
P. X. Antony
Gandhi College of Veterinary and Animal Sciences, Pondicheny, India
G. K. Nair
College of Veterinary and Animal Sciences, Mannuthy, Kerala: India
V. Jayaprakasan
College of Veterinary and Animal Sciences, Mannuthy, Kerala: India
M. Mini
College of Veterinary and Animal Sciences, Mannuthy, Kerala: India
T. V. Aravindakshan
Gandhi College of Veterinary and Animal Sciences, Pondicheny, India
International Journal of Dairy Science
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 178-182







ABSTRACT
Knowledge of the serotype of Pasteurella multocida (P. multocida) involved in an outbreak is essential in order to constitute effective control measures. Antigenic characterization of P. multocida is accomplished by capsular serogrouping and somatic serotyping. These typing techniques are carried out by reference laboratories only. This often leads to delay in knowing the serotype of the isolate. DNA based techniques for differentiation of serotypes can provide an alternative to conventional serotyping. A PCR-REA technique was standardized to distinguish P. multocida serotypes A:1, A:3 and B:2
PDF Abstract XML References
How to cite this article
P. X. Antony, G. K. Nair, V. Jayaprakasan, M. Mini and T. V. Aravindakshan, 2007. Nucleic Acid Based Differentiation of Pasteurella multocida Serotypes. International Journal of Dairy Science, 2: 178-182.
DOI: 10.3923/ijds.2007.178.182
URL: https://scialert.net/abstract/?doi=ijds.2007.178.182
DOI: 10.3923/ijds.2007.178.182
URL: https://scialert.net/abstract/?doi=ijds.2007.178.182
INTRODUCTION
Pasteurella multocida has been recognized as an important Veterinary pathogen for over a century. The organism can occur as a commensal in the naso-pharyngeal region of apparently healthy animals and it can be either a primary or secondary pathogen in the disease processes of a variety of domestic and feral mammals and birds.
Conventional methods for diagnosis of pasteurellosis rely on the detection of the organism by microscopy and its isolation and identification. However as far as pasteurellosis is concerned it is not just sufficient to know the identity of the organism. To constitute effective control measures it is important to know the serotype of the organism. Typing of P. multocida is principally accomplished via passive haemagglutination and gel diffusion precipitin tests. Isolates are classified into five groups according to their capsular antigens and into 16 serotypes on the basis of their lipopolysaccharide antigens (Huber et al., 2002).
Serotyping of P. multocida is currently only undertaken by regional reference laboratories. A limitation of the capsule typing is the difficulty in inducing antibodies to specific antigens. Most workers find it relatively easy to make antibodies against B and E serogroup specific antigens, but not the other serogroup specific antigens. Many times a non-encapsulated strain has not been serotypeable (Rimler and Rhoades, 1989).
Recently a multiplex Polymerase Chain Reaction was introduced as a rapid alternative to capsular serotyping system (Townsend et al., 2001). However by this technique only the capsular serotyping information could be ascertained. Gautam et al. (2004) introduced a PCR technique specific for P. multocida serogroup A.
The PCR-Restriction fragment length polymorphism (PCR-RFLP) has been used by several workers to detect polymorphism within a gene segment and such information has been useful for serotyping of isolates.
Heinemann et al. (2000) investigated the use of PCR-RFLP for differentiation of serovars using 19 different restriction endonucleases and concluded that PCR-RFLP could be used as a rapid detection technique for differentiation of Leptospira sp. serovars.
Restriction endonuclease analysis with Hpa II of 218 isolates of P. multocida from swine revealed 17 patterns. The authors opined that REA was a useful epidemiological tool for identifying different strains of P. multocida (Rubies et al., 2002).
Hirose et al. (2003) while studying the antibiotic resistance patterns of Salmonella enterica serovar typhi and serovar Paratyphi to fluoroquinolones demonstrated the emergence of strains with mutations in the gyrA genes, using PCR-RFLP.
Hong et al. (2003) used a PCR-RFLP flagellar typing scheme to successfully serotype 112 Salmonella isolates obtained from poultry. Restriction enzymes Sau3A and Hha I were used for the purpose. They concluded that PCR-RFLP was a rapid, accurate and economical approach for serotyping Salmonella species.
Until now no PCR-RFLP based techniques have been used for determination of both the capsular and somatic serotyping of P. multocida. Moreover the problems encountered in P. multocida serotyping system does not affect DNA fingerprinting. Hence the present study was undertaken to develop a simple DNA based typing scheme.
MATERIALS AND METHODS
Pasteurella multocida Strains
Pasteurella multocida strain DP1 and FP1 isolated in Kerala, India from a duck and fowl respectively and serotyped as A:1 by Indian Veterinary Research Institute, Izatnagar, India and Pasteurella multocida serotypes A:3, B:2 and a duck isolate LKO, serotype A:1 obtained from Indian Veterinary Research Institute, Izatnagar, India, formed the reference strains for this study.
Oligonucleotide Primers
Two oligonucleotides based on the sequence of P. multocida X-73 OmpH gene, Accession No. U50907 (Luo et al., 1997) were designed using Primer3 software. The primers were custom synthesized by M/s Bangalore Genei India. The sequences of the two primers were as follows:
|
| OmpH 1 5’-GCG TTT CAT TCA AAG CAT CTC-3’ - 21 mer and OmpH 2 5’-ATG ACC GCG TAA CGA CTT TC -3’- 20 mer |
Amplification of OmpH gene
PCR Conditions
A 50 μL reaction mixture was prepared in 0.2 mL thin walled PCR tube. Five microlitres of template DNA was added to a reaction mixture containing 40 pmoles each of primer OmpH 1 and OmpH 2, 200 μM of each dNTP, 1 X Taq buffer with 1.5 M MgCl2 and 2 units of Taq DNA polymerase. The amplification reaction was carried out in an automated thermal cycler (Eppendorf Master Cycler, Germany) according to the following program, an initial denaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 15 sec, annealing at 56°C for 1 min, extension at 72°C for 1 min and a final extension at 72°C for 10 min. Bacterial culture lysates was used as template DNA. The product was analysed by 2% agarose gel electrophoresis. Standard molecular size marker low range DNA ruler with fragments 3000, 2500, 2000, 1500, 1000, 600, 300 and 100 bp was used as DNA molecular size marker to ascertain the size of the amplified PCR product.
Restriction Enzyme Analysis of OmpH-PCR Product
The amplified PCR products were subjected to restriction enzyme digestion using restriction enzymes Dra I and Hinf I. The digestion was carried out by a standard method (Sambrook et al., 1989). Briefly restriction enzyme digestion was carried out at 37°C for two h, followed by inactivation of the enzyme at 80°C for 20 min. Electrophoresis of the resultant digest was conducted on 8% acrylamide gels. The gel was viewed on a transilluminator and photographed.
RESULTS
The primer pairs OmpH 1 and OmpH 2, designed to amplify the OmpH gene of P. multocida successfully amplified the OmpH gene of all three serotypes viz., A:1, A:3 and B:2. The amplified product had a size of approximately 1000 bp (Fig. 1).
Specificity of the Primers
Primer pairs OmpH 1 and OmpH 2 did not amplify the DNA prepared from unrelated bacterial species such as Streptococcus zooepidemicus, Staphylococcus aureus, Escherichia coli, Klebsiella pneumoniae, Leptospira serogroup canicola serovar canicola.
Restriction Enzyme Analysis
Restriction analysis of the amplified products of serotypes A:1, A:3 and B:2 were carried out using the same restriction endonucleases Dra I and Hinf I. Enzyme Hinf I generated patterns similar in A:3 and B:2 but distinct from A:1, while Dra I profiles were distinct from each other for the three serotypes (Fig. 2).
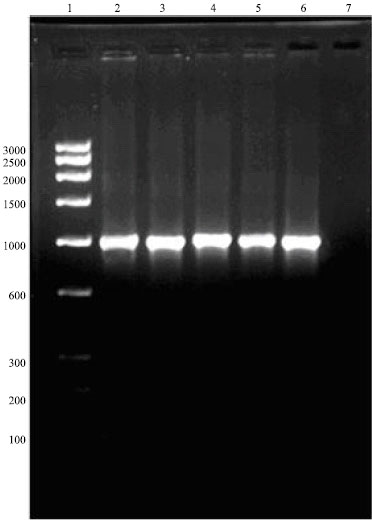 | |
| Fig. 1: | Amplification of OmpH gene of P. multocida Lane 1 Low ragne DNA marker, Lane 2-3 LKO and FPI, Lane 4 DPI, Lane 5 A:3, Lane 6 B:2, Lane 7 Negative control |
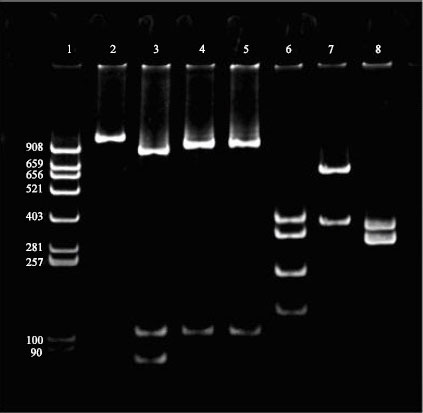 | |
| Fig. 2: | Restriction enzyme profiles of OmpH-PCR products (scrotypes A:1, A:3 and B:2) with Hinf I and Dra I, Lane 1 pBR 322 DNA/Alu digest marker, Lane 2 Undigested OmpH-PCR product, Lane 3-5 A:1, A;3 and B:2 (Hinf I digest), Lane 6-8 A:1, A;3 and B:2 (Dra I digest) |
DISCUSSION
The custom designed primers could successfully amplify the OmpH gene of three serotypes viz., A:1, A:3 and B:2. Similar results were reported by Luo et al. (1999) who had reported successful amplification of OmpH genes of the serotypes 1, 3 and 4 to 16 of P. multocida and the product had an approximate size of 1 kilo-base pair.
REA of amplified products of OmpH-PCR with Dra I generated distinct profiles for the three serotypes A:1, A:3 and B:2 while Enzyme Hinf I generated patterns similar in A:3 and B:2 but distinct from A:1. Thus, REA of amplified products of OmpH-PCR with Dra I offers a novel technique for differentiation of various serotypes of P. multocida. If unique patterns for all serotypes could be determined in a similar manner then we can have a serotyping technique that is simple, rapid and straight forward and can be performed in any laboratory that has the capacity to perform PCR. The development of a DNA-based technique for differentiation of serotypes could provide an alternative to conventional serotyping systems and has a potential to overcome the problems associated with the current serotyping techniques, which rely on inconsistent expression of phenotypic traits.
However, further studies have to be carried out with all the different serotypes to know whether profiles unique to each serotype are obtained, before the technique can be put for routine use. To the best of our knowledge this is the first report of the use of PCR-RFLP for differentiation of P. multocida serotypes.
ACKNOWLEDGMENTS
The authors are grateful to the Indian Council of Agricultural Research, New Delhi, India for providing financial support under the All India Network Programme on Haemorrhagic Septicaemia and the Dean, College of Veterinary and Animal Sciences, Mannuthy, Kerala, India for providing facilities to conduct this study. The first author also expresses his sincere thanks to the Dean. Rajiv Gandhi college of Veterinary and Animal Sciences, Pondicherry, for his keen interest and valuable suggestion during the course of this study.
REFERENCES
- Gautam, K., A.A. Kumar, V.P. Singh, V.P. Singh, T.K. Dutta and S.B. Shivachandra, 2004. Specific identification of Pasteurella multocida serogroup A isolates by PCR assay. Res. Vet. Sci., 76: 179-185.
PubMedDirect Link - Heinemann, M.B., J.F. Garcia, C.M. Nunes, F. Gregori, Z.M.M. Higa, S.A. Vasconcellos and L.J. Richtzenhain, 2000. Detection and differentiation of Leptospira spp. serovars in bovine semen by polymerase chain reaction and restriction fragment length polymorphism. Vet. Microbiol., 73: 261-267.
CrossRefDirect Link - Hirose, K., K. Tamura and H. Watanabe, 2003. Screening method for Salmonella enteriaM serovar Typhi and Serovar Paratyphi a with reduced susceptibility to fluroquinolones by PCR restriction fragment length polymorphism. Microbiol. Immunol., 47: 161-165.
Direct Link - Hong, Y., T. Liu, C. Hofacre, M. Maier, D.G. White, S. Ayers, L. Wang and J.J. Maurer, 2003. A restriction fragment length polymorphism-based polymerse chain reaction as an alternative to serotyping for identifying Salmonella serotypes. Avian Dis., 47: 387-395.
Direct Link - Huber, B.S., D.A. Allred, J.C. Carmen, D.D. Frame and D.G. Whiting et al., 2002. Random amplified polymorphic DNA and amplified fragment length polymorphism analyses of Pasteurella multocida isolates from fatal fowl cholera infections. J. Clin. Microbiol., 40: 2163-2168.
CrossRefDirect Link - Luo, Y., J.R. Glisson, M.W. Jackwood, R.E. Hancock, M. Bains, I.N. Cheng and C. Wang, 1997. Cloning and characterization of the major outer membrane protein gene (OmpH) of Pasteurella multocida X-73. J. Bacteriol., 179: 7856-7864.
Direct Link - Luo, Y., Q. Zeng, J.R. Glisson, M.W. Jackwood, I.H.N. Cheng and C. Wang, 1999. Sequence analysis of Pasteurella multocida major outer membrane protein (OmpH) and application of synthetic peptides in vaccination of chickens against homologous strain challenge. Vaccine, 17: 821-831.
CrossRefDirect Link - Rubies, X., J. Casal and C. Pijoan, 2002. Plasmid and restriction endonuclease patterns in Pasteurella multocida isolated from a swine pyramid. Vet. Microbiol., 84: 69-78.
CrossRef - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 1st Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Direct Link - Townsend, K.M., J.D. Boyce, J.Y. Chung, A.J. Frost and B. Adler, 2001. Genetic organization of Pasteurella multocida cap loci and development of a multiplex capsular PCR typing system. J. Clin. Microbiol., 39: 924-929.
CrossRefPubMedDirect Link