
Zohreh Moradi
Department of Plant Pathology, College of Agriculture, Ferdowsi University of Mashhad, Khorasan, Iran
Asian Journal of Plant Pathology
Year: 2011 | Volume: 5 | Issue: 3 | Page No.: 115-125







ABSTRACT
Watermelon Mosaic Virus (WMV) is a Potyvirus with a worldwide distribution. This virus causes serious economic losses in many cucurbits in Iran. To study relative incidence and molecular variability of WMV, 620 samples of cucurbit plants were collected from the different fields and cucumber greenhouses in some regions of Iran. Three hundred and three samples were infected by WMV in DAS-ELISA using specific polyclonal antibody (The rate of infection was 48.8%). In RT-PCR assay with specific primers of WMV, a single band of about 825 bp in length was produced from the samples. The Coat Protein (CP) region of the genome from 15 representative isolates were sequenced and compared with the sequences available in GeneBank. The identity of WMV CP nucleotide sequences of the 15 Iranian isolates ranged from 95.8 to 99.2%. According to this research Iranian isolates have high variation in CP gene. In most cases, geographical isolation is consistent with the phylogenetic grouping. Molecular weights of coat protein using SDS-PAGE were estimated at 34100 Da. Data shown in this study represent the occurrence of WMV based on ELIZA, RT-PCR and CP gene analyses in some region of Iran. These results make it possible to have a better understanding of the development of WMV disease in Iran.
PDF Abstract XML References Citation
Received: April 15, 2011;
Accepted: July 26, 2011;
Published: September 26, 2011
How to cite this article
Zohreh Moradi, 2011. Diagnosis and Molecular Variability of Watermelon mosaic virus Isolates from North, East, North-east and North-west Regions of Iran. Asian Journal of Plant Pathology, 5: 115-125.
URL: https://scialert.net/abstract/?doi=ajppaj.2011.115.125
URL: https://scialert.net/abstract/?doi=ajppaj.2011.115.125
INTRODUCTION
Watermelon Mosaic Virus (WMV) is a member of the genus Potyvirus (family Potyviridae) and consists of flexuous, filamentous particles, approximately 760 nm long. The genome is a positive-sense, single-stranded RNA and the size of RNA is about 10035-nt long. The genomes of Potyviridae have a single Open Reading Frame (ORF) that is translated into a single large polyprotein which is hydrolyzed, after translation, into several proteins by virus-encoded proteinases (Riechmann et al., 1992; Al-Saleh et al., 2009).
At the molecular level, WMV is closely related to SMV (Soybean mosaic virus) in most of its genome (in spite of its different and much broader host range) and could consider as a divergent strain of this virus. Moreover, it also appears to share a recombination with Bean common mosaic Virus (BCMV) in the P1 coding region. The first report of full sequenced WMV was from France (WMV-Fr) (Desbiez and Lecoq, 2004) followed by China (WMV-CHN) (Wu et al., 2006) and Pakistan (WMV-PK) (Ali et al., 2006). WMV is probably distributed worldwide mostly in temperate regions and was first reported in Citrullus lanatus by Webb and Scott (1965) and Purcifull et al. (1984). The geographical occurrence of the virus is variable and depends on climatic conditions (Desbiez et al., 2007).
The Cucurbitaceae family includes several species of horticultural crops. Iran cultivates the second-largest total cucurbit acreage (770,000 ha) in the world, exceeded only by China. (Massumi et al., 2007). WMV can cause economically important diseases in several horticultural crops, mostly cucurbits and legumes, resulting in quality and yield losses. It can experimentally infect more than 170 plant species belonging to 27 families, including many weeds that can host the virus between crops (Shukla et al., 1994). In Iran, host range of this virus is limited to cucurbitaceous plants but in the East also other plants such as Habenaria and Vanilla have been reported as host (Gara et al., 1997; Wang et al., 1993).
The studies of WMV variation at the nucleotide and amino acid levels focused on analysis of the sequence of the P1, CP and Cylindrical Inclusion (CI) genes (Sharifi et al., 2008). The first report of WMV genetic diversity in Iran was related to Desbiez et al. (2007) that reported a 218-nucleotide sequence of the CP N-terminal region of eight isolates from Iran. Sharifi et al. (2008) analyzed 18 isolates from central and southern regions of Iran based on CP gene. Following Shoeibi et al. (2009) analyzed 4 isolates from Golestan province and one isolate from Mashhad and Shiraz (from north east and southern regions of Iran, respectively).
Although, WMV has long been known to occur in several regions of Iran (Sharifi et al., 2008), there are few reports on its occurrence in North, East, Northeast and Northwest of Iran. The objective of the present study was amplification of CP gene and evaluation of the genetic variation among the CP sequences of the chosen isolates collected from several regions of Iran. Information about geographically diverse range of WMV isolates encouraged to originate facts on the molecular variability of this virus.
MATERIALS AND METHODS
Viral sample sources: WMV isolates were collected from different geographical locations in the northwest (provinces of East Azarbayjane, West Azarbayjane and Ardabil), north (provinces of Mazandaran and Golestan), Northeast and East (provinces of Razavi Khorasan, Northern Khorasan and South Khorasan) of Iran (Table 1), from June 2009 to October 2011. Samples (n = 620) consisted of young fully expanded leaves showing Symptoms such as systemic mosaic, severe yellow mosaic, vein clearing and blistering and including cucumber, Cucurbita pepo varieties, watermelon (Citrullus lanatus) and melon (Cucumis melo varieties including Cantaloupe, snakemelon, longmelon and local cvs.). The samples were stored at 80°C until they were used.
| Table 1: | Accession Number, Origin, name of isolates and host of 15 sequenced isolates of Watermelon mosaic virus |
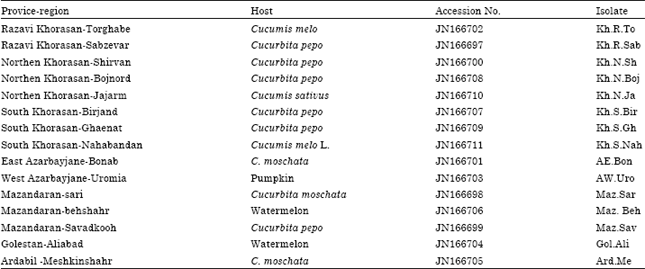 | |
Double-antibody Sandwich (DAS)-ELISA: DAS-ELISA was performed using the specific polyclonal antibody (SEDIAG S.A, Strasbourg, France) following the general protocol of Clark and Adams (1977). Extracts from healthy C. pepo were used as negative control in all tests.
Virus isolation and propagation: Virus isolation biologically was carried out using single local lesion assay following two passages onto Chenopodium amaranticolor CosteandReyn. Then virus was propagated in C. melo and Cucurbita pepo (squash cvs. Black knight and Local) for further investigation. ELISA positive samples were homogenized in 50 mM potassium phosphate buffer (pH 7.0) and the extract was rubbed onto leaves dusted with carborundum powder. Plants were maintained in an insect-proof screen house at 24-27°C and controlled at least for 4 weeks after inoculation. ELISA test was used addition to symptoms observation for checking the presence of the virus.
Protein analysis: The molecular weight of the viral coat protein was estimated by discontinuous SDS-PAGE (Sodium dodecyl sulfate poly acryl amid gel electrophoresis) as described by Laemmli (1970). A purified virion suspension and plant samples infected with the virus were mixed with the sample buffer, boiled at 100°C for 5 min and subjected to electrophoresis on 12% polyacrylamide gel along with poly peptide size standard. The gel was stained with coomassie blue. The molecular weight of polypeptides from WMV was determined by comparison with the size standards.
Total RNA extraction and RT-PCR: Total RNA was extracted from pulverized tissue using AccuZol™ Reagent (Bioneer, Alameda, CA) from fresh, systemically infected cucurbit leaves according to the manufacturer’s instructions. After precipitating with ethanol, total RNA was re-solubilized in 25 μL of RNase-free water. RNA samples were tested for the presence of WMV using specific primers designed to amplify a fragment of the coat protein gene. Forward primer (5'- GAA TCA GTG TCT CTG CAA TCA GG -3') and reverse primer (5'- ATT CAC GTC CCT TGC AGT GTG -3') (Sharifi et al., 2008) corresponding to nucleotides (nt) 8926 to 8948 and nt 9727 to 9747 of WMV-Fr (GenBank accession number EU660584), respectively, were used to amplify a 825-bp fragment covering the CP region. Reverse Transcription (RT) reaction was performed as follows: 1 μL of reverse primer (20 picomoles) and 1 μL of RNA sample were added to 8.5 μL of Diethyl Pyrocarbonate (DEPC)-treated water. The mixture was incubated at 65°C for 10 min and chilled on ice for 3 min to denature the RNA. Then 3.5 μL of DEPC-treated water, 4 μL of 5x M-MLV RT buffer, 2 μL of dNTPs mix (10 mm) and 1 μL M-MLV (200 U μL) reverse transcriptase (Promege, USA) were added to mixture. The RT reactions were incubated at 42°C for 60 min followed by 95°C for 3 min to terminate the RT reaction. Viral cDNA was then amplified by PCR. The PCR reaction was performed using 2.5 μL of cDNA, 13.5 μL DEPC-treated water, 5 μL of 5x GoTag polymerase buffer, 2.5 μL 10x MgCl2, 0.5 μL of each forward and reverse primers (20 pmol), 0.75 μL of dNTP mix (10 mm) and 0.125 μL of GoTag polymerase (2.5 U μL) (Promege, USA). Following program was used for PCR: A first denaturation for 3 min at 94°C was performed and followed by 35 cycle of denaturation for 60 s at 94°C, 60 sec of annealing at 63°C, extension for 1 min at 72°C and a final extension step at 72°C for 10 min. PCR products were analyzed by electrophoresis in 1.7% agarose gel and visualized by ethidium bromide staining.
| Table 2: | GenBank accession numbers and origin of previously reported WMV Isolates/strainsa used for phylogenetic comparison of the 789 nucleotides long fragment of genome including CP gene |
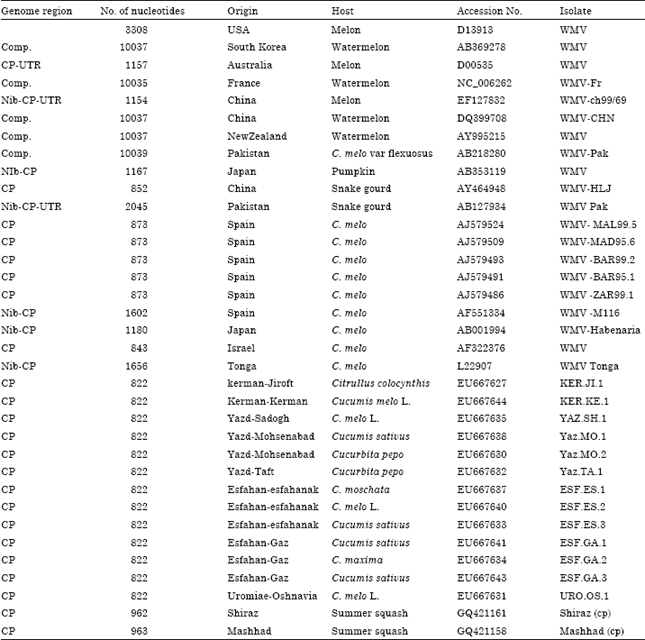 | |
| a: The WMV isolates were identified by location, plant host and series number | |
Sequencing and phylogenetic analysis: RT-PCR products of WMV isolates were then purified from 1% agarose gels using the DNA gel extraction kit (bioneer, korea) and nucleotide sequencing reactions were performed by MWG Company (Biotech, Germany) using dideoxy nucleotide chain termination method with specific forward and reverse primers. Then, these and 34 isolates from the GenBank were analyzed to determine their phylogenetic relationships.
The phylogenetic analysis of the Iranian isolates was conducted by comparing the 789 bp of the CP gene with those of the other isolates of WMV obtained from GenBank (Table 2). Nucleotide sequence analysis and translation to the corresponding amino acid sequence were performed using DNAMAN software (version 4.02) package (BBA, Germany). The nucleotide and deduced amino acid sequences were compared with the equivalent sequences of WMV. Multiple alignments of the nucleotide and amino acid sequences were carried out using the ClustalW program in BioEdit (version 7.0.9.) software and DNAMAN software (version 4.02). Phylogenetic tree for grouping based on nucleotide sequences were constructed by MEGA 4.1 software program using the neighbor-joining method. Tree branches were bootstrapped with 1000 replications and bootstrap values less than 55% were condensed on nodes (Fig. 4).
RESULTS
Virus isolate: Among 620 samples collected from different areas of Iran, 303 samples were positive in ELISA test (The rate of infection was 48.8%). The symptoms consisted of varying degrees of mosaic with leaf blistering and leaf deformation or stunting. Symptoms were diverse in different Cucurbit plants and related to host plant age and time of infection (Fig. 1a-d).
SDS-PAGE: The size of the viral coat protein of some of the isolates was determined about 34,100 Da while no band was found in the healthy plant extracts (Fig. 2) and this was similar to standard protein molecular weight respect to other WMV isolates described by researchers in other parts of the world.
RT-PCR, sequence data and identity matrix: RT-PCR was carried out using the primers WMV/F and WMV/R (described previously) which resulted in a fragment of approximately 825 nts (Fig. 3). Sequence information for these 15 isolates has been submitted to NCBI-GenBank with the accession numbers from JN166697 to JN166711 (Table 1).
 | |
| Fig. 1(a-d): | WMV Symptoms on different cucurbit plants. (a) Mosaic, blistering, vein banding and malformation of leaves on zucchini, (b) Mosaic and blistering on C. melo., (c) Yellowing, blistering and malformation on C. sativus and (d) Sever systemic mosaic on Citrullus lanatus leaf blade |
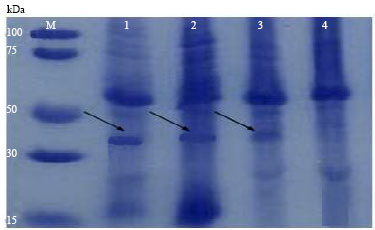 | |
| Fig. 2: | Denaturing polyacrylamide gel electrophoresis of infected plants (line 1, 2, 3) related to Khorasan provinces isolates, healthy plant (zucchini leaf) (4) and molecular weight marker proteins (M). The protein band in gel was stained with Coomassie Blue |
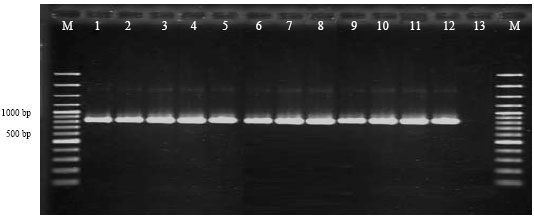 | |
| Fig. 3: | Electrophoresis pattern of DNA fragments amplified by RT-PCR in 1.7% agarose gel related to twelve selected WMV isolates collected from different location of Iran (lanes 1-12) 13: healthy Zucchini plant extract as negative control. M: 100 bp DNA marker (Fermentas) |
Phylogenetic analysis: Phylogenetic tree based on multiple sequence alignment of CP divided all WMV isolates into two large groups: group 1 included 4 subgroups: Sg1, Sg2, Sg3, Sg4 (Fig. 4). Group 1 included Middle East and European isolates, only one isolate from Australia placed in this group. Most of Iranian isolates were classified in the subgroup 1 which can be divided into many clades. 13 out of 15 isolates including Kh.R.To, Kh.R.Sab (collected from Razavi Khorasan province), Kh.N.Sh, Kh.N.Boj, Kh.N.Ja (collected from Northern Khorasan province), Kh.S.Bir, Kh.S.Gh, Kh.S.Nah (collected from South Khorasan province), AE.Bon (collected from East Azarbayjane province), Maz.Sar Maz. Beh, Maz.Sav (collected from Mazandaran province), Ard.Me (collected from Ardabil province) have been reported for the first time herein. Pakistan isolates were classified in subgroup 2. Geographically, only Subgroup 3 is a heterogeneous group and included isolates of center parts of Iran (Yazd, KERJI1), Tonga, USA and New Zealand. Exception Shiraz isolate in subgroup 4 which has been located with Japan isolates, geographical isolation is consistent with the phylogenetic grouping (Shoeibi et al., 2009).
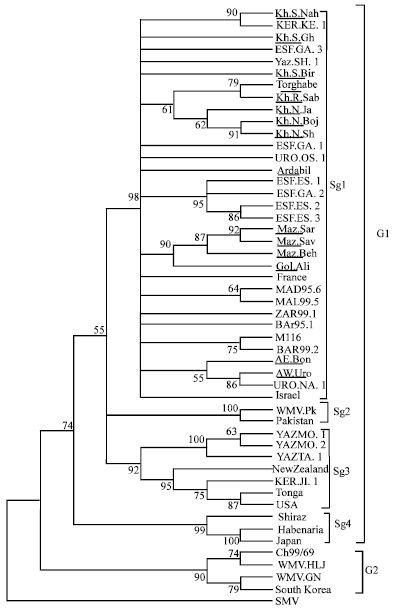 | |
| Fig. 4: | Phylogenetic tree constructed from the alignment of nucleotide sequences of coat protein gene of 45 WMV isolates (Table 1 and 2) using neighbor-joining method based on 1000 replicates. The numbers indicate bootstrap percentage. Bootstrap values higher than 55 are indicated on nodes and the nodes less than 55 were condensed. The isolates which have been sequenced here, were underlined. SMV (Soybean mosaic virus) included as out-group |
Isolates in group 2 included South Korea, Poland and China isolates. Eighteen isolates in subgroup 1 were collected from different regions of Khorasan provinces formed a distinct small group according to the geographical region.
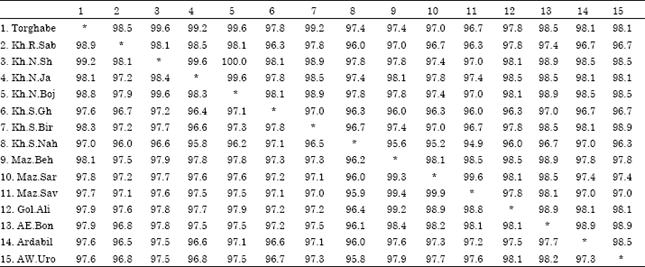
Phylogenetic analyses based on ClustalW Multiple alignments, demonstrated that 15 isolates share 95.8 to 99.2% nucleotide sequence identity with each other. Identity of 15 Iranian isolates in the amino acid levels ranged between 95.2 to 100% (Table 3). The percentage similarities for nucleotide between Iranian isolates and the other isolates of WMV recorded in NCBI ranged from 91 to 99%. The DAG box (Asp-Ala-Gly) which is critical for Potyvirus transmission by aphids, was detected in the all Iranian isolates coat protein gene.
| Table 3: | Comparison of the Percent nucleotide (bottom and left, in bold) and amino acid (top and right) identities of 825 nt CP sequences between selected WMV isolates used in this study |
 | |
DISCUSSION
Virus infections are a major limiting factor in cucurbit production in Iran (Moradi and Jafarpour, 2011). Watermelon Mosaic Virus (WMV) causes serious economic losses in Iran in many cucurbits. Cucurbitaceous crops are grown commercially throughout the world, have been destroyed completely locally or epidemically when infected by plant viruses (Yoon et al., 2008).
The RT-PCR technique is highly sensitive and is found to be a reliable method in comparison to biological indexing and DAS-ELISA. Using RT-PCR the virus could be detected in some plants which were found to be negative on DAS-ELISA (Sidaros et al., 2009). Also, PCR became widely used as a diagnostic method for infection by plant viruses belonging to several different groups such as the geminiviruses, luteoviruses and potyvirus groups (Al-Saleh et al., 2010; Sanchez et al., 2007).
The CP gene is the gene most frequently used for studies of genetic diversity in Potyviruses (El-Araby et al., 2009). Most of the genes that used to identify WMV were using the Polyprotein (P1) gene or the Coat Protein CP) gene (Salem et al., 2007) and hence, in the study, CP gene sequences were used for comparison and analysis.
Among different cucurbit plants, zucchini squash and pumpkin has shown the highest percentage of WMV infection and the lowest of WMV infection is related to cucumbers (data not shown). The most typical symptoms in zucchini squash were observed with systemic mosaic and varying degrees of blistering on leaves and this was in agreement with Shoeibi et al. (2009). In many cases the symptoms mix with other cucurbit infecting viruses (Moradi and Jafarpour, 2011).
According to the results of this study and those of Sharifi et al. (2008) the isolates of group 1 are the most widespread isolates of WMV and maximum molecular variation occurs in Eastern Asia (Shoeibi et al., 2009). The deduced coat protein consisted of 261 amino acids and also, all Iranian isolates had a DAG amino acid triplet which is required in aphid transmissibility this in agreement with Sharifi et al. (2008).
Data shown in this study represent the occurrence of WMV based on ELIZA, RT-PCR and CP gene analyses in some region of Iran. Previous studies were in the central and southern regions (Sharifi et al., 2008) but did not provide information about it in northwest, north, east and northeast of Iran at the molecular level.
Phylogenetic analyses were done by the Neighbor-joining (NJ) method implemented with ClustalW alignment which was used to compare the similarities between the coding nucleotides and their encoded protein. Sequencing results of a coat portion confirmed the RT-PCR analysis and Slight variations were observed between the results obtained in our study with those obtained in previous investigations. The minor differences observed in the nucleotide sequence homology between the Iranian isolates may suggest that all these isolates are closely related. In most cases, geographical isolation is consistent with the phylogenetic grouping.
Plant pathogenic viruses are responsible for increasing economic losses worldwide. There is no chemical treatment to eliminate a plant of virus infection (Youssef and Shalaby, 2009); however, chemotherapy directed to plant viral diseases has evolved significantly. So, identification and analysis of its genetic diversity of isolates and strains of WMV, presenting the appropriate solution to its control.
Determining variability within a virus group and understanding mechanism and factors affecting this variability are of considerable agronomic significance, particularly for determining resistance gene deployment strategies, since natural resistance genes can be rapidly overcome by adapted virus strains. In addition, variability of virus strain, particularly within the capsid protein, raises a problem for the development of reliable diagnosis techniques based on the antigenic properties of the coat protein (Safaeizadeh, 2008). These results make it possible to have a better understanding of the development of WMV disease in Iran.
CONCLUSION
The present study is one of several investigations regarding the significance of pathogenicity of this virus and aims at isolation, biological and molecular characterization of Iranian isolates of WMV which were isolated from cucurbit plants. In summary, the nucleotide sequences of CP gene of 15 WMV isolates was determined and demonstrated their identity and phylogenetic relationships to the other isolates of WMV recorded in NCBI. The identity of WMV CP nucleotide sequences of the 15 Iranian isolates ranged from 95.8 to 99.2%. The results obtained from this study will help managing this disease in cucurbit crops and probably in other crops that were reported to be infected with it, as well.
ACKNOWLEDGMENTS
The author wish to thank plant pathology laboratory manager at the Ferdowsi University of Mashhad. Also, the author is grateful to Dr. L. Prevotat for the WMV antiserum and the help of plant virologist, Dr. M. Mehrvar and those who help in technical works.
REFERENCES
- Ali, A., T. Natsuaki and S. Okuda, 2006. The complete nucleotide sequence of a Pakistani isolate of Watermelon mosaic virus provides further insights into the taxonomic status in the Bean common mosaic virus subgroup. Virus Genes, 32: 307-311.
PubMed - Al-Saleh, M.A., I.M. Al-Shahwan, O.A. Abdalla and M.A. Amer, 2010. Identification and coat protein nucleotide sequence of turnip mosaic potyvirus from Eruca sativa in Saudi Arabia. Asian J. Plant Pathol., 4: 116-127.
Direct Link - Al-Saleh, M.A., I.M. Al-Shahwan, M.A. Amer and O.A. Abdalla, 2009. Etiology of a mosaic disease of radish and lettuce and sequencing of the coat protein gene of the causal agent in Saudi Arabia. Int. J. Virol., 5: 131-142.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Desbiez, C., C. Costa, C. Wipf-Scheibel, M. Girard and H. Lecoq, 2007. Serological and molecular variability of watermelon mosaic virus (genus Potyvirus). Arch. Virol., 152: 775-781.
CrossRefDirect Link - Desbiez, C. and H. Lecoq, 2004. The nucleotide sequence of Watermelon mosaic virus (WMV, Potyvirus) reveals interspecific recombination between two related potyviruses in the 5' part of the genome Arch. Virol., 149: 1619-1632.
CrossRefDirect Link - El-Araby, W.S., I.A. Ibrahim, A.A. Hemeida, A. Mahmoud, A.M. Soliman, A.K. El-Attar and H.M. Mazyad, 2009. Biological, serological and molecular diagnosis of three major potato viruses in Egypt. Int. J. Virol., 5: 77-88.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Massumi, H., A. Samei, A.H. Pour, M. Shaabanian and H. Rahimian, 2007. Occurrence, distribution and relative incidence of seven viruses infecting greenhouse-grown cucurbits in Iran. Plant Dis., 91: 159-163.
CrossRef - Moradi, Z. and B. Jafarpour, 2011. First report of coat protein sequence of Cucumber green mottle virus in cucumber isolated from Khorasan in Iran. Int. J. Virol., 7: 1-12.
CrossRef - Riechmann, J.L., S. Lain and J.A. Garcia, 1992. Highlights and prospects of potyvirus molecular biology. J. Gen. Virol., 73: 1-16.
Direct Link - Salem, T.Z., S.M. El-Gamal and A.S. Sadik, 2007. Use of helper component proteinase gene to identify a new Egyptian isolate of Watermelon mosaic potyvirus. Int. J. Virol., 3: 107-116.
Direct Link - Safaeizadeh, M., 2008. Comparative biological and molecular variability of Zucchini yellow mosaic virus in Iran. Asian J. Plant Pathol., 2: 30-39.
CrossRefDirect Link - Sanchez, F., M. Rodriguez-Mateos, A. Tourino, J. Fresno and C. Gomez-Campo et al., 2007. Identification of new isolates of Turnip mosaic virus that cluster with less common viral strains. Arch. Virol., 152: 1061-1068.
CrossRefDirect Link - Sharifi, M., H. Massumi, J. Heydarnejad, A.H. Pour, M. Shaabanian and H. Rahimian, 2008. Analysis of the biological and molecular variability of Watermelon mosaic virus isolates from Iran. Virus Genes, 37: 304-313.
CrossRefPubMedDirect Link - Sidaros, S.A., S.A. El-Kewey, H.A. Amin, E.A.H. Khatab, A.A. Emeran, S.A. El-Khalik and M.A.S. El-Kady, 2009. Cloning and sequencing of a cDNA encoding the coat protein of an Egyptian isolate of pepper mild mottle virus. Int. J. Virol., 5: 109-118.
CrossRefDirect Link - Wang, Y.Y., D.L. Beck, R.C. Gardner and M.N. Pearson, 1993. Nucleotide sequence, serology and symptomatology suggest that vanilla necrosis potyvirus is a strain of Watermelon mosaic virus II. Arch. Virol., 129: 93-103.
CrossRefPubMedDirect Link - Youssef, S.A. and A.A. Shalaby, 2009. Single-step multiplex reverse transcription-polymerase chain reaction (m-RT-PCR) for simultaneous detection of five RNA viruses affecting stone fruit trees. Int. J. Virol., 5: 100-108.
CrossRefDirect Link - Yoon, J.Y., G.S. Choi, S.K. Choi, J.S. Hong and J.K. Choi et al., 2008. Molecular and biological diversities of Cucumber green mottle mosaic virus from cucurbitaceous crops in Korea. J. Phytopathol., 156: 408-412.
CrossRef