
Baby Joseph
Interdisciplinary Research Unit, Department of Biotechnology, Malankara Catholic College, Mariagiri, Kaliakkavilai-629153 Kanyakumari District, Tamil Nadu, India
LiveDNA: 91.1173
D. Jini
Interdisciplinary Research Unit, Department of Biotechnology, Malankara Catholic College, Mariagiri, Kaliakkavilai-629153 Kanyakumari District, Tamil Nadu, India
Asian Journal of Agricultural Research
Year: 2011 | Volume: 5 | Issue: 1 | Page No.: 17-27







ABSTRACT
Salinity toxicity is a worldwide agricultural and eco-environmental problem. Salt stress is one of the major abiotic stresses faced by plants, which adversely affect their productivity. Salt stress causes reduction of crop yield and alterations in plant metabolism, including a reduced water potential, ion imbalances and toxicity and sometimes severe salt stress may even threaten survival. Salinity also leads to oxidative stress in plants due to the production of Reactive Oxygen Species (ROS) such as the super oxide radical, hydrogen peroxide and hydroxyl radical. Reactive Oxygen Species (ROS) generated during metabolic processes damage cellular functions and consequently lead to disease, senescence and cell death. Plants have evolved an efficient defense system by which the ROS is scavenged by antioxidant enzymes. Attempts to reduce oxidative damages under the salt stress conditions have included the manipulation of ROS scavenging enzymes by gene transfer technology. It is important to increase the productivity under stressful environment by developing plants that have well adapted to environmental stress through manipulating antioxidant system. In this study, the rational approaches to develop stress-tolerant plants by gene manipulation of antioxidant enzymes will be introduced to provide solutions for the global food and environmental problems in the 21st century.
PDF Abstract XML References Citation
Received: June 06, 2010;
Accepted: July 15, 2010;
Published: October 11, 2010
How to cite this article
Baby Joseph and D. Jini, 2011. Development of Salt Stress-tolerant Plants by Gene Manipulation of Antioxidant Enzymes. Asian Journal of Agricultural Research, 5: 17-27.
DOI: 10.3923/ajar.2011.17.27
URL: https://scialert.net/abstract/?doi=ajar.2011.17.27
DOI: 10.3923/ajar.2011.17.27
URL: https://scialert.net/abstract/?doi=ajar.2011.17.27
INTRODUCTION
Many arid and semi-arid regions in the world contain soils and water resources that are too saline for most of the common economic crops, which affect plants through osmotic effects, ion specific effects and oxidative stress (Munns, 2002; Pitman and Lauchli, 2002). High concentrations of salt in soil are causing large decreases in yields for a wide variety of crops all over the world (Sekmen et al., 2007). Currently, 20% of the world's cultivated land is affected by salinity, which results in the loss of 50% of agricultural yield (Zhu, 2001; Bartels and Sunkar, 2005). Many crop plants such as barley, maize and rice, are often subject to salinity stress (Sairam and Tyagi, 2004).
Abiotic pressures like salt stress and chemical insultance can impose limitations on crop productivity and also limit land available for farming, often in regions that can ill afford such constraints, thus highlighting a greater need for understanding how plants respond to adverse conditions with the hope of improving tolerance of plants to environmental stress (Joseph et al., 2010). The effects of various environmental stresses in plants are known to be mediated, at leas in part, by an enhanced generation of Reactive Oxygen Species (ROS) including. O2¯, H2O2 and OH- (Hernandez et al., 2000; Benevides et al., 2000). These ROS are highly reactive and can alter normal cellular metabolism through oxidative damage to membranes, proteins and nucleic acids; they also cause lipid peroxidation, protein denaturation and DNA mutation (Imlay, 2003). To prevent damage to cellular components by ROS, plants have developed a complex antioxidant system. The primary components of this system include carotenoids, ascorbate, glutathione and tocopherols, in addition to enzymes such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPX), peroxidases and the enzymes involved in ascorbate-glutathione cycle (Foyer and Halliwell, 1976), such as ascorbate peroxidase (APX) and Glutathione Reductase (GR). Many components of this antioxidant defense system can be found in various subcellular compartments (Hernandez et al., 2000).
The scavenging of ROS by increased activation of antioxidant enzymes can improve salt tolerance (Alscher et al., 2002). A relationship between salt tolerance and increased activation of antioxidant enzymes has been demonstrated in Plantago (Sekmen et al., 2007), pea (Hernandez et al., 2000), Arabidopsis, rice (Dionisio-Sese and Tobita, 1998), tomato, soybean and maize (Azevedo-Neto et al., 2006). However, this antioxidative mechanism seems to be not enough to protect plants from the elevated environmental stresses such as salinity. To maintain the productivity of plants under the salt stress condition, it is important to fortify the antioxidative mechanism of the chloroplasts by manipulating the antioxidant enzymes and small antioxidant molecules in the chloroplast. Transgenic plants over expressing single transgene of SOD, APX and GR separately in chloroplast or other compartment of plant cell were generated and displayed increased tolerance against the oxidative stress. It seems that multiple expressions of tolerance genes in target organelle and their efficient regulation of expressions using a stress-inducible promoter are required for the ideal transgenic plants with enhanced tolerance to environmental stresses.
Oxidative damage by salt stress: Oxygen is essential for the existence of aerobic life, but toxic Reactive Oxygen Species (ROS), which include the superoxide anion radical (.O2-), hydroxyl radical (OH-) and hydrogen peroxide (H2O2), are generated in all aerobic cells during metabolic processes (Foyer et al., 1994; Asada, 1999). Injury caused by these ROS is known as oxidative stress, which is one of the major damaging factors to plants exposed to environmental stress.
Production of ROS is increased under saline conditions (Hasegawa et al., 2000) and ROS-mediated membrane damage has been demonstrated to be a major cause of the cellular toxicity by salinity in rice, tomato and citrus (Mittova et al., 2004). The effects of various environmental stresses in plants are known to be mediated, at least in part, by an enhanced generation of Reactive Oxygen Species (ROS) including O2¯, H2O2 and OH¯ (Hernandez et al., 2000; Benevides et al., 2000). These ROS are highly reactive and can alter normal cellular metabolism through oxidative damage to membranes, proteins and nucleic acids; they also cause lipid peroxidation, protein denaturation and DNA mutation (Imlay, 2003).
Salt stress induces cellular accumulation of ROS which can damage membrane lipids, proteins and nucleic acids (Hernandez et al., 1993, 1999; Mansour et al., 2005; Ben-Amor et al., 2007; Eyidogan and Oz, 2007). Much of the injury to plants exposed to stress is connected with oxidative damage at the cellular level (Foyer and Noctor, 2003). During oxidative stress, the excess production of ROS causes membrane damage that eventually leads to cell death. For protection against ROS, plants contain antioxidant enzymes as well as a wide array of non-enzymatic antioxidants (Blokhina et al., 2003).
Regulation of Reactive Oxygen Species (ROS) at normal environment: Under normal growth conditions, low amounts of ROS such as superoxide radical (.O2¯), hydrogen peroxide (H2 O2), hydroxyl radical (.OH¯) and singlet oxygen (1O2) are metabolic byproducts of plant cells (Cai-Hong et al., 2005). Plants have developed the scavenging mechanism of ROS categorized as enzymatic and non-enzymatic (Reddy et al., 2004; Demiral and Turkan, 2005). When ROS increases, chain reactions start, in which superoxide dismutase (SOD) catalyzes the dismutation of. O2¯ radicals to molecular O2 and H2O2 (Meloni et al., 2003). The H2O2 is then detoxified in the ascorbate-glutathione cycle (Asada, 1999; Mittler, 2002), which involves the oxidation and re-reduction of ascorbate and glutathione through the ascorbate peroxidase (APX) and Glutathione Reductase (GR) action (Noctor and Foyer, 1998). If there is a serious imbalance in any cell compartment between the production of Reactive Oxygen Species (ROS) and antioxidant defense, oxidative stress and damage occurs (Mittler, 2002).
Role of antioxidant enzymes for salt tolerance in plants: To minimize the effects of oxidative stress, plant cells have evolved a complex antioxidant system, which is composed of low-molecular mass antioxidants (glutathione, ascorbate and carotenoids) as well as ROS-scavenging enzymes, such as: superoxide dismutase (SOD), calatase (CAT), ascorbate peroxidase (APX), guaiacol peroxidase (GPX) and Glutathione Reductase (GR) (Apel and Hirt, 2004). Moreover, plants with the high activity of peroxidase and active proline accumulation could acclimate to salts stress (100 mM NaCl, 24 h) independently of SOD activity (Kartashov et al., 2008). Several studies have pointed out that salt-tolerant species increased their antioxidant enzyme activities and antioxidant contents in response to salt treatment, whereas salt-sensitive species failed to do so (Shalata et al., 2001; Demiral and Turkan, 2005). The scavenging of ROS by increased activation of antioxidant enzymes can improve salt tolerance (Alscher et al., 2002). A relationship between salt tolerance and increased activation of antioxidant enzymes has been demonstrated in Plantago (Sekmen et al., 2007), pea (Hernandez et al., 2000), tomato, maize (Azevedo-Neto et al., 2006), sorghum (Costa et al., 2005; Heidari, 2009), soybean (Cicek and Cakyrlar, 2008) and mulberry (Harinasut et al., 2003). The antioxidant enzymes present in different organelles of the plant cell is shown in the Fig. 1.
The generation of ROS and increased the activity of many antioxidant enzymes during salt stress have been reported in cotton (Desingh and Kanagaraj, 2007), mulberry (Sudhakar et al., 2001; Harinasut et al., 2003), wheat (Sairam et al., 2002), tomato (Mittova et al., 2002), rice (Vaidyanathan et al., 2003), sugar beet (Bor et al., 2003) and maize (Azevedo-Neto et al., 2006). In general, the activities of antioxidant enzymes were increased in the root and shoot under saline stress. But the increase was more significant and consistent in the root (Kim et al., 2005).
The quantitative and qualitative aspects of changes are often related to the levels of resistance to salinity. Meratan et al. (2008) showed a remarkable increase in Peroxidase and Polyphenol oxidase between 50 to 200 mM NaCl in three Acanthophyllum species. In tomato and citrus, salt-tolerance is attributed to the increased activities of SOD, APX and CAT (Gueta-Dahan et al., 1997; Mittova et al., 2002). In rice, the salt-tolerant varieties have higher SOD activity and lower lipid peroxidation than the salt-sensitive varieties (Dionisio-Sese and Tobita, 1998). It was suggested that the ratio between superoxide dismutase and H2O2-scavenging enzyme activities could be used as a working hypothesis for a biochemical marker for salt tolerance in sorghum (Costa et al., 2005).
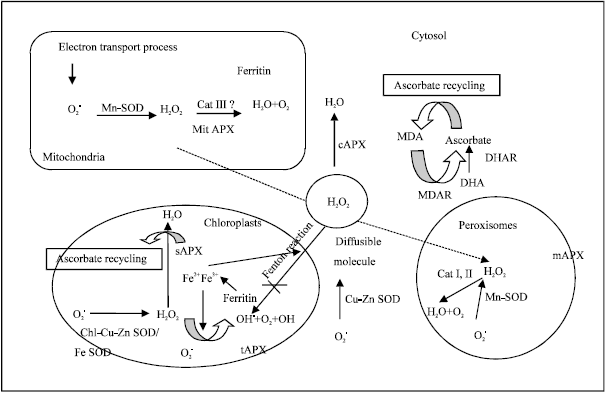 | |
| Fig. 1: | Antioxidant network in different organelles of the plant cell (Jithesh et al., 2006) |
Further supporting evidence on the involvement of antioxidant enzymes in salt tolerance has been provided by transgenic plants with a reduced or an increased expression of antioxidant enzymes (Willekens et al., 1997). Increased protection to salt stress has been demonstrated by the over expression of cytosolic APX (Torsethaugen et al., 1997).
Transgenic plant for salt tolerance
Expression of superoxide dismutase in salt tolerant transgenic plants: Superoxide dismutases catalyze the breakdown of superoxide radicals and provide the first line of defense against oxygen toxicity. They are essential for aerobic life and are involved in stress resistance and longevity (Carlioz and Touati, 1986; Orr and Sohal, 1994). Three groups of superoxide dismutases (SODs) can be distinguished on the basis of the metal cofactor at the active site: Cu/Zn SOD, Mn-SOD and Fe-SOD (Fridivich, 1989). Mn-SOD and Fe-SOD appear to be closely related both in structural and in evolutionary aspects, but have no resemblance to Cu/ZnSOD (Bowler et al., 1994). Cu/Zn SODs occur in the cytoplasm and chloroplasts; Mn-SODs are usually found in the mitochondrial matrix and in peroxisomes; and Fe-SODs are present within the chloroplasts of some plants (Raychaudhuri and Deng, 2000).
Zhao et al. (2009) introduced the transcription factor YAP1, originally from yeast (Saccharomyces cerevisiae), into Arabidopsis thaliana (ecotype Columbia). When treated with various NaCl concentrations, transgenic plants showed increased activities of antioxidant enzymes catalase, superoxide dismutase, ascorbate peroxidase, peroxidase, glutathione S-transferase and glutathione reductase compared with the wild-type Arabidopsis. This demonstrated that an active oxygen scavenging system was enhanced to protect plants from salt stress by equilibrating ROS metabolism. Transgenic Arabidopsis maintained higher photosynthesis levels and lower amounts of H2O2, suggesting that ROS production was reduced (Zhao et al., 2009). Wang et al. (2010) introduced a MnSOD gene (TaMnSOD) from Tamarix androssowii, under the control of the CaMV35S promoter, was introduced into poplar (Populus davidianaxP. bolleana). They showed that SOD activity was enhanced in transgenic plants and the MDA content were significantly decreased when exposed to NaCl stress (Wang et al., 2010).
Expression of catalase in salt tolerant transgenic plants: Catalases (CAT) are haem-containing tetrameric enzymes involved in the removal of H2O2 (Guan and Scandalios, 1996). Plant catalases are involved in photorespiratory functions (Canvin, 1990) and scavenging of H2O2 during β-oxidation of fatty acids in germinating seeds (Willekens et al., 1995) and also during salt stress and other abiotic stress conditions (Willekens et al., 1997). Catalase enzymes that can remove the Reactive Oxygen Species (ROS) produced by the salt stress play an important role in tolerance. Thus quenching of H2O2 was an important factor for salt tolerance as observed in cyanobacteria (Kaku et al., 2000). Nagamiya et al. (2007) were able to produce salt tolerant japonica rice (at 100 mM salt concentration) by over expressing the catalase gene katE. Prodhan et al. (2008) introduced katE, a catalase gene of Escherichia coli, into the indica rice cultivar Kasalath. Transgenic rice plants at a very young stage (three-four days) were able to grow up to 15 days in 100 mM NaCl solution and seven days in 250 mM NaCl solution whereas, control plants died within five days in 100 mM and seven days in 50 mM NaCl. Moriwaki et al. (2008) integrated the katE gene into BR5 rice plants using an Agrobacterium tumefaciens-mediated method. The introduced katE gene was actively expressed in the transgenic BR5 rice plants and catalase activity in transgenic rice was approximately 150% higher than in non transgenic plants. Under NaCl stress conditions, the transgenic rice plants exhibited high tolerance compared with non transgenic rice plants (Moriwaki et al., 2008).
Expression of glutathione s-transferase: Glutathione S-transferases (GSTs) are a family of multifunctional enzymes that play important roles in oxidative stress resistance. The Key Laboratory of Plant Stress, P. R. China, obtained a GST from Suaeda salsa cDNA library (accession number BE859255) and showed that it played an important role in salt stress resistance (Wang et al., 2002; Qi et al., 2004). Zhao and Zhang (2006) proved that the expression of Suaeda salsa GST gene in transgenic rice (Oryza sativa L. cv) could confer resistance to salt stress. Overexpression of a tobacco glutathione S-transferase with glutathione peroxidase activity (GST/GPX) in transgenic tobacco (Nicotiana tabacum L.) enhanced seedling growth under salt stress conditions (Roxas et al., 2000). Qi et al. (2010) introduced the Suaeda salsa glutathione S-transferase gene (GST) into Arabidopsis under the control of the cauliflower mosaic virus 35S promoter. The expression of the GST gene promoted a higher level of salt tolerance in vivo in transgenic Arabidopsis plants. Southern and northern blot analyses confirmed that GST was transferred into the arabidopsis genome and the GST and GPX activities in transgenic plants (GT) were much higher than in wild-type plants (WT) (Qi et al., 2010).
Expression of other enzymes in transgenic plants: The transgenic plants overexpressing GR have higher ascorbate contents and improved tolerance to oxidative stress (Foyer et al., 1991, 1995; Aono et al., 1993). On the other hands, reduced GR activity resulted in increased stress sensitivity (Aono et al., 1993). Ascorbate-deficient mutant of Arabidopsis is sensitive to oxidative stress such as UV and pollutants (Conklin et al., 1997).
Ascorbate acts as an important antioxidant in both enzymatic and non-enzymatic (reacting directly with hydroxyl radicals, superoxide and singlet oxygen) reactions in plant cells. Some of the monodehydro ascorbate (MDHA), oxidized form of ascorbate, is re-reduced by monodehydro ascorbate reductase (MDHAR) using NAD(P)H, but the remainder undergoes spontaneous dismutation to AsA (reduced ascorbate) and dehydro ascorbate (DHA, oxidized ascorbate). DHAR (EC 1.8.5.1) catalyzes the re-reduction of DHA to AsA with simultaneous oxidation of GSH to GSSG. Thus, DHAR, as well as MDHAR, is critical for protection of cellular components against oxidative injury (Asada, 1999). Also, DHAR activity is enhanced in response to various environmental stresses (Urano et al., 2000). The DHAR-overexpressing tobacco plants have been recently developed in our laboratory and have elevated level of tolerance to oxidative stress derived from various sources (Kwon et al., 2000).
Strategies to improve salt stress tolerance: Recent advances in plant genome mapping and molecular biology techniques offer a new opportunity for understanding the genetics of salt stress-resistance genes and their contribution to plant performance under salt stress. These biotechnological advances will provide new tools for breeding in salt stress environment. Molecular genetic maps have been developed for major crop plants, including rice, wheat, maize, barley, sorghum and potato, which make it possible for scientists to tag desirable traits using known DNA landmarks. Molecular genetic markers allow breeders to track genetic loci controlling stress resistance without having to measure the phenotype, thus reducing the need for extensive field-testing over time and space. Moreover, gene pyramiding or introgression can be done more precisely using molecular tags. Together, molecular genetic markers offer a new strategy known as marker assisted selection. Another molecular strategy which depends on gene cloning and plant transformation technology is genetic engineering of selected genes into elite breeding lines. What makes a particular goal attainable or unattainable in genetic engineering experiments is the availability of the following three inputs: (1) the gene of interest, (2) an effective technique for transferring the desired gene from one species to another and (3) promoter sequences for regulated expression of that gene. Amongst these, the first is considered a rate-limiting factor. Arrays of salt stress-induced genes have been isolated. Salt stress-responsive genes can be analyzed following targeted or non-targeted strategy. The targeted approach relies upon the availability of relevant biochemical information (i.e., in terms of defined enzyme, protein, a biochemical reaction or a physiological phenomenon). The non-targeted strategy to obtain a desired gene is indirect. This strategy, for instance, includes differential hybridization and shotgun cloning. The list of genes, whose transcription is upregulated in response to stress, is rapidly increasing. Understanding of the mechanisms which regulate gene expression and the ability to transfer genes from other organisms into plants will expand the ways in which plants can be utilized. To exploit the full potential of these approaches, it is essential that the knowledge is applied to agriculturally and ecologically important plant species.
CONCLUSION AND FUTURE PROSPECTS
Salinity in the soil and irrigation water is an environmental problem and a major constraint for crop production. Salt stress (NaCl) has both osmotic (cell dehydration) and toxic (ion accumulation) effects on plants. Although a wide range of genetic adaptations to saline conditions has been observed and a number of significant physiological responses have been associated with tolerance, underlying mechanisms of salt tolerance in plants are still poorly understood.
The ROS, especially hydrogen peroxide, have been proved as a central component of plant adaptation to biotic and abiotic stresses (50) (Mittler, 2002). Under the stress conditions, ROS may play in damaging the cellular components or signaling the activation of defense responses. To allow for these different roles, cellular levels of ROS must be tightly controlled. In first, precise understanding the roles of each ROS scavenging enzyme and small molecular antioxidants in stress adaptation and accurate characterization of the complex stress tolerance phenotypes is necessary to develop stress tolerant plants.
Cloned plant genes and transgenic plants have become a standard tool in plant-stress biology. These technologies have mainly been applied to model systems and have greatly enlarged the knowledge of mechanisms of tolerance. The various abiotic stresses cause changes in plant processes at all levels of organization (morphological, physiological, biochemical and molecular). In recent years, attention has focused on alterations in gene expression. The list of genes whose transcription is upregulated in response to stress is rapidly increasing. Functions for some of these polypeptides are close to being identified and their likely role in stress physiology is being determined. The understanding of mechanisms that regulate gene expression and the ability to transfer genes from other organisms into plants will expand the ways in which plants can be utilized. The exploitation of cloned genes to alter the function of gene products in transgenic plants provides novel opportunities to assess their biological role in a salt stress response. We are trying to develop ideal transgenic plants with enhanced tolerance to salt stress by expression of multiple antioxidant enzymes in target organelle of a plant cell under the control of salt stress-inducible promoter.
ACKNOWLEDGMENTS
The authors gratefully acknowledged to our Malankara Catholic College Correspondent Fr. Prem Kumar (MSW) given encouragement and support for preparation of this manuscript. We wish to express our sincere thanks to Dr. Sujatha (ICBM), Mr. K. Suresh and the research scholars in the Interdisciplianary Research centre for updated article collection and all the efficient supports of this review preparation.
REFERENCES
- Aono, M., A. Kubo, H. Saji, K. Tanaka and N. Kondo, 1993. Enhanced tolerance to photooxidative stress of transgenic Nicotiana tabacum with high chloroplastic glutathione reductase activity. Plant Cell Physiol., 34: 129-135.
Direct Link - Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Asada, K., 1999. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50: 601-639.
CrossRefPubMedDirect Link - De Azevedo-Neto, A.D., J.T. Pitsco, J. Eneas-Filho, C.E.B. de Abreu and E. Gomes-Filho, 2006. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot., 56: 87-94.
CrossRefDirect Link - Bartels, D. and R. Sunkar, 2005. Drought and salt tolerance in plants. Crit. Rev. Plant Sci., 24: 23-58.
CrossRefDirect Link - Benevides, M.P., P.L. Marconi, S.M. Gullego, M.E. Comba and M.L. Tomaro, 2000. Relationship between antioxidant defense system and salt tolerance in Solanum tuberosum. Aust. J. Plant Physiol., 27: 273-278.
CrossRef - Blokhina, O., E. Virolainen and K.V. Fagerstedt, 2003. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot., 91: 179-194.
CrossRefPubMedDirect Link - Bor, M., F. Ozdemir and I. Turkan, 2003. The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Sci., 164: 77-84.
CrossRefDirect Link - Bowler, C., W. van Camp, M. van Montagu, D. Inze and K.Asada, 1994. Superoxide dismutase in plants. Crit. Rev. Plant Sci., 13: 199-218.
CrossRefDirect Link - Cai-Hong, P., Z. Su-Jun, G. Zhi-Zhong and W.W. Bao-Shan, 2005. NaCl treatment markedly enhances H2O2-scavenging system in leaves of halophyte Suaeda salsa. Physiol. Plant., 125: 490-499.
CrossRef - Carlioz, A. and D. Touati, 1986. Isolation of superoxide dismutase mutants in Escherichia coli: Is superoxide dismutase necessary for aerobic life. EMBO J., 5: 623-630.
Direct Link - Cicek, N. and H. Cakırlar, 2008. Changes in some antioxidant enzyme activities in six soybean cultivars in response to long-term salinity at two different temperatures. Gen. Appl. Plant Physiol., 34: 267-280.
Direct Link - Conklin, P.L., J.E. Pallanca, R.L. Last and N. Smirnoff, 1997. L-Ascorbic acid metabolism in the ascorbate-deficient Arabidopsis mutant vtc1. Plant Physiol., 115: 1277-1285.
Direct Link - Costa, P.H.A., A. Neto, M. Bezerra, J. Prisco and E. Filho, 2005. Antioxidant-enzymatic system of two sorghum genotypes differing in salt tolerance. Braz. J. Plant Physiol., 17: 353-361.
Direct Link - Demiral, T. and I. Turkan, 2005. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ. Exp. Bot., 53: 247-257.
CrossRefDirect Link - Desingh, R. and G. Kanagaraj, 2007. Influence of salinity stress on photosynthesis and antioxidative systems in two cotton varieties. Gen. Applied Plant Physiol., 33: 221-234.
Direct Link - Dionisio-Sese, M.L. and S. Tobita, 1998. Antioxidant responses of rice seedlings to salinity stress. Plant Sci., 135: 1-9.
CrossRefDirect Link - Eyidogan, F. and M.T. Oz, 2007. Effect of salinity on antioxidant responses of chickpea seedlings. Acta Physiol. Plant, 29: 485-493.
CrossRef - Foyer, C.H., N. Souriau, S. Perret, M. Lelandais, K.J. Kunert, C. Pruvost and L. Jouanin, 1995. Overexpression of glutathione reductase but not glutathione synthetase leads to increases in antioxidant capacity and resistance to photoinhibition in poplar trees. Plant Physiol., 109: 1047-1057.
Direct Link - Foyer, C.H. and B. Halliwell, 1976. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta, 133: 21-25.
CrossRefPubMedDirect Link - Foyer, C.H. and G. Noctor, 2003. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant., 119: 355-364.
CrossRefDirect Link - Foyer, C.H., P. Descourvieres and K.J. Kunert, 1994. Protection against oxygen radicals: An important defense mechanism studied in transgenic plants. Plant Cell Environ., 17: 507-523.
CrossRef - Foyer, C.H., M. Lelandais, C. Galap and K.J. Kunert, 1991. Effects of elevated cytosolic glutathione reductase activity on the cellular glutathione pool and photosynthesis in leaves under normal and stress conditions. Plant Physiol., 97: 863-872.
Direct Link - Fridivich, I., 1989. Superoxide dismutases: An adaptation to a paramagnetic gas. J. Biol. Chem., 264: 7761-7764.
PubMedDirect Link - Guan, L.Q. and J.G. Scandalios, 1996. Molecular evolution of maize catalases and their relationship to other eukaryotic and prokaryotic catalases. J. M. Evol., 42: 570-579.
CrossRef - Gueta-Dahan, Y., Z. Yaniv, B.A. Zilinskas and G. Ben-Hayyim, 1997. Salt and oxidative stress: Similar and specific responses and their relation to salt tolerance in citrus. Planta, 204: 460-469.
CrossRef - Harinasut, P., D. Poonsopa, K. Roengmongkol and R. Charoensataporn, 2003. Salinity effects on antioxidant enzymes in mulberry cultivar. Sci. Asia, 29: 109-113.
Direct Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Heidari, M., 2009. Antioxidant activity and osmolyte concentration of sorghum (Sorghum bicolor) and wheat (Triticum aestivum) genotypes under salinity stress. Asian J. Plant Sci., 8: 240-244.
CrossRefDirect Link - Hernandez, J.A., A. Campillo, A. Jimenez, J.J. Alarcon and F. Sevilla, 1999. Response of antioxidant systems and leaf water relations to NaCl stress in pea plants. New Phytol., 141: 241-251.
CrossRefDirect Link - Hernandez, J.A., F.J. Corpas, M. Gomez, L.A. Rio and F. Sevilla, 1993. Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria. Physiol. Plant., 89: 103-110.
CrossRefDirect Link - Hernandez, J.A., A. Jimenez, P. Mullineaux and F. Sevilia, 2000. Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ., 23: 853-862.
CrossRefDirect Link - Jithesh, M.N., S.R. Prashanth, K.R. Sivaprakash and A.K. Parida, 2006. Antioxidative response mechanisms in halophytes: Their role in stress defence. J. Genet., 85: 237-254.
CrossRefDirect Link - Kartashov, A.V., N.L. Radyukina, Y.V. Ivanov, P.P. Pashkovskii, N.I. Shevyakova and V.V. Kuznetsov, 2008. Role of antioxidant systems in wild plant adaptation to salt stress. Russ. J. Plant Physiol., 55: 463-468.
CrossRef - Kim, S., J. Lim, M. Park, Y. Kim, T. Park, Y. Seo and S. Yun, 2005. Enhanced antioxidant enzymes are associated with reduced hydrogen peroxide in barley roots under saline stress. J. Biochem. Mol. Biol., 38: 218-224.
Direct Link - Mansour, M.M.F., K.H.A. Salama, F.Z.M. Ali and A.F. Abou Hadid, 2005. Cell and plant responses to NaCl in Zea mays L. cultivars differing in salt tolerance. Gen. Applied Plant Physiol., 31: 29-41.
Direct Link - Meloni, D.A., M.A. Oliva, C.A. Martinez and J. Cambraia, 2003. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot., 49: 69-76.
CrossRefDirect Link - Meratan, A.A., S.M. Ghaffari and V. Niknam, 2008. Effects of salinity on growth, proteins and antioxidant enzymes in three Acanthophyllum species of different ploidy levels. JSUT, 33: 1-8.
Direct Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Mittova, V., M. Guy, M. Tal and M. Volokita, 2004. Salinity upregulates the antioxidative system in root mitochondria and peroxisomes of the wild salt-tolerant tomato species Lycopersicon pennellii. J. Exp. Bot., 55: 1105-1113.
Direct Link - Mittova, V., M. Tal, M. Volokita and M. Guy, 2002. Salt stress induces up-regulation of an efficient chloroplasts antioxidant system in the salt-tolerant wild tomato species Lycopersicon pennellii but not in the cultivated species. Plant Physiol., 115: 393-400.
Direct Link - Moriwaki, T., Y. Yamamoto, T. Aida, T. Funahashi and T. Shishido and M. Asada et al., 2008. Overexpression of the Escherichia coli catalase gene, katE, enhances tolerance to salinity stress in the transgenic indica rice cultivar, BR5. Plant Biotechnol. Rep., 2: 41-46.
CrossRef - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Mol. Biol., 49: 249-279.
CrossRefPubMedDirect Link - Orr, W.C. and R.S. Sohal, 1994. Extension of life-span by overexpression of superoxide dismutase and catalase in Drosophila melanogaster. Science, 263: 1128-1130.
CrossRefDirect Link - Qi, Y.C., W.Q. Liu, L.Y. Qiu, S.M. Zhang, L. Ma and H. Zhang, 2010. Overexpression of glutathione S-transfer gene increases salt tolerance of arabidopsis. Russian J. Plant Physiol., 57: 233-240.
CrossRef - Qi, Y.C., S.M. Zhang, L.P. Wang, M.D. Wang and H. Zhang, 2004. Overexpression of GST gene accelerates the growth of transgenic Arabidopsis under salt stress. J. Plant Physiol. Mol. Biol., 30: 517-522.
PubMed - Raychaudhuri, S.S. and X.W. Deng, 2000. The role of superoxide dismutase in combating oxidative stress in higher plants. Bot. Rev., 66: 89-98.
CrossRef - Reddy, A.R., K.V. Chaitanya and M. Vivekanandan, 2004. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol., 161: 1189-1202.
CrossRefPubMedDirect Link - Roxas, V.P., S.A. Lodhi, D.K. Garrett, J.R. Mahan and R.D. Allen, 2000. Stress tolerance in transgenic tobacco seedlings that overexpress glutathione S-transferase/glutathione peroxidase. Plant Cell Physiol., 41: 1229-1234.
Direct Link - Sairam, R.K. and A. Tyagi, 2004. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci., 86: 407-421.
Direct Link - Sairam, R.K., K.V. Roa and G.C. Srivastava, 2002. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci., 163: 1037-1046.
CrossRefDirect Link - Sekmen, A.H., I. Turkana, and S. Takiob, 2007. Differential responses of antioxidative enzymes and lipid peroxidation to salt stress in salt-tolerant Plantago maritime and salt-sensitive Plantago media. Physiol. Plant, 131: 399-411.
CrossRef - Shalata, A., V. Mittova, M. Volokita, M. Guy and M. Tal, 2001. Response of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii to salt-dependent oxidative stress: The root antioxidative system. Physiol. Plant., 112: 487-494.
CrossRefPubMedDirect Link - Sudhakar, C., A. Lakshmi and S. Giridarakumar, 2001. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Sci., 161: 613-619.
CrossRefDirect Link - Vaidyanathan, H., P. Sivakumar, R. Chakrabarty and G. Thomas, 2003. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)-differential response in salt-tolerant and sensitive varieties. Plant Sci., 165: 1411-1418.
CrossRefDirect Link - Wang, Y.C., G.Z. Qu, H.Y. Li, Y.J. Wu, C. Wang, G.F. Liu, and C.P. Yang, 2010. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Bio. Rep., 37: 1119-1124.
CrossRef - Zhao, F. and H. Zhang, 2006. Expression of Suaeda salsa glutathione S-transferase in transgenic rice resulted in a different level of abiotic stress resistance. J. Agric. Sci., 144: 547-554.
CrossRef - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Alscher, R.G., J.L. Donahue and C.L. Cramer, 1997. Reactive oxygen species and antioxidants: Relationship in green cells. Physiol. Plant., 100: 224-233.
CrossRefDirect Link