
Dewoowoogen Porras Baclayon
Department of Applied Biological Sciences, Faculty of Agriculture, Kagawa University,
Miki, Kagawa 761-0795, Japan
Toshiyuki Matsui
Department of Applied Biological Sciences, Faculty of Agriculture, Kagawa University,
Miki, Kagawa 761-0795, Japan
Haruo Suzuki
Department of Applied Biological Sciences, Faculty of Agriculture, Kagawa University,
Miki, Kagawa 761-0795, Japan
Yusuke Kosugi
Department of Applied Biological Sciences, Faculty of Agriculture, Kagawa University,
Miki, Kagawa 761-0795, Japan
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 4 | Page No.: 614-619







ABSTRACT
Tissue toughness or textural changes which are closely associated with lignifications in vegetables often provide problem to consumer acceptability particularly those which are consumed raw or half-cooked. To obtain fundamental knowledge underlying this change, enzymatic and molecular analyses of phenylalanine ammonia-lyase (PAL; EC 4.3.1.5), enzyme catalyzing the conversion of L-phenylalanine to trans-cinnamic acid in the first step of phenylpropanoid pathway were conducted in the edible branchlets of broccoli during early postharvest senescence. While toughness of the tissue or breaking force as estimated by resistance to pressure on shearing continuously increased, PAL activity decreased after few hours from harvest. The expression of BoPAL was at maximum level on the 12 h from harvest in the upper branchlets. In the lower branchlets, the expression was initially high but declined 6 h from harvest and slightly increased until 24 h of storage. The results imply that, in addition to PAL, other lignin-specific enzymes and/or tissue structural changes may have more significant roles in tissue toughening of broccoli branchlets.
PDF Abstract XML References Citation
How to cite this article
Dewoowoogen Porras Baclayon, Toshiyuki Matsui, Haruo Suzuki and Yusuke Kosugi, 2007. Changes in Activity and Gene Expression of Phenylalanine Ammonia-lyase in Broccoli Branchlets During Early Postharvest Senescence. Journal of Biological Sciences, 7: 614-619.
DOI: 10.3923/jbs.2007.614.619
URL: https://scialert.net/abstract/?doi=jbs.2007.614.619
DOI: 10.3923/jbs.2007.614.619
URL: https://scialert.net/abstract/?doi=jbs.2007.614.619
INTRODUCTION
Perishable plant commodities are inevitably exposed to ambient temperatures during early postharvest handling. Immediately after harvest, plant cells are very active such that they are vulnerable to adverse environmental conditions. It has been reported that major physiological, biochemical and molecular changes have occurred in broccoli heads within hours after severed from the mother plant. Major losses of sugars, organic acid and proteins were detected during the first 6 h of storage (King and Morris, 1994a, b; Pramanik et al., 2005). Moreover, textural changes or toughening of branchlets or brocco in Italian which means branch during storage often provide problem to consumer acceptability. Texture is an important product quality especially in vegetables consumed raw or half-cooked.
Textural changes or toughening of plant tissues is closely associated with lignin formation. Lignin, a polymer of aromatic subunits derived from phenylalanine serves as a matrix around the polysaccharide components of some plant cell walls, providing rigidity and comprehensive strength (Whetten and Sederoff, 1995). Phenylalanine ammonia-lyase (EC 4.3.1.5; PAL), catalyzes the committed step in phenylpropanoid metabolism, hence it could have a regulatory role in controlling biosynthesis of all phenylpropanoid compounds including lignin (Northcote, 1985). Analysis of phenylpropanoid metabolites in transgenic tobacco showed that lignin content was affected when PAL activity is reduced to 20-25% of wild type levels (Bate et al., 1994). The enzyme has also been reported to control the synthesis of lignin causing textural changes in bamboo shoots (Chen et al., 1989; Matsui et al., 2004). Moreover, PAL activity was found to be enhanced in several plant species in response to wounding imposed by either harvest or other stresses during postharvest handling as observed in newly harvested asparagus spears (Bhowmik et al., 2003), lettuce (Lopez-Galvez et al., 1996) and Cucurbita maxima (Kato et al., 2000). Hence, it is worth noting that textural changes in broccoli could be induced by stress in response to harvest and trimmings.
Understanding the early changes in response to stress in harvested perishable commodity would provide fundamental knowledge for better interpretation of subsequent changes and designing appropriate control measures. To our knowledge, the possible involvement of PAL in the textural changes of the edible branchlet of broccoli has not been thoroughly investigated. Hence, this study was conducted to examine the changes in PAL activity and gene expression patterns in relation to texture in the edible branchlet tissues during early postharvest handling.
MATERIALS AND METHODS
Plant material: Field-grown broccoli (Brassica oleracea var. Endeavor) heads were harvested in March 2006 from Kagawa Agricultural Experiment Station, Miki, Kagawa, Japan. Right after harvest, the heads were trimmed, enclosed with perforated plastic sheet and kept at 20 °C for 30 h. At 6 h interval, the branchlets were separated from the head. The edible branchlets were classified into lower tissues which included the first four lower branchlets and the upper tissues which included the rest of the upper branchlets. The texture as estimated by the resistance of the tissues to pressure on shearing was determined at harvest (0 h) and after 6, 12, 18, 24 and 30 h in both upper and lower branchlets. The sample tissues were immediately kept at -30 and -80 °C until enzyme analysis and RNA extraction, respectively.
Texture measurement and PAL extraction and assay: The texture or breaking force of the branchlet tissues was determined following the method of Matsui et al. (2005) while the extraction and assay procedures for the determination of PAL activity were performed as described by Matsui et al. (2004). The result of PAL activity was expressed as nmol of trans-cinnamate formed per hour per milligram of protein.
RNA extraction and amplification of poly (A) + RNA by RT-PCR: RNA was extracted from the branchlet tissue of the broccoli head following the hot borate method of Wan and Wilkins (1994). The first strand cDNA was synthesized as described in our previous report (Baclayon et al., 2006). The primers 5 ’-ATY GAG GCT GCT GCY ATT ATG-3 ’ (upstream) and 5 ’-ACA TCT TGG TTG TGY TGC TC-3 ’ (downstream) were designed (Matsui et al., 2004) and synthesized on the basis of the amino acid domains (IEAAAIM and AEQHNQD, respectively) conserved from various PAL genes in the database. The PCR procedure was performed in an ASTEC Program Temperature Control System PC-700 under the following conditions of initial denaturation at 95 °C for 10 min followed by 35 cycles of 40 sec at 95 °C, 40 sec at 63 °C, 40 sec at 72 °C and 5 min at 72 °C. The PCR products were confirmed by gel electrophoresis using 1% agarose gel stained with 1 μg mL -1 ethidium bromide and visualized under UV light.
Cloning and sequencing of cDNA: The amplified cDNA product was ligated to the plasmid pT7Blue vector (Novagen, Inc., USA) using DNA ligation Kit v2.1 (TaKaRa Bio, Inc., Japan) and cloned into E. coli strain DH5α (Invitrogen Life Technologies, USA) and plated on LB/ampicilin/IPTG/x-gal medium. White clones/colonies were chosen for plasmid extraction. Bacterial plasmid DNA was extracted using QIAprep ® Spin Miniprep Kit (QIAGEN Sciences, USA). Sequencing of cDNA was carried out on ABI 3100 Genetic Analyzer (Applied Biosystems, USA) using a BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, USA).
Sequence data analysis: The sequence of the cloned cDNA fragment was compared with those in the GenBank. Homology analysis and phylogenetic tree construction were performed using computer software BioEdit (Hall, 1999) and Gapped BLAST and PSI-BLAST (Altschul et al., 1997).
Probe synthesis and RNA blot analysis: Probe synthesis and northern blot analysis were performed as described in the previous report (Baclayon et al., 2006).
Gene bank accession number: The nucleotide sequence of partial cDNA was submitted to the nucleotide sequence database of the Center for Information Biology and DNA Data Bank of Japan (DDBJ) and was designated as BoPAL with assigned accession number of AB281591.
RESULTS
Changes in breaking force: Initially, the breaking force, which indicated the toughness of the tissues, of the lower branchlets was greater than that of the upper branchlets (Fig. 1A). However, after 12 h from harvest, the breaking force of the upper branchlets started to increase and slightly exceeded that of the lower branchlets. At the end of the experimental period, the upper branchlets had a significantly higher resistance to pressure than the lower ones.
PAL activity: A transient increase in PAL activity was observed in the upper branchlets 12 h after harvest (Fig. 1B). Thereafter, the enzyme activity rapidly declined until the end of the experimental period. In the lower branchlets, the enzyme activity slightly declined after harvest and remained almost unchanged until 24 h of storage, then again declined at hour 30. There was a significant higher PAL activity in the upper than the lower branchlets throughout the storage period.
Isolation and identification of the cDNA clone encoding PAL and phylogenetic tree analysis: The partial clone cDNA (BoPAL; AB281591) encoding a harvest-induced transcript from broccoli branchlets contained 486 nucleotides (Fig. 2). Based on BLAST searching in the DDBJ/EMBL/GenBank database, BoPAL exhibited high sequence homology with PAL gene from other plant species (Table 1). The nucleotide sequence is 87% homologous to PAL from Brassica napus (AY795080), Raphanus sativus (AB087212) and Latuca sativa (AF 411134) and 98, 96, 95 and 89% homologous to PAL from Arabidopsis thalina (AK221928), R. sativus, B. napus and L. sativa, respectively, at the deduced amino acid level.
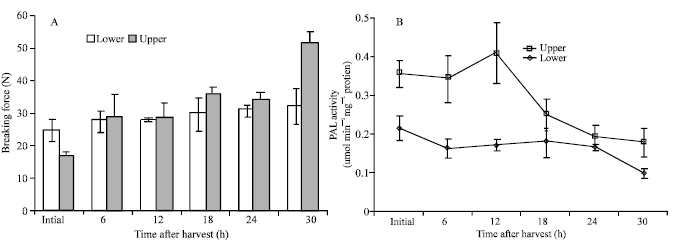 | |
| Fig. 1: | Changes in breaking force or resistance to pressure on shearing (A) and activities of phenylalanine ammonia-lyase (B) in broccoli branchlets during early postharvest senescence. Each point represents the mean of three replications and bars show SE |
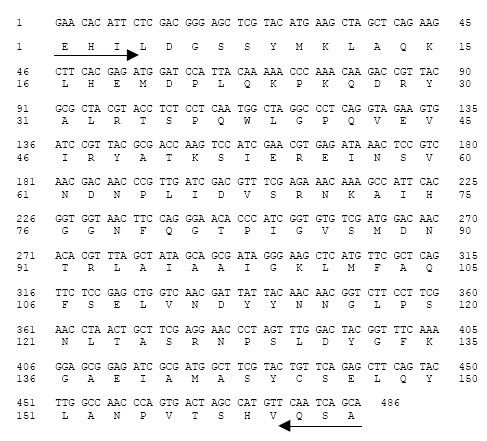 | |
| Fig. 2: | Nucleotide and deduced amino acid sequences of the cDNA clone corresponding BoPAL. The predicted amino acid sequence is given in single letter code for each amino acid. The arrows indicate the position of degenerate primers (sense |
The phylogenetic tree derived from deduced amino acid sequences analyzed by Neighbor-Joining/UPGMA method version 3.6a2.1 using BioEdit software revealed a closer relationship between B. olaracea with A. thalina, R. sativus and B. napus; all of Brassicaceae family (Fig. 3).
PAL gene expression: The expression of BoPAL encoding PAL transcript from broccoli branchlets was examined by RNA blot analysis of total RNA with a Dig-labeled probe.
| Table 1: | Percent homology of nucleotide and deduced amino acid sequences between PAL from broccoli and other plant species in the database |
 | |
Brassica oleracea (AB281591) was calculated as 100% | |
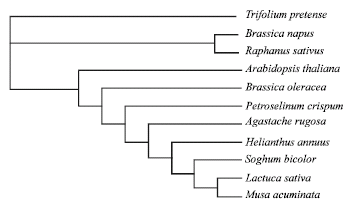 | |
| Fig. 3: | Phylogenetic tree of the BoPAL deduced amino acid sequence with other plant species in the database. The amino acid sequences were aligned using Clustral multiple alignment and phylogenetic tree was constructed by Neighbor-Joining/UPGMA method ver. 3.6a2.1 using BioEdit software |
The maximum level of transcript was observed on the 12 h from harvest in the upper branchlets. In the lower branchlets, the expression was initially high but declined 6 h from harvest and slightly increased until 24 h of storage (Fig. 4).
DISCUSSION
Texture is an important quality of vegetable crops especially those which are eaten raw or half-cooked. The edible portion of broccoli head is termed as branchlets and florets; the former usually toughens after harvest. Toughening or textural changes in plant tissues is closely associated with lignin formation. In this study, the texture as estimated by resistance to pressure on shearing continuously increased after harvest (Fig. 1A). The upper branchlets which are composed of less mature tissues became tougher than the lower ones at the end of the experimental period. However, the activity of PAL, one of the enzymes involved in the lignification process declined 12 h and immediately after harvest in the upper and lower tissues, respectively (Fig. 1B). The same pattern was
found in asparagus spears that fiber content generally increased throughout the experimental period while PAL activity decreased after 24 h from harvest (Bhowmik et al., 2003). This result may suggest that PAL activity could not be pointed out as a single factor controlling the toughening of the branchlet tissues; other lignin-specific enzyme is likely involved in the regulation related to lignin biosynthesis or textural changes. For instance, cinnamyl alcohol dehydrogenase (CAD) has been singled out as the primary gene involved in lignin biosynthesis in the floral stem of A. thaliana by supplying both coniferyl and sinapyl alcohols (Sibout et al., 2005). Moreover, specific promoter elements common to PAL and CAD genes (Whetten and Sederoff, 1995) as found in Arabidopsis PAL1 (Ohl et al., 1990), bean PAL2 (Leyva et al., 1992) and poplar plants CAD (Feuillet et al., 1995) could have regulatory role in the liginification process. However, these suggestions need further elucidation as other factors are also likely to be involved in the toughening of brassica tissues. Femenia et al. (1998), for instance, suggested that localization of pectic polysaccharide in cell wall of cauliflower stem tissues could be related to textural properties of cell wall. Furthermore, the thickened vascular cell walls in broccoli stems are perceived tough enough to cause problems with consumer acceptability (Muller et al., 2003).
To obtain fundamental knowledge on the induction of PAL in broccoli branchlets during early postharvest senescence, a cDNA fragment was isolated using reverse transcription-polymerase chain reaction. The partial clone cDNA encodes an mRNA of 486 bp (Fig. 2). The nucleotide sequence had 87% homology with B. napus, R. sativus and 84% with A. thaliana (Table 1); clustered closer in a subgroup belonged to Brassicaceae family (Fig. 3). RNA blot analysis showed that BoPAL transcript was highly expressed during the first 12 h from harvest but declined with some fluctuations thereafter (Fig. 4).
 | |
| Fig. 4: | RNA gel blots analysis of BoPAL transcript. The upper panels (A) are gel blots of total RNA (each lane containing 10 μg) isolated from broccoli branchlets and separated by electrophoresis on 1% agarose gel containing 1x MOPS and 5% formaldehyde. The bottom panels (B) are representatives from ethidium bromide stained gels showing rRNA |
The expression pattern which coincided with the enzyme activity further revealed that toughening of the tissues in broccoli branchlets could be partly due to PAL. Therefore, based on the results of enzymatic and molecular analyses, efforts in preventing lignifications due to PAL could unlikely maintain the textural quality of the tissues during storage. Further studies on the activities and gene expressions of other lignin-associated enzymes and the mechanical and structural properties of the cell walls in the tissues of the edible branchlets are helpful in attempting to maintain textural quality in broccoli during early postharvest handling.
ACKNOWLEDGMENT
The authors gratefully acknowledge the financial support of the Government of Japan through the Ministry of Education, Culture, Science and Technology.
REFERENCES
- Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Porras Baclayon, D., M. Toshiyuki, S. Haruo and K. Yusuke, 2006. Cloning, sequencing and expression analysis of a cDNA encoding glutamate dehydrogenase gene in broccoli during postharvest senescence. Biotechnology, 5: 118-124.
CrossRefDirect Link - Bate, N., J. Orr, W. Ni, A. Meromi and T. Nadler-Hassar et al., 1994. Quantitative relationship between phenylalanine ammonia-lyase levels and phenylpropanoid accumulation in transgenic tobacco identifies a rate-determining step in natural product synthesis. Proc. Nat. Acad. Sci. USA., 91: 7608-7612.
Direct Link - Bhowmik, P.K., T. Matsui, H. Suzuki and Y. Kosugi, 2003. A phenylalanine ammonia-lyase gene from asparagus: cDNA cloning, sequence and expression in response to wounding. Asian J. Plant Sci., 2: 425-430.
CrossRefDirect Link - Femenia, A., P. Garosi, K. Roberts, K.W. Waldron, R.R. Selvendran and J.A. Robertson, 1998. Tissue-related changes in methyl-esterification of pectic polysaccharides in cauliflower (Brassica oleracea L. var. botrytis) stems. Planta, 205: 438-444.
Direct Link - Feuillet, C., V. Lauvergeat, C. Deswarte, G. Pilate, A. Boudet and J. Grima-Pettenati, 1995. Tissue- and cell-specific expression of a cinnamyl alcohol dehydrogenase promoter in transgenic poplar plants. Plant Mol. Biol., 27: 651-667.
Direct Link - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Kato, M., Y. Hayakawa, H. Hyodo, Y. Ikoma and M. Yano, 2000. Wound induced ethylene synthesis and formation of 1-aminocyclopropane-1-carboxylate (ACC) synthase. ACC oxidase, phenylalanine ammonia-lyase and peroxidase in wounded mesocarp tissue of Cucurbita maxima. Plant Cell Physiol., 41: 440-447.
Direct Link - King, G.A. and S.C. Morris, 1994. Early compositional changes during postharvest senescence of broccoli. J. Am. Soc. Hortic. Sci., 119: 1000-1005.
Direct Link - Leyva, A., X. Liang, J.A. Pintor-Toro, R.A. Dixon and C.J. Lamb, 1992. cis-element combinations determine phenylalanine ammonia-lyase gene tissue-specific expression patterns. Plant Cell, 4: 263-271.
Direct Link - Lopez-Galvez, G., M. Saltviet and M. Cantwell, 1996. Wound-induced phenylalanine ammonia-lyase activity: Factors affecting its induction and correlation with the quality of minimally processed lettuces. Postharv. Biol. Technol., 9: 223-233.
Direct Link - Matsui, T., P.K. Bhowmik and K. Yokozeki, 2004. Phenylalanine ammonia-lyase in moso bamboo shoot: Molecular cloning and gene expression during storage. Asian J. Plant Sci., 3: 315-319.
CrossRefDirect Link - Matsui, T., Y. Kosugi and E. Miki, 2005. Postharvest changes in activities of GS, ACC synthase, ACC oxidase and PAL in bamboo shoots at different storage temperatures. Food Preserv. Sci., 31: 303-308.
CrossRefDirect Link - Muller, S., W.G. Jardine, B.W. Evans, R.J. Vietor, C.E. Snape and M.C. Jarvis, 2003. Cell wall composition of vascular and parenchyma tissues in broccoli stems. J. Sci. Food Agric., 83: 1289-1292.
Direct Link - Ohl, S., S.A. Hedrick, J. Chory and C.J. Lamb, 1990. Functional properties of a phenylalanine ammonia-lyase promoter from Arabidopsis. Plant Cell, 2: 837-848.
Direct Link - Pramanik, B.K., T. Matsui, H. Suzuki and Y. Kosugi, 2005. A sucrose synthetase gene from broccoli: cDNA cloning, sequencing and its expression during storage. Biotechnology, 4: 288-295.
Direct Link - Sibout, R., A. Eudes, G. Mouille, B. Pollet, C. Lapierre, L. Jouanin and A. Seguin, 2005. Cinnamyl alcohol dehydrogenase-C and-D are the primary genes involved in lignin biosynthesis in the floral stem of Arabidopsis. Plant Cell, 17: 2059-2076.
Direct Link - Wan, C.Y. and T.A. Wilkins, 1994. A modified hot borate method significantly enhances the yield of high quality RNA from cotton (Gossypium hirsutum L.). Anal. Biochem., 223: 7-12.
Direct Link