
Haitham K.A. EL-Amin
Sudan Academy of Sciences, Ministry of Science and Technology, P.O. Box 86, Khartoum, Sudan
LiveDNA: 249.8062
Nada B. Hamza
Department of Molecular Biology, Commission for Biotechnology and Genetic Engineering, National Centre for Research, Khartoum, P.O. Box 2404, Sudan
LiveDNA: 249.11885
American Journal of Biochemistry and Molecular Biology
Year: 2013 | Volume: 3 | Issue: 1 | Page No.: 127-134







ABSTRACT
Sorghum (Sorghum bicolor L.) is believed to be domesticated for the first time within the geographical range of Sudan, so, studying the genetic variation of sorghum genotypes collections attracts special interest. In this study, 17 sorghum accessions with important agronomic traits, representing four states in Sudan were assayed for polymorphism using Random Amplified Polymorphic DNA technique (RAPD). Ten primers out of 40 tested (A-1, B-20, C-20, D-18, OPE-04, UBC-101, UBC-103, UBC-127, UBC-155 and UBC-157) showed high polymorphism among the accessions. The results indicated 110 polymorphic bands out of 145 bands with percentage of polymorphic bands of 77.6%. Unweighted Pair Group Method with Arithmetic Mean (UPGMA) result showed two major clusters, with a clear trend of grouping of each region. A cluster contained all accessions from eastern Sudan (Red Sea). However, accessions from north Sudan (River Nile), western Sudan (North Kordofan) and south Sudan (Blue Nile) were diverse in another cluster. Based on the results of this study RAPD technique proved to be useful to study genetic variation among the Sudanese sorghum accessions.
PDF Abstract XML References Citation
Received: February 06, 2012;
Accepted: March 07, 2012;
Published: October 04, 2012
How to cite this article
Haitham K.A. EL-Amin and Nada B. Hamza, 2013. Phylogenetic Diversity of Sorghum bicolor (L.) Moench Accessions from Different Regions in Sudan. American Journal of Biochemistry and Molecular Biology, 3: 127-134.
DOI: 10.3923/ajbmb.2013.127.134
URL: https://scialert.net/abstract/?doi=ajbmb.2013.127.134
DOI: 10.3923/ajbmb.2013.127.134
URL: https://scialert.net/abstract/?doi=ajbmb.2013.127.134
INTRODUCTION
In Africa, especially in Sudan Sorghum bicolor is a large subsistence food, forge and ethanol production crop for more than 75% of the population (Dawelbeit et al., 2010). Sorghum has attracted the attention of breeders not only in Sudan but around the world for his rich source of proteins, vitamins, carbohydrates and economical important (Shiringan, 2009). Sorghum is a diploid (2n = 20) and inter fertile (Curtis, 1968). About 2512 loci exist in its genetic map (Bowers et al., 2003). Sorghum has small genome (730 Mb); therefore was considered as model for functional genomics of C4 grasses, it also set a good example to understand the biology of weeds (Paterson, 2008). The Morphological variation doesn’t express the genetic variation due to the genotype-environment interactions (Smith and Smith, 1992). Therefore, studying genetic variation is important (Bandyopadhyay, 2011). Wide spectrum of methods and techniques exist for the analysis of discrete morphological traits to molecular traits (Soller and Beckmann, 1983). Therefore, understanding the real genetic variation is crucial for breeding programs in order to help maintain genetic diversity (Pu et al., 2009).
Molecular markers proved to be powerful and reliable tools for studying the variation within crop genotypes compared with the biochemical markers (Gepts, 1993). Random Amplified Polymorphic DNA (RAPD) technique is one of the most used molecular methods, as it offers many advantages such as detect high polymorphism and relatively fast (Fahima et al., 1999), RAPD could also reveal within accessions variation more efficiently (Patra et al., 2011). The RAPD technique is based on the PCR amplification of discrete regions inside the genome with short oligonucleotide primers of arbitrary sequence (Thangaraj et al., 2011). A minimal amount of DNA is required and no DNA sequence information is needed before carrying the technique (Subudhi and Huang, 1999). Although of the robustness of the method, its reproducibility can vary from low to extremely high variability (Fang and Roose, 1997; Nagaoka and Ogihara, 1997). From its advantages is that it is simple and fast method for detecting high amount of DNA markers (Younis et al., 2007). Over 800 markers are mapped in sorghum, these maps are compared to those of rice (Paterson et al., 1995, 2004), maize (Bowers et al., 2003; Whitkus et al., 1992), sugarcane (Ming et al., 1998; Dufour et al., 1997), millet (Jessup et al., 2003), switch grass (Missaoui et al., 2005) and Bermuda grass (Bethel et al., 2006).
The main objectives of the study were to investigate the degree of polymorphism detected using 10 RAPD (Random Amplified Polymorphic DNA) markers in 17 sorghum accessions from four different regions in Sudan.
MATERIALS AND METHODS
The DNA of 17 sorghum accessions was extracted from fresh leaf tissues (Table 1) using modified CTAB method (Porebski et al., 1997). The modification was made in intention to improve the quantity and the quality of the DNA. In this method the fine powdered plant materials were immediately transferred into 15 mL Falcon tubes containing 5 mL of pre-warmed lysis solution. Tubes containing the samples were then incubated in a water bath at 60°C with gentle shaking for 30 min and left to cool at room temperature for 5 min. Isoamylalcohol and chloroform mixture (1:24) was added to each tube and the phases were mixed gently for 5 min at room temperature to make a homogenous mixture.
| Table 1: | Sorghum genotypes list contained accessions number, accessions name, Area location of collection and the region of collection |
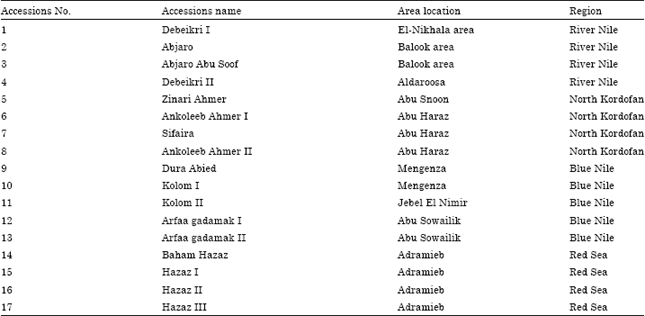 | |
The cell debris was removed by centrifugation at 4000 rpm for 15 min and the resulted clear aqueous phases (containing DNA) were transferred to new sterile tubes. The step of the chloroform: isoamyl alcohol extraction was repeated twice. The nucleic acids in the aqueous phase were precipitated by adding equal volume of deep cooled isopropanol. The contents were mixed gently and collected by centrifugation at 4000 rpm for 10 min. The formed DNA pellet was washed twice with 70% ethanol and the ethanol was discarded after spinning with flash centrifugation. The remained ethanol was removed by leaving the pellet to dry at room temperature. The pellet was dissolved in TE buffer (10 mM Tris, 1 mM EDTA, pH 8) and stored at -20°C for further use. The extracted DNA samples were observed under UV illumination after staining with ethidium bromide and agarose gel electrophoresis. The purity and the concentrations of the DNA were then spectrophotometrically assessed following Sambrook et al. (1989) method.
Forty RAPD primers (Operon technologies and university of British Columbia (UBC)) were used in the Polymerase Chain Reaction (PCR) in a final volume of 25 μL containing 1.0 μL DNA diluted, 0.5 μL Taq polymerase, 2.5 μL 10x buffer, 2.5 μL (2 mM μL-1) dNTPs, 1.5 μL (50 mM) MgCl2, 2.0 μL (10 pmol μL-1) RAPD primer and 15 μL dH2O. The amplifications were performed in a thermal cycler following the program: 94°C for 5 min, 40 cycles (1 min at 94°C, 1 min at 33°C and 1 min at 72°C) and final elongation of 7 min at 72°C.
Four Microliter of PCR product was mixed with 2 μL of loading dye and 1.5 μL of 1 kbp DNA ladder were electrophoresed using 2% agarose gel at 73 volts followed by staining with ethidium bromide then the separated fragments were visualized with an Ultraviolet (UV) transilluminator.
The number of polymorphic and monomorphic bands was determined for each primer. Genotype were scored (1) for present band and (0) for absent band and then entered into a data matrix. Percentage of polymorphism was calculated as the following equation: (polymorphic bands/total number of bands x100). The tree diagram was produced by clustering the similarity data with the unweighted pair group average method using statistical software package STATISTCA-SPSS following the method of El-Amin et al. (2011).
RESULTS
To isolate a good quality of DNA, the CTAB-based procedure optimized in the present study, yielded high quality DNA free of phenols which may inhibit the activity of Taq polymerase. Several primers were tested on the 17 accessions (Sorghum bicolor L.) and the results indicated that 10 primers out of the 40 tested showed a high polymorphic band percentage (75.9%). The 10 informative primers were selected and used to evaluate the degree of polymorphism and genetic relationships among the genotypes under study. Total of 145 amplified fragments were distinguished across the selected primers and the statistical analysis showed 110 polymorphic bands among the accessions with an average of 11 polymorphic bands per primer. The maximum number of bands were produced by the primer UBC-157 (21 bands) with 52.4% polymorphism respectively while the minimum numbers of fragments were produced by the primer C-20 (8 bands) with 62.5%, polymorphism, respectively. RAPD fragments pattern produced by the 10 primers is shown in Table 2.
The Tree Diagram reflects the accessions of the four regions which were divided into two clusters based on their genetic similarity. The first cluster contained 3 groups: River Nile group, North Kordofan group each grouped separately from each other and Blue Nile group. River Nile group have accessions Abjaro Abu Soof and Debeikri II from Aldaroosa area as sisters, Debeikri I from El-Nikhala area and Abjaro in same group but Abjaro more genetically related than Debeikri from El-Nikhala area.
| Table 2: | Polymorphic bands percentages of 10 RAPD primers used to study the 17 sorghum accessions |
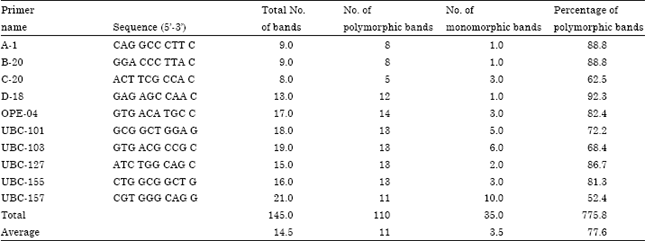 | |
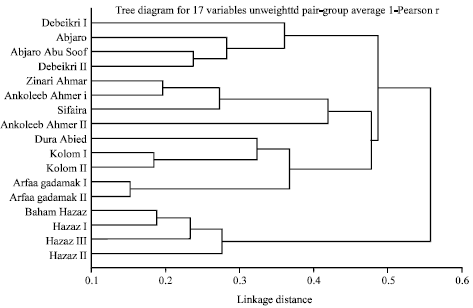 |
| Fig. 1: | Tree diagram constructed for 17 sorghum accessions based on their genetic distances using 10 RAPD primers |
North Kordofan group contained Zinari Ahmer and Ankoleeb Ahmer I as sisters, within the same group Sifaira closer to the sisters than Ankoleeb Ahmer II. Blue Nile group have two sisters (Kolom from Mengenza area, Kolom from Jebel El Nimir) and (Arfaa gadamak I, Arfaa gadamak II) beside Dura Abied. The second cluster have Red Sea group alone, accessions Baham Hazaz, Hazaz I appeared as sister where Hazaz III and Hazaz II were within the same group, with Hazaz II more genetically distant (Fig. 1).
Six out of the 17 sorghum accessions had unique bands that distinguished them from the others and could be useful as fingerprints. Kolom II had unique fragment that appear with the C-20, Ankoleeb Ahmer II accession had a unique fragment with UBC-101; Dura Abied accession had specific fragment with UBC-103; Baham Hazaz accession had a specific fragment with primer UBC-127; Abjaro Abu Soof accession had specific fragments with primer UBC-155 and Abjaro accession had specific fragment with primer UBC-157.
| Table 3: | Distance matrix among the 17 sorghum accessions studied |
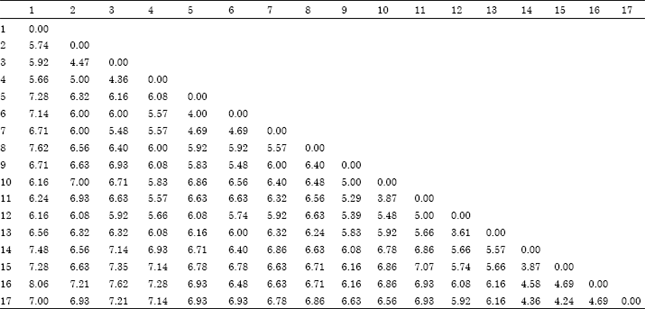 | |
Where; 1: Debeikri I, 2: Abjaro, 3: Abjaro Abu Soof, 4: Debeikri II, 5: Zinari Ahmer, 6: Ankoleeb Ahmer I, 7: Sifaira, 8: Ankoleeb Ahmer II, 9: Dura Abied, 10: Kolom I, 11: Kolom II, 12: Arfaa gadamak I, 13: Arfaa gadamak II, 14: Baham Hazaz, 15: Hazaz I, 16: Hazaz II and 17: Hazaz III | |
Accessions Arfaa gadamak I and Arfaa gadamak II from the Blue Nile region were the closest to each other, where the genetic distance matrix between them was (3.61), according to the genetic distance matrix shown in Table 3. Whereas, accessions Debeikri I and Hazaz II were the most distant among all (8.06).
DISCUSSION
In this study, the average number of amplification products produced per primer was 11 which is higher than finding in earlier reports (Iqbal et al., 2010; Amrapali et al., 2008; Mohamed et al., 2008). Accessions from Red sea region formed a distinct cluster by itself. According to Kachapur et al. (2009), it may be due to the presence of unique alleles inside those accessions and such alleles are important because, they may be diagnostic for particular regions with a genome specific to a particular type of sorghum. However, using these primers, access on specific DNA fingerprints were found for six accessions out of 17 studied. This will be of high value for sorghum breeders working in the improvement of the crop. Also, the primers (C-20, UBC-101, UBC-103, UBC-127, UBC-155 and UBC-157) that showed specific fingerprints can be used in between to screen longer spectrum of Sudanese sorghum accessions. An interesting finding from this study, are those accessions having the same name as classified earlier based on their morphological characteristics, appeared to be genetically different. This is clear for accessions Debeikri I and Debeikri II, although they clustered within one cluster, there were considerable genetic distance among them (Fig. 1, Table 3), the same was for Ankoleeb Ahmer I and Ankoleeb Ahmer II, although from the same region. Hazaz I, Hazaz II and Hazaz III similarly. Our results are in agreement with reports of Ayana et al. (2000) and Iqbal et al. (2010) who observed no genetic relationship with quantitative agro-morphological traits which shows no correlation between molecular markers and morphological traits. Ezzat et al. (2010) mentioned that different weather patterns, soil type and the location might cause agronomic differences among genotypes. Likewise, an insufficient relationship between RAPD markers and agronomic descriptors was also reported by Dahlberg et al. (2002). Different environmental conditions might affect the phenotypic traits, leading to clear differences even among identical genotypes (Shehzad et al., 2009). So, the indigenous or local names given by farmers of a region cannot be considered a good guide to the presence of diversity (Chakauya et al., 2006).
Agrama and Tuinstra (2003) reported that despite the sorghum is predominantly a self pollinated cultivars it has unusual amount of genetic diversity. Minkir et al. (1997) also concluded that the multiple origin theory of sorghum is also considered to be a factor contributing to the extensive genetic diversity observed in grain sorghum.
It may be suggested that RAPD marker have important implications for accessions identification and determination of hybridity and phylogenetic diversity of sorghum because of a requirement of simple agarose gel electrophoresis. Eventually, our study agrees with previous studies that RAPD analysis can be utilized with confidence in such applications (Udupa et al., 1998; Agrama and Tuinstra, 2003). So, it is concluded that RAPD technique proved to be useful to study genetic variation among the Sudanese sorghum accessions.
REFERENCES
- Agrama, H.A. and M.R. Tuinstra, 2003. Phylogenetic diversity and relationships among sorghum accessions using SSRs and RAPDs. Afr. J. Biotechnol., 2: 334-340.
CrossRefDirect Link - Amrapali, A.A., S.B. Sakhare, P.L. Kulwal and D.B. Dhumale, 2008. RAPD profile studies in sorghum for identification of hybrids and their parents. Inter. J. Integ. Biol., 3: 18-24.
Direct Link - Ayana, A., E. Bekele and T. Bryngelsson, 2000. Genetic variation in wild sorghum (Sorghum bicolor ssp. Verticilliflorum (L.) Moench) germplasm from Ethiopia assessed by Random Amplified Polymorphic DNA (RAPD) Hereditas, 132: 249-254.
CrossRefDirect Link - Bandyopadhyay, T., 2011. Molecular marker technology in genetic improvement of tea. Int. J. Plant Breed. Genet., 5: 23-33.
CrossRefDirect Link - Bethel, C.M., E.B. Sciara, J.C. Estill, J.E. Bowers, W. Hanna and A.H. Paterson, 2006. A framework linkage map of bermudagrass (Cynodon dactylon×transvaalensis) based on singledose restriction fragments. Theor. Appl. Genet., 112: 727-737.
CrossRefPubMedDirect Link - Bowers, J.E., C. Abbey and S. Anderson, C. Chang and X. Draye et al., 2003. A high-density genetic recombination map of sequence-tagged sites for Sorghum, as a framework for comparative structural and evolutionary genomics of tropical grains and grasses. Genetics, 165: 367-386.
CrossRefDirect Link - Chakauya, E., P. Tongoona, E.A. Matibiri and M. Grum, 2006. Genetic diversity assessment of sorghum landraces in Zimbabwe using microsatellites and indigenous local names. Int. J. Bot., 2: 29-35.
CrossRefDirect Link - Curtis, D.L., 1968. The relationship between the date of heading of Nigerian sorghums and the duration of growing season. J. Applied Ecol., 5: 215-226.
Direct Link - Dahlberg, J.A., X. Zhang, G.E. Hart and J.E. Mullet, 2002. Comparative assessment of variation among sorghum germplasm accessions using seed morphology and RAPD measurements. Crop Sci., 42: 291-296.
CrossRefPubMedDirect Link - Dufour, P., M. Deu, L. Grivet, A. D'Hont and F. Paulet et al., 1997. Construction of a composite sorghum genome map and comparison with sugarcane, a related complex polyploid. Theor. Appl. Genet., 94: 409-418.
CrossRefDirect Link - Ezzat, E.M., M.A. Ali and A.M. Mahmoud, 2010. Agronomic performance, genotype X environment interactions and stability analysis of grain sorghum (Sorghum bicolor L. Moench). Asian J. Crop Sci., 2: 250-260.
CrossRefDirect Link - El-Amin, H.K.A., N.B. Hamza and A.I. Abuali, 2011. Molecular and agronomical assessment of six wheat (Triticum aestivum L.) cultivars under salt-stress conditions. Int. J. Agric. Res., 6: 163-171.
CrossRefDirect Link - Fahima, T., G.L. Sun, A. Beharav, T. Krugman, A. Beiles and E. Nevo, 1999. RAPD polymorphism of wild emmer wheat populations, Triticum dicoccoides, in Israel. Theor. Appl. Genet., 98: 434-447.
CrossRefDirect Link - Iqbal, A., B. Sadia, A.I. Khan, F.S. Awan, R.A. Kainth and H.A. Sadaqat, 2010. Biodiversity in the sorghum (Sorghum bicolor L. Moench) germplasm of Pakistan. Genet. Mol. Res., 9: 756-764.
CrossRefDirect Link - Jessup, R.W., B.L. Burson, G. Burow, Y.W. Wang and C. Chang et al., 2003. Segmental allotetraploidy and allelic interactions in buffelgrass (Pennisetum ciliare (L.) Link syn. Cenchrus ciliaris L.) as revealed by genome mapping. Genome, 46: 304-313.
CrossRefDirect Link - Kachapur, R.M., P.M. Salimath and B.V.S. Reddy, 2009. Genetic diversity within sweet sorghum (Sorghum bicolor (L.) Monech) accessions as revealed by RAPD markers. J. Maharashtra Agric. Univ., 34: 38-42.
Direct Link - Ming, R., S.C. Liu, Y.R. Lin, J. da Silva and W. Wilson et al., 1998. Detailed alignment of saccharum and sorghum chromosomes: Comparative organization of closely related diploid and polyploid genomes. Genetics, 150: 1663-1682.
Direct Link - Minkir, A., P. Goldsbrough and G. Ejeta, 1997. RAPD based assessment of Genetic diversity in cultivated races of sorghum. Crop Sci., 37: 564-569.
Direct Link - Missaoui, A.M., A.H. Paterson and J.H. Bouton, 2005. Investigation of genomic organization in switchgrass (Panicum virgatum L.) using DNA markers. Theor. Appl. Genet., 110: 1372-1383.
CrossRefPubMedDirect Link - El-Awady, M., S.S. Youssef, E.E.M. Selim and M.M. Ghonaim, 2008. Genetic diversity among Sorghum bicolor genotypes using Simple Sequence Repeats (SSRs) markers. Arab. J. Biotechnol., 11: 181-192.
Direct Link - Nagaoka, T. and Y. Ogihara, 1997. Applicability of inter-simple sequence repeat polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD markers. Theoret. Applied Genet., 94: 597-602.
CrossRefDirect Link - Paterson, A.H., J.E. Bowers and B.A. Chapman, 2004. Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics. Proc. Natl. Acad. Sci., 101: 9903-9908.
CrossRefPubMedDirect Link - Patra, A.P., A.K. Mukherjee and L. Acharya, 2011. Comparative study of RAPD and ISSR markers to assess the genetic diversity of betel vine (Piper betle L.) in Orissa, India. Am. J. Biochem. Mol. Biol., 1: 200-211.
CrossRefDirect Link - Porebski, S., L.G. Bailey and B.R. Baum, 1997. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep., 15: 8-15.
CrossRefDirect Link - Pu, Z.E., Y.C. Hou, X.X. Xu, Z.H. Yan, Y.M. Wei, X.J. Lan and Y.L. Zheng, 2009. Genetic diversity among barley populations from West China based on RAMP and RAPD markers. Asian J. Plant Sci., 8: 111-119.
CrossRefDirect Link - Shehzad, T., H. Okuizumi, M. Kawase and K. Okuno, 2009. Development of SSR-based sorghum (Sorghum bicolor (L.) Moench) diversity research set of germplasm and its evaluation by morphological traits. Genet. Res. Crop Evol., 56: 809-827.
CrossRefDirect Link - Shiringan, A.L., 2009. Identification of genomic regions of Sorghum bicolor (L.) Moench linked to biofuel-related traits in grain x sweet sorghum recombinant inbred lines. Ph.D. Thesis, Faculty of Agricultural Sciences, Nutritional Sciences and Environmental Management, Justus Liebig University, Giessen, Germany.
- Smith, J.S.C. and O.S. Smith, 1992. Fingerprinting crop varieties. Adv. Agron., 47: 85-140.
CrossRefDirect Link - Soller, M. and J.S. Beckmann, 1983. Genetic polymorphism in varietal identification and genetic improvement. Theor. Appl. Genet., 67: 25-33.
CrossRefDirect Link - Subudhi, P.K. and N. Huang, 1999. RAPD mapping in a doubled haploid population of rice (Oryza sativa L.). Hereditas, 130: 41-49.
CrossRefPubMedDirect Link - Thangaraj, M., V. Prem, T. Ramesh and A.P. Lipton, 2011. RAPD fingerprinting and demonstration of genetic variation in three pathogens isolated from mangrove environment. Asian J. Biotechnol., 3: 269-274.
CrossRefDirect Link - Udupa, S.M., F. Weigand, M.C. Saxena and G. Kahl, 1998. Genotyping with RAPD and microsatellite markers resolves pathotype diversity in the ascochyta blight pathogen of chickpea. Theor. Appl. Genet., 97: 299-307.
CrossRefDirect Link - Whitkus, R., J. Doebley and M. Lee, 1992. Comparative genome mapping of sorghum and maize. Genetics, 132: 1119-1130.
Direct Link - Younis, R.A.A., M.F. Ahmed and M.M. El-Menshawy, 2007. Molecular genetic markers associated with salt tolerance in grain sorghum. Arab J. Biotechnol., 10: 249-258.
Direct Link - Fang, D.Q. and M.L. Roose, 1997. Identification of closely related citrus cultivars with inter-simple sequence repeat markers. Theor. Applied Genet., 95: 408-417.
CrossRefDirect Link