controlling action of EGCG thus may still be observed.
Adipocytes derived from the HFD-fed rats had a lower response to EGCG in the condition of basal lipolysis. EGCG at every concentration tested (1, 5, 10 μM) produced a significant lipolytic action in adipocytes derived from the NPD-fed rats, but EGCG only at the concentration of 5 μM exhibited this action in adipocytes derived from the HFD-fed rats. This may be due to the difference in ERK pathway between adipocytes derived from the different diet type-fed rats. ERK phosphorylation was found to be increased in mice fed with high fat diet for 4, 6, 8 and 15 weeks (Ito et al., 2007). ERK phosphorylation and activation of MAPK pathway has been found to play an important role in adipocyte proliferation and differentiation. Bost et al. (2005) showed that mice lacking ERK1 (ERK1-/-mice) were resistant to diet-induced obesity and also protected from insulin resistance. The increased level of phosphorylated ERK in adipocytes derived from HFD-fed rodents is likely to be essential for adipocyte proliferation and differentiation. Most of the increased level of phosphorylated ERK may be shifted to the adipocyte proliferation and differentiation pathways, thus less amount of phosphorylated ERK would be available for the activation of ERK-related lipolytic pathway which involves in the lipolytic action of EGCG. This may partly be the reason why adipocytes derived from the HFD-fed rats had the lower basal lipolytic response to EGCG. Although ERK1/2 is activated early in the differentiation process, deactivation of ERK1/2 is needed during adipogenesis to enhance terminal differentiation (Gehart et al., 2010). According to this, it can be alternatively proposed that ERK pathway may be less activated in adipocytes derived from the HFD-fed rats, thus the lipolytic stimulating action of EGCG was lessened. However, further study is needed to prove these speculations. It is interesting that in adipocytes derived from the HFD-fed rats, EGCG at the concentration of 10 μM slightly increased basal lipolysis, but the level did not reach a significant difference. Further experiment thus should be performed with more various concentrations of EGCG to investigate whether EGCG performs its lipolytic-stimulating action in a concentration-dependent manner in HFD-derived adipocytes.
From this study, EGCG did not produce a significant lipolytic stimulating action in the presence of isoprenaline in adipocytes derived from both the NPD-fed rats and the HFD-fed rats. However, EGCG at the highest concentration tested (10 μM) tended to produce a lipolytic stimulating effect in adipocytes derived from the HFD-fed rats, although its lipolytic stimulating action did not reach a statistical significance. The reason for the increase in lipolytic response to EGCG in the presence of isoprenaline in the HFD-fed rat derived adipocytes is still unknown. It is not known if there is an alteration in isoprenaline-induced lipolytic pathway in HFD-derived adipocytes. In adipocytes derived from the HFD-fed rats, isoprenaline may probably act mainly via ERK-independent lipolytic pathway. The ERK-dependent lipolytic pathway is thus reserved for EGCG activation. Thus, the lipolytic stimulating action of EGCG can be observed more clearly. It has been documented that HFD feeding affects on various key components involving in adipocyte lipolysis such as perilipin, HSL enzyme, ATGL enzyme (Collin et al., 2006; Gaidhu et al., 2010). The modifications of these proteins may partly be the reason for the dissimilar responses to EGCG in adipocytes derived from the different diet type-fed rats.
In conclusion, EGCG at every concentration tested (1, 5 and 10 μM) significantly stimulated basal adipocyte lipolysis in the NPD-fed rat derived adipocytes but a significant lipolytic stimulating action of EGCG was found only when 5 μM EGCG was added in adipocytes derived from the HFD-fed rats. Thus, adipocytes derived from the HFD-fed rats were less responsive to the lipolytic action of EGCG in the condition of basal lipolysis. Present findings partially explain and confirm the mechanism of weight-controlling action of EGCG. In addition, the results found here also imply that EGCG possesses the protective role against obesity since its lipolytic action was prominent only when over accumulation of the adipose tissue has not been found yet. EGCG was likely to produce less lipolytic stimulating action in the condition of isoprenaline-induced lipolysis in adipocytes derived from both the NPD-fed rats and the HFD-fed rats. Although the reason for the difference in EGCG response found in this study can be reasonably proposed as mentioned earlier, further study is essentially required to prove those speculations, especially in the molecular aspects.
ACKNOWLEDGMENTS
This study was financially supported by Mahasarakham University research grant year 2012. We thank the Faculty of Pharmacy, Mahasarakham University for the laboratory support. We would like to express our gratitude to Associate Professor Songsak Jumpawadee, Faculty of Veterinary Medicine and Animal Sciences, Mahasarakham University for preparation of animal diet. We are also grateful to Dr. Benjamart Cushnie and Dr. Sakulrat Rattanakiat for their useful comments on the project.
REFERENCES
- Arner, P., 2005. Human fat cell lipolysis: Biochemistry, regulation and clinical role. Best Pract. Res. Clin. Endocrinol. Metab., 19: 471-482.
Direct Link - Balentine, D.A., S.A. Wiseman and L.C. Bouwens, 1997. The chemistry of tea flavonoids. Crit. Rev. Food Sci. Nutr., 37: 693-704.
CrossRefPubMedDirect Link - Biro, F.M. and M. Wien, 2010. Childhood obesity and adult morbidities. Am. J. Clin. Nutr., 91: 1499S-1505S.
CrossRef - Bost, F., M. Aouadi, L. Caron, P. Even and N. Belmonte et al., 2005. The extracellular signal-regulated kinase isoform ERK1 is specifically required for in vitro and in vivo adipogenesis. Diabetes, 54: 402-411.
PubMed - Cash, S.W., S.A. Beresford, J.A. Henderson, A. McTiernan, L. Xiao, C.Y. Wang and D.L. Patrick, 2011. Dietary and physical activity behaviours related to obesity-specific quality of life and work productivity: Baseline results from a worksite trial. Br. J. Nutr., 6: 1-9.
PubMed - Chan, C.Y., L. Wei, F. Castro-Munozledo and W.L. Koo, 2011. (-)-Epigallocatechin-3-gallate blocks 3T3-L1 adipose conversion by inhibition of cell proliferation and suppression of adipose phenotype expression. Life. Sci., 89: 779-785.
Direct Link - Collin, P., N. Chapados, E. Dufresne, P. Corriveau, P. Imbeault and J.M. Lavoie, 2006. Time course of changes in in vitro lipolysis of intra-abdominal fat depots in relation to high-fat diet-induced hepatic steatosis in rats. Br. J. Nutr., 96: 268-275.
PubMed - Gaidhu, M.P., N.M. Anthony, P. Patel, T.J. Hawke and R.B. Ceddia, 2010. Dysregulation of lipolysis and lipid metabolism in visceral and subcutaneous adipocytes by high-fat diet: Role of ATGL, HSL and AMPK. Am. J. Physiol. Cell Physiol., 298: C961-C971.
CrossRef - Gehart, H., S. Kumpf, A. Ittner and R. Ricci, 2010. MAPK signaling in cellular metabolism: Stress or wellness?. EMBO Rep., 11: 834-840.
CrossRef - Greenberg, A.S., W.J. Shen, K. Muliro, S. Patel, S.C. Souza, R.A. Roth and F.B. Kraemer, 2001. Stimulation of lipolysis and hormone-sensitive lipase via the extracellular signal-regulated kinase pathway. J. Biol. Chem., 276: 45456-45461.
Direct Link - Harris, R.B., 1991. Growth measurements in Sprague-Dawley rats fed diets of very low fat concentration. J. Nutr., 121: 1075-1080.
PubMed - Ito, A., T. Suganami, Y. Miyamoto, Y. Yoshimasa, M. Takeya, Y. Kamei and Y. Ogawa, 2007. Role of MAPK phosphatase-1 in the induction of monocyte chemoattractant protein-1 during the course of adipocyte hypertrophy. J. Biol. Chem., 282: 25445-25452.
PubMed - Lee, M.S., C.T. Kim and Y. Kim, 2009. Green tea (-)-epigallocatechin-3-gallate reduces body weight with regulation of multiple genes expression in adipose tissue of diet-induced obese mice. Ann. Nutr. Metab., 54: 151-157.
PubMed - Lee, M.S., C.T. Kim, I.H. Kim and Y. Kim, 2009. Inhibitory effect of green tea catechin on the lipid accumulation in 3T3-L1 adipocytes. Phytother. Res., 23: 1088-1091.
PubMed - Luppino, F.S., L.M. de Wit, P.F. Bouvy, T. Stijnen, P. Cuijpers, B.W. Penninx and F.G. Zitman, 2010. Overweight, obesity and depression: A systematic review and meta-analysis of longitudinal studies. Arch. Gen. Psychiatry., 67: 220-229.
CrossRefPubMedDirect Link - Naim, M., J.G. Brand, M.R. Kare and R.G. Carpenter, 1985. Energy intake, weight gain and fat deposition in rats fed flavored, nutritionally controlled diets in a multichoice (cafeteria) design. J. Nutr., 115: 1447-1458.
CrossRefPubMedDirect Link - Pulbutr, P., K. Thunchomnang, K. Lawa, A. Mangkhalathon and P. Saenubol, 2011. Lipolytic effects of zingerone in adipocytes isolated from normal diet-fed rats and high fat diet-fed rats. Int. J. Pharmacol., 7: 629-634.
CrossRefDirect Link - Rodbell, M., 1964. Metabolism of isolated fat cells. I. Effects of hormones on glucose metabolism and lipolysis. J. Biol. Chem., 239: 375-380.
PubMed - Srinivasan, K., B. Viswanad, L. Asrat, C.L. Kaul and P. Ramarao, 2005. Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: a model for type 2 diabetes and pharmacological screening. Pharmacol. Res., 52: 313-320.
PubMed - Sung, H.Y., C.G. Hong, Y.S. Suh, H.C. Cho and J.H. Park et al., 2010. Role of (-)-epigallocatechin-3-gallate in cell viability, lipogenesis, and retinol-binding protein 4 expression in adipocytes. Naunyn. Schmiedebergs. Arch. Pharmacol., 382: 303-310.
PubMed - Thielecke, F. and M. Boschmann, 2009. The potential role of green tea catechins in the prevention of the metabolic syndrome: A review. Phytochemistry, 70: 11-24.
CrossRefPubMedDirect Link - Von Lengerke, T. and C. Krauth, 2011. Economic costs of adult obesity: A review of recent European studies with a focus on subgroup-specific costs. Maturitas, 69: 220-229.
PubMed