
P. Theerakulpisut
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
W. Gunnula
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Asian Journal of Crop Science
Year: 2012 | Volume: 4 | Issue: 4 | Page No.: 165-170







ABSTRACT
Salinity is one of the major factors limiting plant growth by imposing the osmotic, ion toxicity and oxidative stresses. The accumulation of osmoprotectants is a mechanism in which plants employ to protect tissue damages caused by salinity stress. The present study aimed to investigate the effect of two osmoprotectants (Sorbitol: Sor and Trehalose: Tre) on growth and physiology of rice (Oryza sativa L.) cvs. Khao Dawk Mali (KDML105; salt-sensitive) and Pokkali (PK: salt-tolerant) under NaCl stress. Under salt stress, KDML105 showed a significantly reduced growth and a large increase in the amount of hydrogen peroxide (H2O2), lipid peroxidation as indicated by Malondialdehyde (MDA) content and membrane Electrolyte Leakage (EL). The effects of salt were less severe in PK. Exogenously supplied Sor and Tre were able to enhance growth of salt-stressed KDML105 and alleviated the negative effects of salt by reducing H2O2 and MDA content. However, Sor and Tre did not have any beneficial effect on growth of PK plants. In some cases Sor worsened the salt-induced effects on lipid peroxidation and membrane damage. It can be concluded that exogenous Sor and Tre showed protective roles for salt-sensitive KDML105 but not salt-tolerant PK and the effects were more pronounced for Tre than Sor.
PDF Abstract XML References Citation
Received: December 23, 2011;
Accepted: April 02, 2012;
Published: May 12, 2012
How to cite this article
P. Theerakulpisut and W. Gunnula, 2012. Exogenous Sorbitol and Trehalose Mitigated Salt Stress Damage in Salt-sensitive but not Salt-tolerant Rice Seedlings. Asian Journal of Crop Science, 4: 165-170.
DOI: 10.3923/ajcs.2012.165.170
URL: https://scialert.net/abstract/?doi=ajcs.2012.165.170
DOI: 10.3923/ajcs.2012.165.170
URL: https://scialert.net/abstract/?doi=ajcs.2012.165.170
INTRODUCTION
Rice, one of the world’s most important staple crops, can be cultivated in most climatic conditions but is relatively intolerant to soil salinity. Plants grown in the medium containing high salt concentrations suffer from osmotic stress and high concentration of salts, Na+ can displace Ca2+ from the plasma membrane, resulting in a change in plasma membrane permeability that can be detected as leakage of ions from cells (Taiz and Zeiger, 2002; Nabipour et al., 2007; Amirjani, 2010). The simplest way to maintain favorable water potential gradient between the plants and the medium is by increasing cell’s solute concentration. Many plants produce and accumulate high level of compatible solutes, such as glycine betaine, proline, sorbitol (Sor), trehalose (Tre) and ectoine (Hare et al., 1998), to defend against osmotic stress. Sugar alcohols such as Sor, mannitol and inositol also play a role in tolerance to low temperature, drought and salt-stress (Williamson et al., 2002). Tre is a non-reducing disaccharide of glucose which plays an important physiological role as an abiotic stress protectant in a large number of organisms (Tao et al., 2008). Salt stress also induces oxidative stress due to over-accumulation of Reactive Oxygen Species (ROS). Lipid peroxidation is considered the most damaging process caused by ROS attack on polyunsaturated fatty acids in the membrane, leading to loss of membrane integrity and Malondialdehyde (MDA) amount is often used to monitor the level of lipid peroxidation under oxidative stresses. (El-baky et al., 2003; Li, 2009; Joseph and Jini, 2010, 2011).
Many studies have demonstrated that exogenously supplied osmoprotectants are able to improve plant tolerance under abiotic stress. In particular, Kadota et al. (2001) demonstrated in Japanese pear that applying Sor to growth media improved the shoot proliferation efficiency and fresh mass. Supplementing Sor in the growth media dramatically increased plantlets regeneration frequency from mature- and immature-derived calli of four rice cultivars (Geng et al., 2008). For Tre, Rasanen et al. (2004) found that exogenous Tre was advantageous for the Acacia senegal-Sinorhizobium symbiosis and protected Sinorhizobium strains from salt and osmotic stress. Pre-treatment of winter wheat with Tre was able to protect the proteins in the thylakoid membranes and the photosynthetic capacity under heat stress (Luo et al., 2010).
The present study was conducted to compare the effect of exogenous sugars (Sor and Tre) on growth, H2O2 content, MDA accumulation and membrane Electrolyte Leakage (EL) in seedlings of two rice cultivars under salt stress. Pokkali (PK) is a well-known salt-tolerant landrace from coastal India popularly used as donor of salt- tolerant traits in breeding programs. Khao Dawk Mali 105 (KDML105) is an economically important salt-sensitive Thai cultivar famous for its distinctive fragrance and high cooking quality.
MATERIALS AND METHODS
Two rice cvs. PK and KDML105 were used in this study. Seeds were sterilized with 70% ethyl alcohol and then incubated with 35% sodium hypochlorite for 1 h and washed extensively with distilled water three times. Seeds were then germinated in half strength Murashige and Skoog (½ MS) medium (Murashige and Skoog, 1962) for 4 days. The plants were then transferred to fresh ½ MS medium (C), ½ MS subjected to 170 mM NaCl (N) and NaCl combined with 5 mM (5 SN) and 10 mM Sor (10 SN) or 5 mM (5 TN) and 10 mM Tre (10 TN). The seedlings were cultured under controlled or NaCl-stressed media at 25±2°C with 16 h-light/8 h-dark cycles (40 μmol m-2 sec-1). Plants were analyzed for shoot length, dry weight (DW), H2O2 contents, MDA content and the EL percentage. H2O2 was measured according to the method described by Velikova et al. (2000). Determination of MDA content was preformed according to the methods described by Heath and Packer (1968). The percentage of EL was estimated according to Lui et al. (1985).
RESULTS
Under salt treatment, reduction of growth was observed in both rice cultivars. Exogenous application of Sor or Tre did not affect shoot length of KDML105 (Fig. 1a). However, supplementing Sor at 10 mM or Tre (5 and 10 mM) resulted in an increase in KDML105 DW (Fig. 1b). In addition, a significant reduction in H2O2 production and decreasing MDA content were observed in KDML105 supplied with Sor or Tre (Fig. 2a). The percentage reduction in H2O2 content was much higher for Tre than Sor while Sor showed more pronounced positive effect on the reduction of MDA content than Tre (Fig. 2b). Salt stress caused a significant increase in EL of KDML105 (Fig. 2c). Adding Sor or low concentration of Tre did not alleviate this salt-induced effect but higher concentration of Tre helped reduce membrane damage of KDML105 tissues.
For PK, exogenous Sor and low concentration of Tre (5 mM) caused a further reduction in plant shoot length (Fig. 1a) but 10 mM of Tre did not have any effect. Moreover, Sor (5 mM) further reduced PK DW (Fig. 1b).
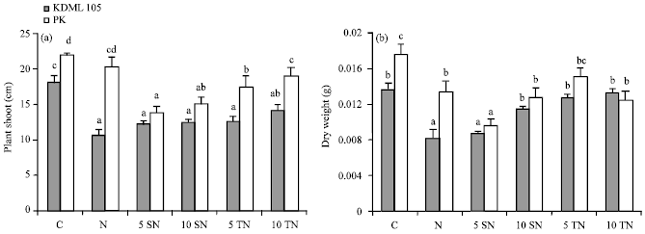 | |
| Fig. 1(a-b): | The effect of NaCl (N), NaCl combined with 5 mM (5 SN) and 10 mM (10 SN) Sor, NaCl combined with 5 mM (5 TN) and 10 mM (10 TN) Tre on (a) Shoot length and (b) Dry weight of rice seedlings |
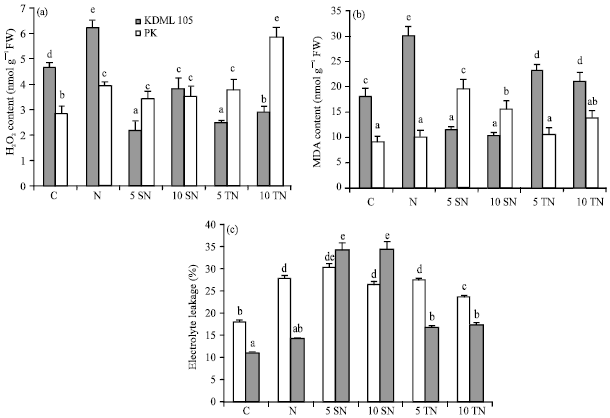 | |
| Fig. 2(a-c): | The effect of NaCl (N), NaCl combined with 5 mM (5 SN) and 10 mM (10 SN) Sor, NaCl combined with 5 mM (5 TN) and 10 mM (10 TN) Tre on (a) H2O2, (b) MDA and (c) Electrolyte leakage of rice seedlings |
Nevertheless, high concentration of Sor (10 mM) or Tre (5 and 10 mM) did not alter PK DW. Both concentrations of Sor and low level of Tre (5 mM) did not alter H2O2 content in PK plants but 10 mM Tre caused a great deal of enhancement in H2O2 production (Fig. 2a). PK plants fed with Sor showed a remarkable increase in MDA whilst 5 mM Tre did not affect MDA content but 10 mM Tre cause a small increase (Fig. 2b). Notably, adding Sor in combination with NaCl worsened the salt-induced effect on membrane damage in PK by increasing EL while Tre caused a slight insignificant increase in EL (Fig. 2c).
DISCUSSION
Exogenous Sor and Tre were more effective in alleviating salt-stress damage in the salt-sensitive rice seedlings than the salt-tolerant ones. Supplementing low concentration of Sor to PK even caused a further reduction in plant growth. Similar results showing the negative effects of Sor were observed in Guinean cashew; exogenously applied Sor showed the lowest percentage of shoots initiation and shoot length (Gemas and Bessa, 2006). Transgenic tobacco transformed with sorbitol-6-phosphate dehydrogenase (Stpd 1) gene from apple over-accumulated Sor and showed a slower growth, chlorophyll loss and necrotic lesions in young leaves. Transgenic lines producing higher concentrations of Sor also suffered from severe root growth inhibition. It was suggested that hyperaccumulation of Sor led to osmotic imbalance and possibly act as a signal affecting carbohydrate allocation and transport (Sheveleva et al., 1998). Conversely, high concentration of Sor improved growth of the salt-sensitive KDML105. These results corresponded with Jain et al. (2010) who reported that exogenous Sor increased DW in maize under osmotic stress. This suggested that the benefit of Sor varied depending on plant species, cultivars of the same species and the concentrations used. The positive effect of Tre on growth improvement was also observed in KDML105. The mitigating effects of Tre were also observed in maize pre-treated with Tre before exposure to NaCl (Zeid, 2009). In addition, under drought stress, foliar-applied Tre significantly increased maize biomass (Ali and Ashraf, 2011).
Salt stress usually leads to oxidative stress through the increase in ROS including H2O2 (Mudgal et al., 2010). The H2O2 content increased in both cultivars in response to salt stress and the level was much higher in the salt-sensitive cultivar corresponding to an earlier report (Theerakulpisut et al., 2005). Supplementing with both Sor and Tre caused marked reduction in H2O2 content in KDML105. A decrease in H2O2 accumulation has been shown in wheat treated with Tre. Luo et al. (2008) suggested that Tre played a direct role in eliminating H2O2 and O2-C. In general, salt stress caused a significant increase in MDA content and membrane damage in salt-sensitive than salt-tolerant rice. The level of salt stress in this study did not significantly increase the content of MDA and the EL values in PK. Surprisingly, adding Sor to PK plants exacerbated the salt stress effects and led to a dramatic increase in MDA content and percentage of EL. On the contrary, Sor and Tre showed a positive role to alleviate H2O2 production and membrane damage from ROS in KDML105. Nery et al. (2008) has shown that the presence of Tre on both side of lipid bilayer membrane indeed helps protect membrane damage from oxidative stress by minimizing damage caused to lipids and proteins.
CONCLUSION
In conclusion, Sor and Tre were effective in reversing the adverse effects of salt stress on growth and physiological parameters relating to ROS generation, lipid peroxidation and membrane damage especially for salt-sensitive rice. Interestingly, these osmoprotectants, in some cases, exacerbated growth and physiological aspects in salt-tolerant rice. These observations suggest that Sor and Tre application may be considered an interesting alternative approach with a practical potential for the enhancement of salinity tolerance in salt-sensitive rice and other crops.
ACKNOWLEDGMENT
This study was supported by KKU Research Grant to the Genomics and Proteomics Research Group for Improvement of Salt-tolerant Rice.
REFERENCES
- Ali, Q. and M. Ashraf, 2011. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defence mechanism. J. Agron. Crop Sci., 197: 258-271.
CrossRefDirect Link - Amirjani, M.R., 2010. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. Am. J. Plant Physiol., 5: 350-360.
CrossRefDirect Link - El-Baky, A., H. Hanaa, M.A. Amal and M.M. Hussein, 2003. Influence of salinity on lipid peroxidation, antioxidant enzymes and electrophoretic patterns of protein and isoenzymes in leaves of some onion cultivars. Asian J. Plant Sci., 2: 1220-1227.
CrossRefDirect Link - Gemas, V. and A. Bessa, 2006. Influence of various carbohydrates in shoot development in nodal culture of Guinean Anacardium occidentale genotypes. Plant Cell, Tissue Organ Cult., 85: 103-108.
CrossRef - Geng, P., H. La, H. Wang and E.J.C. Stevens, 2008. Effect of sorbitol concentration on regeneration of embryogenic calli in upland rice varieties (Oryza sativa L.). Plant Cell Tissue Organ Cult., 92: 303-313.
Direct Link - Hare, P.D., W.A. Cress and J. van Staden, 1998. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ., 21: 535-553.
CrossRefDirect Link - Jain, M., S. Tiwary and R. Gadre, 2010. Sorbitol-induced changes in various growth and biochemical parameters in maize. Plant Soil Environ., 56: 263-267.
Direct Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Joseph, B. and D. Jini, 2010. Insight into the role of antioxidant enzymes for salt tolerance in plants. Int. J. Bot., 6: 456-464.
CrossRefDirect Link - Joseph, B. and D. Jini, 2011. Development of salt stress-tolerant plants by gene manipulation of antioxidant enzymes. Asian J. Agric. Res., 5: 17-27.
CrossRefDirect Link - Li, Y., 2009. Effects of NaCl stress on antioxidative enzymes of glycine Soja sieb. Pak. J. Biol. Sci., 12: 510-513.
CrossRefPubMedDirect Link - Lui, H.X., Y.R. Zeng, D.F. Chan and J.Y. Guo, 1985. The effect of temperate on superoxide dismutase in various organelles of cucumber seedling cotyledon with different cold torerance. Acta. Phytophys. Sinica., 1: 48-57.
Direct Link - Kadota, M., K. Imizu and T. Hirano, 2001. Double-phase in vitro culture using sorbitol increases shoot proliferation and reduces hyperhydricity in Japanese pear. Sci. Horticult., 89: 207-215.
CrossRefDirect Link - Luo, Y., W.M. Li and W. Wang, 2008. Trehalose: Protector of antioxidant enzymes or reactive oxygen species scavenger under heat stress?. Environ. Exp. Bot., 63: 378-384.
CrossRef - Luo, Y., F. Li, G.P. Wang, X.H. Yang and W. Wang, 2010. Exogenously-supplied trehalose protects thylakoid membranes of winter wheat from heat-induced damage. Biol. Plant., 54: 495-501.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Mudgal, V., N. Madaan and A. Mudgal, 2010. Biochemical mechanisms of salt tolerance in plants: A review. Int. J. Bot., 6: 136-143.
CrossRefDirect Link - Nabipour, M., M. Meskarbashee and S. Farzad, 2007. Sodium and potassium accumulation in different parts of wheat under salinity levels. Asian J. Agric. Res., 1: 97-104.
CrossRefDirect Link - Rasanen, L.A., S. Saijets, K. Jokinen and K. Lindstrom, 2004. Evaluation of the roles of two compatible solutes, glycine betaine and trehalose, for the Acacia senegal-Sinorhizobium symbiosis exposed to drought stress. Plant Soil, 260: 237-251.
CrossRef - Sheveleva, E.V., S. Marquez, W. Chmara, A. Zegeer, R.G. Jensen and H.J. Bohnert, 1998. Sorbitol-6-phosphate dehydrogenase expression in transgenic tobacco: High amounts of sorbitol lead to necrotic lesions. Plant Physiol., 117: 831-839.
CrossRefDirect Link - Tao, D., Y. Mu, F.L. Fu and W.C. Li, 2008. Transformation of maize with trehalose synthase gene cloned from Saccharomyces cerevisiae. Biotechnology, 7: 258-265.
CrossRefDirect Link - Theerakulpisut, P., S. Bunnag and K. Kong-ngern. 2005. Genetic diversity, salinity tolerance and physiological responses to NaCl of SIX RICE (Oryza sativa L.) cultivars. Asian J. Plant Sci., 4: 562-573.
Direct Link - Williamson, J.D., D.B. Jennings, W.W. Guo, D.M. Pharr and M. Ehrenshaft, 2002. Sugar alcohols, salt stress and fungal resistance: Polyols: Multifunctional plant protection?. J. Am. Soc. Hort. Sci., 127: 467-473.
Direct Link - Zeid, I.M., 2009. Trehalose as osmoprotectant for maize under salinity-induced stress. Res. J. Agric. Biol. Sci., 5: 613-622.
Direct Link - Velikova, V., I. Yordanov and A. Edreva, 2000. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci., 151: 59-66.
CrossRefDirect Link