
Luqman Hakim
Faculty of Animal Science, Brawijaya University (UB), Malang, Indonesia
LiveDNA: 62.32118
Agus Susanto
Faculty of Animal Science, University of Jenderal Soedirman (UNSOED), Purwokerto, Indonesia
LiveDNA: 62.32119
Agus Budiarto
Faculty of Animal Science, Brawijaya University (UB), Malang, Indonesia
International Journal of Dairy Science
Year: 2020 | Volume: 15 | Issue: 2 | Page No.: 99-107







ABSTRACT
Background and Objective: Recording linear traits in a dairy breeding program by scoring the traits have become a more common practice in developed countries where certified classifiers are available. Estimating the genetic parameters of linear traits (measured directly on the animals) of Holstein cows population raised by BBPTUHPT Baturraden of Indonesia was the aim of this paper. Materials and Methods: A total 323 lactating primiparous Holstein cows were measured for 6 linear traits (angularity, stature, body depth, rump angle, rump width and chest width) and 6,961 animals in the pedigree were included. The systematic environmental factors included month and year of birth. The variance component analysis was performed by using Restricted Maximum Likelihood Method (REML) incorporated in DMU program. Results: The means of stature, angularity, chest width, body depth, rump angle and rump width were all within intermediate category. The heritability estimates of the corresponding linear traits ranged from small to medium where angularity and rump width had the lowest and highest heritability, respectively. The phenotypic correlations were very low and varied where angularity-rump width and angularity-rump angle are having the smallest and biggest coefficient of correlation, respectively. The genetic correlation were also very low and varied being chest width-angularity and rump angle-angularity to have the smallest and highest coefficient of correlation. Conclusion: The new approach of recording the linear traits by measuring directly the traits is possible with reasonable variability. The considerable magnitude of heritabilities indicates that the traits will respond to a selection program incorporating those linear traits. The nearly zero estimates of the correlations could be improved by using a more complete pedigree file in the analysis.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Luqman Hakim, Agus Susanto and Agus Budiarto, 2020. Heritability and Correlation of Linear Traits in Holstein Cows in Indonesia. International Journal of Dairy Science, 15: 99-107.
DOI: 10.3923/ijds.2020.99.107
URL: https://scialert.net/abstract/?doi=ijds.2020.99.107
DOI: 10.3923/ijds.2020.99.107
URL: https://scialert.net/abstract/?doi=ijds.2020.99.107
INTRODUCTION
The milk yield of dairy cattle has successfully been increased through both genetic improvement programs and non-genetic ones such as; improved feed/nutrient, health and reproduction management. This however has caused some drawback in other aspects of production that have drawn serious concerns from animal geneticists. Numerous studies have shown that despite the success of increased milk yield, reproductive performances of dairy cows have deteriorated1-4. The same phenomena were also reported on health performance of dairy cows5,6. Even though the evidence of increased milk yield and reduced reproductive and health performances were clear, the underlying biological mechanism of these relationships remains uncover. The antagonistic relation between milk yield and reproduction (or health) could be explained by a theory so called Resource Allocation Theory7 saying that if an output e.g., milk yield is increased, the other functions such as; fertility and health are affected due to limited resources that animals have.
Selection strategies to accommodate the antagonistic relationship between milk yield and reproduction (or health) have been introduced by incorporating linear traits in addition to the milk yield. Linear traits have been reported to be genetically associated with both production (e.g., milk yield)8,9 and functional traits (reproduction and health)10,11. They also generally have bigger heritability than reproduction or health related-traits12,13 and hence selection is more effective on linear traits than directly on reproductive and health traits. Milk yield and its component were reported in the literatures to have medium heritability estimate14,15.
The National Breeding Centre for Dairy Cows and Forage of Indonesia (BBPTU-HPT Baturraden) has focused on milk yield and has not incorporated linear traits in its genetic improvement program. Inclusion of linear traits in a breeding program requires the knowledge of genetic parameters of those traits. Instead of being scored, measuring linear traits recommended by WHFF16 directly on the cows was initiated in the centre. Lacking the certified classifiers for scoring the linear traits was one of reasons why the traits were measured directly instead of scored. The information about the genetic correlation between traits is also useful for management to decide what traits are included in the recording system. Hence, the objective of the present study was to estimate the genetic parameters of the selected linear traits i.e. angularity, stature, body depth, rump angle, rump width and chest width of Holstein dairy cows in The National Breeding Centre for Dairy Cows and Forage of Indonesia (BBPTU-HPT Baturraden).
MATERIALS AND METHODS
Experimental site: Phenotypic Data of Angularity (ANG), Stature (STA), Body Depth (BD), Rump Angle (RA), Rump Width (RW) and Chest Width (CW) and pedigree file were obtained from the National Breeding Centre for Dairy and Forage of Indonesia (BBPTU-HPT Baturraden). The breeding centre is located in Baturraden, Central Java of Indonesia which have the average ambient temperature of 18-28°C, 70-80% humidity and 6000-9000 mm rainfall per year17. The center is situated in an upland of +600 m above sea level. This study project was conducted from July 1, 2017 to December 15, 2018 in BBPTU-HPT Baturraden, Indonesia.
Materials and research tools: The linear traits were measured directly on lactating Holstein cows. Data pedigree were retrieved from the database available in spreadsheet type files owned by the centre. Since animal identification were using alphanumeric, RE-NUM-OR program of Yazgan18 was used to convert and re-order the pedigree file so that it is usable for the variance component analysis by DMU program19. Angularity and rump angle were measured with digital angle level of Nankai® using angle degree unit (°). Body depth, rump width, chest width and stature were measured with caliper using centimeter (cm) unit measurement. More detailed information about these selected linear traits measurements was reported by Susanto et al.20.
Research procedure: Primary data (linear traits) were measured and collected directly on lactating cows of Holstein cows. Secondary data i.e., date of birth (age) were retrieved from the database owned the breeding centre. Individual’s animal identification were available for each animal available in the farm. Pedigree data were obtained from the database and animal’s identification were converted into integer format for further analysis. Farm location where animals were reared was identified and included in the model analysis. Data was then analyzed using Restricted Maximum Likelihood (ReML) for the variance components.
Data collection: The traits were measured within the months of July-December, 2017. As many as 323 lactating Holstein cows were measured for the selected linear traits along with its systematic environmental factors. The cows were reared in an intensive system where grass and concentrate were provided according to the production stage. The animals are most of the time housed in a free stall barn, though the animals are sometimes released into a grazing field to exercise where elephant grass are cultivated and taken care. The cows are milked twice daily in a modern milking parlour and milk yield is recorded individually.
Experimental design: The study was conducted through a survey through a purposive sampling method where all lactating dairy cows available in the farm were included in the study. The six linear traits i.e., Angularity (ANG), Stature (STA), Body Depth (BD), Rump Angle (RA), Rump Width (RW) and Chest Width (CW) were measured directly on the lactating cows. Other systematic environmental effects recorded in the study were month of birth and year of birth and age of cows (month) when they were measured.
Parameters measured: Data of angularity, stature, body depth, rump angle, rump width and chest width of Holstein lactating cows were measured directly from the animals instead of scored. The traits were measured following the definition given by WHFF16. Angularity and rump angle were measured in a unit of angle degree and stature, body depth, rump width and chest width were measured in a unit of cm length. Along with the linear traits, the month of birth, year of birth and the age of animals when the linear traits were measured were also recorded. Angularity was defined as the angle of the ribs measured from the horizontal line by disregarding the openness and the flatness of the bones. Body depth was measured as the distance between the top of spine and bottom of barrel at last rib. Rump angle was measured as the angle of the rump structure from hips to pins from the vertical line. Rump width was measured as the distance between the most posterior points of pin bones. Chest width was measured as the distance from the inside surface between the top of the front legs. Stature was measured as the distance from top of the spine between hips to ground. Definition of the measured linear traits was according to WHFF16 except for angularity which did not include ribs openness and flatness.
Statistical analysis: Animal mixed model was used in variance component analysis including month and year of birth as fixed
effects, individual animal as random effect and age of animals when the linear traits were measured as random-fixed covariate (covariable) effects. Multivariate of 6 linear traits with equal design model was employed in variance component analysis. The mathematical model used was as the following:
Yij = μ+F1+b (Xij)+ai+eij | (1) |
where, yij is the corresponding trait being analyzed (angularity, stature, body depth, rump angle, rump width and chest width), μ is the overall mean, Fi is the fixed effect of month and year of birth, b is the regression coefficient of yij on age when the linear traits were measured, a is the random additive genetic effect of animals and e includes the random residual effects. The variance structure of genetic additive and residual random effects were assumed as var(a) = Aσ2a and var(e) = Iσ2e where, A and I are numerator additive relationship matrix between animals in the pedigree file and identity matrix, respectively. No covariance between random effects are assumed in the model (cov(a,e) = cov (a,e) = 0). DMU1 and DMUAI modules of DMU software19 which is based on restricted maximum likelihood approach were used to estimate variance/covariance of the random effects. The maximum likelihood computation was according to Expectation Maximization (EM) algorithm (option 3 of DMUAI). Prior (starting values) of variance/covariance of the random effects used default values (identity matrix) set by the program. The numerator additive relationship matrix (A matrix) was computed by DMU program with option 1 for inbred population. The computation of standard errors of heritability of each trait and genetic correlation between traits was based on Taylor series approximation19. The convergence criteria for norm of the update vector and of the gradient vector (AI) were set by default within the DMU program.
RESULTS AND DISCUSSION
The basic statistics of selected linear of the present study is presented in Table 1. The means of stature, chest width and rump width are shown along with the means of the traits in 1-9 scale of measurement.
| Table 1: | Basic statistics of the linear traits data |
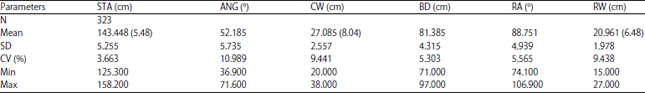 | |
Values in the brackets was computed by converting the observed values into the 1-9 scale scoring system according to WHFF16, means of angularity, body depth and rump angle were not converted into 1-9 scale since these linear traits are not considered as true linear traits16, ANG: Angularity, STA: Stature, BD: Body depth, RA: Rump angle, RW: Rump width, CW: Chest width | |
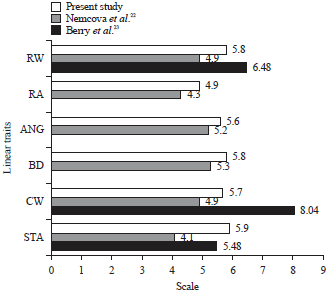 | |
| Fig. 1: | Means of linear traits in 1-9 scale compared with other studies |
| Values for RA, ANG and BD are not available since those traits are not true linear traits | |
The phenotypic variability determined by the coefficient of variation revealed that the new approach of measuring directly the linear traits (as opposed to scoring) resulted in reasonable and low variability where the values of coefficient of variation were ranging 3.663-10.989%. The coefficients of variation of this present study in general are smaller than those reported by Toghiani21, who reported coefficient of variation of 10 linear traits ranging 2.55-32.28%. According to WHFF16, stature and rump width fall into category intermediate and chest width falls into category wide. Compared to the study of Toghiani21 who reported stature and rump width of 139.1+3.5 and 19.3+1.6, respectively, the cows under present study had slightly taller stature and about the same rump width as shown in Table 1. The 1-9 scale converted value of stature was similar to the study of Nemcova et al.22 whereas, chest width and rump width were wider. Nemcova et al.22 reported the mean+sd of stature, chest width and rump width of 5.9+1.3, 5.7+1.3 and 5.8+1.36 cm, respectively. However, Berry et al.23 reported shorter stature, narrower chest width and rump width compared those of present study. Even though Berry et al.23 and Nemcova et al.22 both conducted their study on primiparous cows and Holstein breed using the same 1-9 scale of observation, the results were different (Fig. 1). Many factors could be the cause of these differences such as age at classification, classifiers, stage of lactation and herd-year-season. White24 reported that age at classification had the major effect on linear score whereas, classifiers, stage of lactation and herd-year-season had small effect.
| Table 2: | Heritabilities (standard error) of the linear traits |
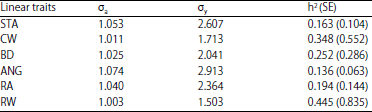 | |
h2: Heritability in the narrow sense | |
Hayes and Mao25 reported that parity, age and stage of lactation affected linear type scores. Angularity, body depth and rump angle which were directly measured on cows instead of scored could not be compared with standard given by the World Holstein Friesian Federation since those traits are not true linear traits16.
Heritabilities and variances: Heritabilities along with the associated standard error estimates obtained in this study are presented in Table 2 where the estimates varied from 0.136 (angularity) to 0.445 (rump width). Except for rump width and chest width, the heritability estimates of the present study were smaller compared to those reported by DeGroot et al.26, Berry et al.23, Dal Zoto et al.11, Nemcova et al.22, Toghiani21 and Mazza et al.27. The heritability estimates obtained from the present study is visualized in Fig. 2 along with those reported in the literature. The heritability estimates of these body-related linear traits which falls into small to medium heritability agree with those reported in the literature. Heritability estimates are not absolute values instead it is the property of a population under study, of the environmental where the animals live and of the method of analysis and measurement of the phenotype28 and therefore, it is expected to have different estimates from study to study. In the present study, the linear traits were measured directly instead of scored in oppose to most studies of linear traits where the traits were scored. Most of the studies on linear traits such as Degroot et al.26, Berry et al.23, Dal Zotto et al.11, Nemcova et al.22 and Mazza et al.27 used 1-9 scale scored by classifiers when recording the traits. In the present study, the linear traits were measured instead of scored because in BBPTU-HPT Baturraden recording linear traits has not been initiated and included in the genetic evaluation. Measuring the linear traits was believed to be more accurate than scoring them when the certified classifiers were not available.
The estimates of variance component estimated using animal model depends on the completeness of the pedigree data used in the analysis.
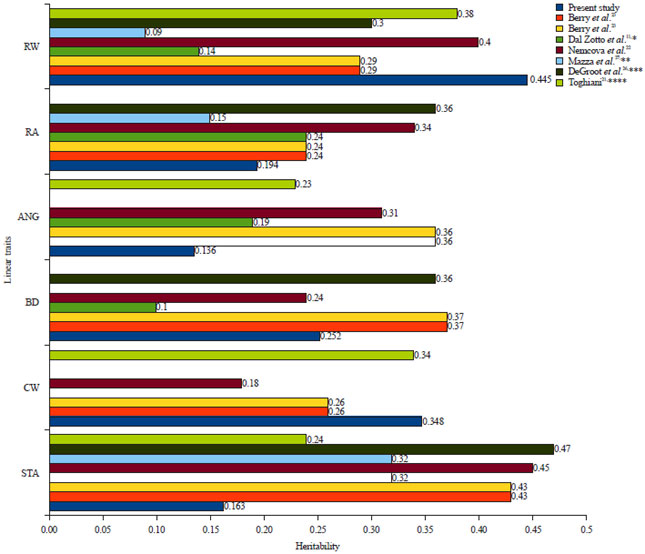 | |
| Fig. 2: | Heritability estimates compared with other studies |
*Heritability of CW was not reported, **Heritability of CW, BD and ANG were not reported, ***Heritability of CW and ANG were not reported, ****Heritability of BD and RA were not reported | |
The contribution of pedigree data to the estimate is through the additive relationship matrix (matrix A) in the Mixed Model Equation (MME). Cantet et al.29 has demonstrated through a simulation study that if the pedigree data used in the analysis is complete then the heritability estimate is close to the actual heritability. Missing pedigree information will result in reduced heritability estimate. The missing information of sire in the pedigree will lead to more bias estimate than missing dam information. Susanto et al.30 has shown that the complexity of pedigree determined by the number generation of random mating and magnitude of the actual heritability influence the strength of association between the estimated and the actual variance components, the more complex of pedigree and the higher heritability of a trait, the stronger is the relationship between the estimated and the actual variance components.
The availability of parents information on the pedigree data recorded in BBPTU-HPT Baturraden are presented in Table 2. As much as 4.38 and 10.19% out of 6,961 animals in the pedigree file had no information of the sire and dam, respectively. As much as 27.71% of the animals in pedigree file was missing both sire and dam information whereas only 57.72% of the animals in pedigree file had complete information of both sire and dam. Obtaining complete pedigree data in developing countries including Indonesia is a challenge.
The heritability estimates of the present study ranging from 0.136 (angularity) to 0.445 (rump width) indicated that the linear traits under study by measuring directly the traits are heritable and any either direct or indirect selection on these traits will result in some changes in the phenotypic means of corresponding traits. Since these traits have been reported to be genetically correlated with other important traits, care has to be taken when developing breeding objective so that the achieved genetic improvement on the main selected trait will not adverse the other traits. Or, these linear traits could be used to genetically improve other traits in an indirect selection program due the existence of genetic correlation between these linear traits with other important traits. Chest width and angularity were reported to be genetically correlated with somatic cell count31,32. Rump width and rump angle were genetically correlated with the incidence of distochia33. Chest width was genetically associated calving intervals and conception rate to first service31. Royal et al.34 have reported that chest width and rump width were genetically associated with commencement of luteal activity. Stature, body depth, rump angle, rump width and angularity were reported to be genetically associated with survival at age 48, 54 and 84 months35. When recording of phenotypic data of a trait is expensive or difficult, genetic improvement of that particular trait could be done through an indirect selection program by means of traits that have significant genetic correlation with the trait of interest.
Genetics and phenotypic correlation: Variances/co V variances of genetic and phenotypic are presented in Table 3 and 4. Data table showed that in general, as expected, the genetic variances/co-variances are smaller in magnitude compared to the phenotypic variances/co-variances. The variation of these linear traits are attributed to genetic and phenotypic factors. The genetic and phenotypic correlation of the selected linear traits in the present study is given in Table 5. It is shown that the phenotypic correlation varied from -0.0260 (ANG-RW) to 0.0214 (ANG-RA) and the genetic correlation ranged from 0.0009 (CW-ANG) to 0.0214 (RA-ANG).
Most of the phenotypic correlation agreed in direction with those reported by Berry et al.23 except for STA-CW, BD-RA, ANG-RA and ANG-RW. In magnitude however, all the phenotypic correlations of the present study were smaller. The same pattern was also observed in the direction of genetic correlation that almost all correlation were the same except for STA-CW, BD-RA, ANG-RA and ANG-RW. Magnitude of the genetic correlation estimated in the present study were so small (nearly zero). This is probably associated with the data and pedigree structure available for the study. Nilforooshan et al.36 has shown that incomplete pedigree will result in genetic additive variance to be underestimated. This underestimated genetic variance will eventually affect the coefficient of correlation since the correlation is derived from variance/covariance between two variables. The incomplete pedigree data is common and a serious problem in developing countries37. Therefore, the genetic/phenotypic correlation obtained in the present study does not necessarily represent the actual genetic correlation between traits under study. Magnitude of the correlation might be increased if the pedigree data used in the analysis is more complete. The phenotypic and genetic correlation obtained in the present study is visualized in Fig. 3 and 4 along with the results reported in the literatures.
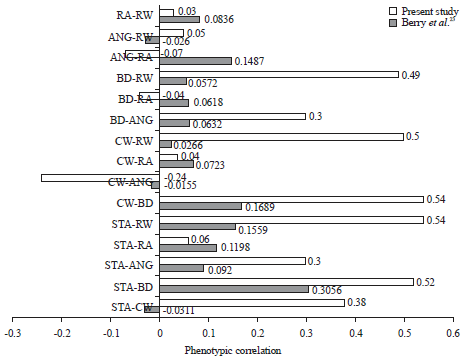 | |
| Fig. 3: | Phenotypic correlation compared with literature23 |
| ANG: Angularity, STA: Stature, BD: Body depth, RA: Rump angle, RW: Rump width, CW: Chest width | |
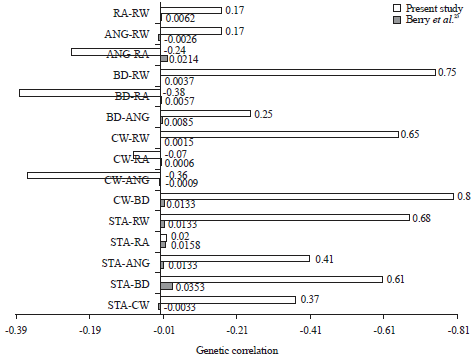 | |
| Fig. 4: | Genetic correlation compared with literature23 |
| ANG: Angularity, STA: Stature, BD: Body depth, RA: Rump angle, RW: Rump width, CW: Chest width | |
| Table 3: | Distribution of pedigree file |
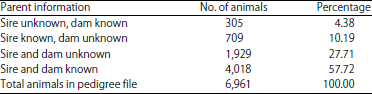 | |
| Table 4: | Estimated genetic variances (diagonal) and co-variances (off-diagonal) |
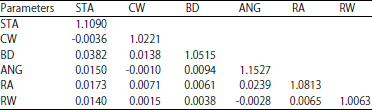 | |
| ANG: Angularity, STA: Stature, BD: Body depth, RA: Rump angle, RW: Rump width, CW: Chest width | |
CONCLUSION
The new approach of recording the linear traits by measuring directly the traits instead of scoring are possible with reasonable variability. The considerable magnitude of heritabilities indicated that the selected traits will respond to a selection program incorporating those linear traits. The small and nearly zero estimates of phenotypic and genetic correlation could be improved by using a more complete pedigree file in the analysis.
SIGNIFICANCE STATEMENT
This study discover the genetic parameters of selected linear traits that can be beneficial for the national breeding centre of dairy cows and forage (BBPTUHPT Baturraden) where the traits measurement were approached directly on the animals as opposed to scored. The genetic parameters of linear traits of dairy cows for this population have not been published in the literature and hence the results serve as the reference point for dairy cow breeder especially in Indonesia.
This study will help the researcher to uncover the genetic parameters of linear traits analyzed by animal model using Restricted Maximum Likelihood using pedigree data. The genetic parameters of linear traits are useful to predict selection response when selection program is applied to the population. The genetic correlation within linear traits under study can be used as reference when selective recording system needs to be operated. Until recently, the national breeding centre of dairy cows and forage (BBPTUHPT Baturraden) has not adopted the World Holstein Frisian Federation recommendation for recording linear trait in dairy cow breeding centre.
ACKNOWLEDGMENTS
The authors wish to thank the research grant for Professors and Doctors of Brawijaya University (Peraturan Rektor Universitas Brawijaya Nomor 15 Tahun 2019) for funding this research. Thanks are extended to the National Breeding Centre for Dairy Cattle and Forage of Indonesia (BBPTUHPT Baturraden) for providing data.
REFERENCES
- Royal, M.D., A.Q. Darwash, A.P.F. Flint, R. Webb, J.A. Woolliams and G.E. Lamming, 2000. Declining fertility in dairy cattle: Changes in traditional and endocrine parameters of fertility. Anim. Sci., 70: 487-501.
CrossRefPubMedDirect Link - Lucy, M.C., 2001. Reproductive loss in high-producing dairy cattle: Where will it end. J. Dairy Sci., 84: 1277-1293.
CrossRefDirect Link - Pryce, J.E., M.D. Royal, P.C. Garnsworthy and I.L. Mao, 2004. Fertility in the high-producing dairy cow. Livest. Prod. Sci., 86: 125-135.
CrossRefDirect Link - Oltenacu, P.A. and D.M. Broom, 2010. The impact of genetic selection for increased milk yield on the welfare of dairy cows. Anim. Welfare, 19: 39-49.
Direct Link - Ingvartsen, K.L., R.J. Dewhurst and N.C. Friggens, 2003. On the relationship between lactational performance and health: Is it yield or metabolic imbalance that cause production diseases in dairy cattle? A position paper. Livest. Prod. Sci., 83: 277-308.
CrossRefDirect Link - Oltenacu, P.A. and B. Algers, 2005. Selection for increased production and the welfare of dairy cows: Are new breeding goals needed? AMBIO: J. Hum. Environ., 34: 311-315.
CrossRefDirect Link - Harris, B.L., A.E. Freeman and E. Metzger, 1992. Genetic and phenotypic parameters for type and production in Guernsey dairy cows. J. Dairy Sci., 75: 1147-1153.
CrossRefDirect Link - Campos, R.V., J.A. Cobuci, E.L. Kern, C.N. Costa and C.M. McManus, 2015. Genetic parameters for linear type traits and milk, fat and protein production in Holstein cows in Brazil. Asian-Aust. J. Anim. Sci., 28: 476-484.
CrossRefPubMedDirect Link - Dal Zotto, R., M. de Marchi, C. Dalvit, M. Cassandro, L. Gallo, P. Carnier and G. Bittante, 2007. Heritabilities and genetic correlations of body condition score and calving interval with yield, somatic cell score and linear type traits in brown swiss cattle. J. Dairy Sci., 90: 5737-5743.
CrossRefDirect Link - Wall, E., I.M.S. White, M.P. Coffey and S. Brotherstone, 2005. The relationship between fertility, rump angle and selected type information in holstein-friesian cows. J. Dairy Sci., 88: 1521-1528.
CrossRefDirect Link - Zink, V., M. Štípková and J. Lassen, 2011. Genetic parameters for female fertility, locomotion, body condition score and linear type traits in Czech Holstein cattle. J. Dairy Sci., 94: 5176-5182.
CrossRefDirect Link - Yaeghoobi, R., A. Doosti, A.M. Noorian and A.M. Bahrami, 2011. Genetic parameters and trends of milk and fat yield in holstein's dairy cattle of West provinces of Iran. Int. J. Dairy Sci., 6: 142-149.
CrossRefDirect Link - Khorshidie, R., A.A. Shadparvar, N.G. Hossein-Zadeh and S.J. Shakalgurabi, 2012. Genetic trends for 305-day milk yield and persistency in Iranian Holsteins. Livest. Sci., 144: 211-217.
CrossRefDirect Link - Atabany, A., B.P. Purwanto, T. Toharmat and A. Anggraeni, 2013. Hubungan masa kosong dengan produktivitas pada sapi perah Friesian Holstein di Baturraden, Indonesia. Media Peternakan, 34: 77-82.
CrossRefDirect Link - Yazgan, K., 2018. RE-NUM-OR: Python-based renumbering and reordering software for pedigree files. Czech J. Anim. Sci., 63: 70-77.
CrossRefDirect Link - Toghiani, S., 2011. Genetic parameters and correlations among linear type traits in the first lactation of Holstein dairy cows. Afr. J. Biotechnol., 10: 1507-1510.
Direct Link - Nemcova, E., M. Stipkova and L. Zavadilova, 2011. Genetic parameters for linear type traits in Czech Holstein cattle. Czech J. Anim. Sci., 4: 157-162.
CrossRefDirect Link - Berry, D.P., F. Buckley, P. Dillon, R.D. Evans and R.F. Veerkamp, 2004. Genetic relationships among linear type traits, milk yield, body weight, fertility and somatic cell count in primiparous dairy cows. Irish J. Agric. Food Res., 43: 161-176.
Direct Link - White, J.M., 1974. Genetic parameters of conformational and managemental traits. J. Dairy Sci., 57: 1267-1278.
CrossRefDirect Link - Hayes, A.E. and I.L. Mao, 1987. Effects of parity, age and stage of lactation at classification on linear type scores of Holstein cattle. J. Dairy Sci., 70: 1898-1905.
CrossRefDirect Link - Mazza, S., C. Sartori and R. Mantovani, 2015. Genetic parameters of type traits in two strains of dual purpose autochthonous Valdostana cattle. Livest. Sci., 178: 35-42.
CrossRefDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Cantet, R.J., A.N. Birchmeier, M.G. Santos-Cristal and V.S. De Avila, 2000. Comparison of restricted maximum likelihood and method ℜ for estimating heritability and predicting breeding value under selection. J. Anim. Sci., 78: 2554-2560.
CrossRefDirect Link - Susanto, A., A.T.A. Sudewo and S.A. Santosa, 2013. Efek struktur pedigree populasi terhadap dugaan komponen variansi. In Seminar Nasional: Pengembangan Agribisnis Peternakan Menuju Swasembada Protein Hewani. Purwokerto: Fakultas Peternakan Universitas Jenderal Soedirman Bekerja Sama Dengan Ikatan Sarjana Peternakan Indonesia, pp: 283-287.
- Pryce, J.E., R.J. Esslemont, R. Thompson, R.F. Veerkamp and M.A. Kossaibati et al., 1998. Estimation of genetic parameters using health, fertility and production data from a management recording system for dairy cattle. Anim. Sci., 66: 577-584.
CrossRefDirect Link - Lund, T., F. Miglior, J.C.M. Dekkers and E.B. Burnside, 1994. Genetic relationships between clinical mastitis, somatic cell count and udder conformation in Danish Holsteins. Livest. Prod. Sci., 39: 243-251.
CrossRefDirect Link - Ali, T.E., E.B. Burnside and L.R. Schaeffer, 1984. Relationship between external body measurements and calving difficulties in Canadian Holstein-Friesian cattle. J. Dairy Sci., 67: 3034-3044.
CrossRefDirect Link - Royal, M.D., J.E. Pryce, J.A. Woolliams and A.P.F. Flint, 2002. The genetic relationship between commencement of luteal activity and calving interval, body condition score, production and linear type traits in Holstein-Friesian dairy cattle. J. Dairy Sci., 85: 3071-3080.
CrossRefPubMedDirect Link - Rogers, G.W., B.T. McDaniel, M.R. Dentine and D.A. Funk, 1989. Genetic correlations between survival and linear type traits measured in first lactation. J. Dairy Sci., 72: 523-527.
CrossRefDirect Link - Nilforooshan, M.A., A. Khazaeli and M.A. Edriss, 2008. Effects of missing pedigree information on dairy cattle genetic evaluations. Arch. Anim. Breed., 51: 99-110.
CrossRefDirect Link - Sanders, K., J. Bennewitz and E. Kalm, 2006. Wrong and missing sire information affects genetic gain in the angeln dairy cattle population. J. Dairy Sci., 89: 315-321.
CrossRefDirect Link