
Ashraf A. Khalil
Department of Protein Technology, Genetic Engineering and Biotechnology Research Institute, Mubarak City for Scientific Research and Technology Applications, Research Centers District, Borg Elarab, Alexandria, Egypt
Asian Journal of Biotechnology
Year: 2012 | Volume: 4 | Issue: 3 | Page No.: 108-119







ABSTRACT
Every year thousands of tones of Bolti fish (Tilapia nilotica) viscera are either discarded leading to environmental pollution and wastage of bioresources. This biomass has however a potential to generate considerable revenue and can be turned into a commercially viable business. The objective of this study was to evaluate the proximate protein characterization of the aqueous soluble phase of acidified and autolyzed Bolti fish viscera. Solubilization of fish viscera at natural pH (6.2); slight acidified pH (5.2) as well as acidified pH (4.2 and 3.2) was determined. Autohydrolysis reactions were conducted on freshly thawed viscera utilizing gradient temperatures and terminated at various time points (0, 8, 18, 32 and 47 h) by heat inactivation of the endogenous enzymes. Viscera were then characterized with respect to total nitrogen content, soluble protein, ammonia content, amino acid compositions and protein molecular weight distribution. Interestingly, endogenous enzyme activities were sufficient for obtaining a solubilization yield protein of up to 30 g L-1. SDS-PAGE profile showed that no characteristic band can be individualized for each hydrolysate and at the same time, greater variation was found only in the hydrolysate digested under initial pH 4.2 for 47 h. Amino acid analysis revealed that several amino acids were present in considerable amounts in the obtained hydrolysates, among them alanine, glutamic acid and proline. In addition, many hydrolysates contained useful quantities of the essential amino acids such as leucine, isoleucine, phenylalanine, histidine and valine and that may represent an interesting approach to upgrade these byproducts for food or feed purposes. In fact the results of our research would provide the incentive for commercial developments leading to large-scale and cost-effective production of fish peptone hydrolysates rich with essential amino acids from Bolti fish viscera.
PDF Abstract XML References Citation
Received: February 26, 2012;
Accepted: May 30, 2012;
Published: July 04, 2012
How to cite this article
Ashraf A. Khalil, 2012. Protein Characterization of the Aqueous Soluble Phase of Acidified and Autolyzed Bolti Fish (Tilapia nilotica) Viscera. Asian Journal of Biotechnology, 4: 108-119.
DOI: 10.3923/ajbkr.2012.108.119
URL: https://scialert.net/abstract/?doi=ajbkr.2012.108.119
DOI: 10.3923/ajbkr.2012.108.119
URL: https://scialert.net/abstract/?doi=ajbkr.2012.108.119
INTRODUCTION
The Nile tilapia (Tilapia nilotica) is a freshwater fish species in the Cichlids family that is hardy, adaptable and prolific. They are fast growing fish that are often farmed in ponds or tanks for commercial food use mainly in Africa and Asia. Tilapia fish is nutritious and forms a healthy part of a balanced diet to human beings that is high in protein as much of required proteins as in meat (Ghorbani and Mirakabad, 2010; Foh et al., 2011a). According to Egyptian General Authority for Fish Resources Development, about 477.458 tones of tilapia fish consisting of 55.6% of the total Egyptian production from fish culture in 2008. It is worthy to mention that Egyptian production represents 12% of the world farmed tilapia (2.121.009 tones) (FAO, 2008). According to the FAO fisheries statistics, total Middle East and North Africa (MENA) region tilapias production from fresh fisheries showed a steady rise along the period from 2004-2008. Egypt produces over 92% of total tilapia production in MENA region. Furthermore, Tilapia nilotica is of high economic value for aquaculture because it can be raised to uniform and marketable sizes within a short period of time. In earlier days of industrialized fisheries, fish sources appeared to be inexhaustible and by-products were considered as worthless and routinely discarded without an attempt of recovery. However, with a dramatically increasing world catch of fish these wastes became a problem when it started to accumulate in fishing harbors. Although, some people consider these wastes a hazard to health and an environmental problem to the fishing industry, many others considered it as significant source of protein (Nurdiyana and Mazlina, 2009). Usually, the major utilization of fish by-products is using them in production of powdered animal feed (Strom and Eggum, 1981).
To convert the by-products and wastes into more acceptable and marketable form, novel means of processing are required (Herpandi et al., 2011). One of the best approaches to maximize value of proteinaceous fish waste is the enzymatic processing to produce hydrolysates. Such hydrolysates of fish by-products can be utilized into four major applications, namely: (1) Ingredients in animal feed (Faid et al., 1997; Ouellet et al., 1997), (2) ingredients in food (Adler-Nissen, 1986; Mahmoud, 1994), (3) as the peptone ingredient in microbial growth media (Frokjaer, 1994; Gildberg et al., 1989; Vecht-Lifshitz et al., 1990) or (4) as materials for fertilizing (Kurbanoglu and Algur, 2002). Protein hydrolysates that are soluble in water and not coagulable by heat are called “peptones” (Green et al., 1977).
The economic value of peptones is higher than fish by-products (fish silage). One alternative is to apply such enzymatic hydrolysis to produce fish protein hydrolysates or fish powders that may be used as nitrogen and/or carbon source for bacterial growth and biomass production (Gildberg et al., 1989; Vecht-Lifshitz et al., 1990; Clausen et al., 1985; Dufosse et al., 2001). Many investigations stated that fish protein hydrolysates are efficient nitrogen sources for bacterial growth (Guerard et al., 2001) enzyme and bacteriocin production (Triki-Ellouz et al., 2003; Ghorbel et al., 2005; Deraz et al., 2011).
Generally, before adding to the bacterial growth media the proteins as nutrients should be hydrolyzed into peptides and amino acids. Enzymatic hydrolysis is an alternative approach for protein recovery and has wide use in food industry as products of high functionality and nutritive value (Foh et al., 2011b). It has, however, some disadvantages such as slow reaction rate and high cost due to the requirements of feedstock pretreatment, enzyme production and enzyme recovery. Furthermore, from an economic viewpoint, the protein hydrolysates produced by enzymatic hydrolysis are not applicable to the nutrient sources in the large scale production. On the contrary, acid-hydrolysis has the advantages of the low cost, short hydrolysis time and simple operation so that, it is applicable to industrial processes (See et al., 2011).
The major cost in the production of microbial cell mass is the growth substrate. The expansion of biotechnology will probably create an increasing demand for high-quality microbial growth media and stimulate the search for inexpensive sources. At present the nitrogen source is supplied by preparations from plants and slaughterhouse waste and it is usually the most expensive component of bacterial growth substrates. Despite its availability in large quantities and its low cost, fish waste has been used to a slight extent. When acidified fish viscera are stored they autolyzed to yield a liquid of soluble peptides and amino acids (Raa and Gildberg, 1976) and should therefore be an excellent source of nutrients for bacterial growth.
The production of fish protein hydrolysates using enzymes is not new. Traditionally, such hydrolysates are produced using the endogenous proteolytic enzymes present in muscle or fish viscera (Kristinsson and Rasco, 2000; Venugopal, 1994; Gildberg, 1994). Processes based on the use of endogenous proteases are usually referred to as “autolytic” processes. Such processes may either be run at neutral or slightly alkaline pH, thus exploiting the serine proteases of the intestine, trypsin, chymotrypsin and elastase, or at acidic pH, thus exploiting the carboxyl protease pepsin of the gastric juice (Pastoriza et al., 2004).
The objective of this study was to assess the potential utilization of tilapia fish viscera protein hydrolysates. The effect of autohydrolysis was investigated at various pH (6.2, 5.2, 4.2 and 3.2) for 8, 18, 32 and 47 h on the proximate protein compositions, such as total nitrogen (%), crude protein (%), soluble protein concentration (g L-1) and ammonia concentration (mg L-1). Amino acid concentrations were determined in samples autohydrolyzed for 47 h.
MATERIALS AND METHODS
Materials and chemicals: Bolti fish was purchased from the local market of Borg Elarab City and was transported in ice-filled containers to the laboratory within 15 min. The viscera were removed by hand as soon as possible and then stored in sealed plastic bags at -20°C until homogenated for a maximum period of 15 days. All chemicals used were of analytical grade.
Preparation of fish auto-hydrolysates: The frozen viscera masses were thawed at room temperature, grinded with 10% (v/w) of sterilized distilled water, then the homogenate was divided into 20 portions corresponding to four different pH values (6.2, 5.2, 4.2 and 3.2) and five incubation times (0, 8, 18, 32 and 47 h). The pH values used were adjusted by adding the necessary quantities of 4 N HCl. The auto-hydrolyses were carried out by maintaining the homogenates at 20°C, with orbital shaking at 100 rpm, for periods of 0, 8, 18, 32 and 47 h. After each incubation period, the auto-hydrolysates were heated at 85°C for 20 min to inactivate the endogenous enzymes (Guerard et al., 2001) and to facilitate the removal of fat residues present in the material (Bhaskar et al., 2007). The heated homogenates were centrifuged at 6000 rpm for 15 min to obtain the corresponding supernatants. The supernatants (hereinafter termed viscera hydrolysates) were stored at -20°C until used.
Chemical analysis of Bolti viscera hydrolysates (BVHs): Total nitrogen contents of BVHs were determined following micro-Kjeldahl method according to AOCS (1998). Crude protein content was estimated by multiplying total nitrogen content by the conversion factor of 6.25 (Havilah et al., 1977). Soluble protein contents were determined according to the method of Lowry et al. (1951). Ammonia concentrations were measured using ammonia assay kit AA0100 (Sigma-Aldrish). Analyses were performed in triplicate.
Amino acid composition of BVHs: Amino acids were determined as described by Pellet and Young (1980). The analysis was performed in the Central Lab, Mubarak City for Scientific Research, Egypt using S433 amino acid analyzer (Syknm). The technique was based on the separation of the amino acids using strong cation exchange chromatography followed by the ninhydrin colour reaction and photometric detection at 570 nm. Standard amino acids were used for comparison of resulting profiles, allowing quantitation of amino-acid residues. The samples were hydrolyzed with 6 N HCl at 110°C in Teflon capped vials for 24 h. After vacuum removal of HCl, the residues were dissolved in a lithium citrate buffer, pH 2.2. Twenty microliter of the solution were loaded onto the cation exchange column (pre-equilibrated with the same buffer), then four lithium citrate buffers with pH values of 2.2, 2.8, 3.3 and 3.7, respectively, were successively applied to the column at a flow rate of 20 mL min-1. The ninhydrin flow rate was 10 mL h-1 under these conditions and a typical analysis required 160 min.
Characterization of BVHs by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE): Auto-hydrolysated samples under various initial pH of 6.2, 5.2, 4.2 and 3.2 for 47 h were resolved in SDS-PAGE gel. One volume of sample was mixed with one volume of an SDS-PAGE sample buffer containing 62.5 mM tris-HCl, pH 6.8, 10% (v/v) glycerol, 2% (w/v) SDS, 5% (v/v) β-mercaptoethanol and 0.025% (w/v) bromphenol blue. The samples were incubated for 10 min in a boiling water bath. SDS-PAGE was conducted essentially according to Laemmli (1970), using a separating gel (15%), in a Bio-Rad Mini Protean II system. The gels were stained by Coomassie blue R-250. The protein standard used was “PageRuler™ unstained protein ladder” purchased from Fermentas contained a mixture of 14 highly purified recombinant proteins. Molecular masses of the protein ladder ranging from 10-200 kDa.
RESULTS
Proximate protein characterization: The changes in proximate protein characteristics such as nitrogen contents (%), crude protein contents (%), soluble protein content (%) ammonia concentration (mg mL-1) after autohydrolysis of BVHs under various initial pH of 6.2, 5.2, 4.2 and 3.2 for 0, 8, 18, 32 and 47 h are shown in Fig. 1-4.
The nitrogen contents were measured by the micro-Kjeldahl method and expressed as a percentage of total nitrogen content in the dry matter of fish viscera. As indicated in Fig. 1 and 2, the proximate total nitrogen and crude protein percentage of the fish hydrolysates were comparable in most samples irrespective of the initial pH used or time of hydrolysis. However, some differences were noticed in the samples hydrolyzed under pH 3.2 for various times. Another interesting notice was also recorded in sample hydrolysated for 32 h under pH 3.2 which contained almost twofold concentration.
As presented in Fig. 3, soluble protein contents of viscera hydrolysates were not greatly affected either by initial pH or hydrolysis time with values ranged from 23.41 (pH 4.2 for 8 h) to 30.9 (pH 4.2 for 47 h) g L-1. It has been noticed that long hydrolysis time (47 h) increases soluble protein contents in viscera hydrolysates with values of 27.47, 29.2, 30.9 and 30.05 g L-1 under pH of 6.2, 5.2, 4.2 and 3.2, respectively.
Ammonia concentration was one of the factors that may contribute to observed differences in hydrolysis performance of BVHs at various pH. Ammonia concentrations in fish hydrolysates under various initials pH for various times are shown in Fig. 4.
It was found that untreated hydrolysates contained approximately comparable range of the ammonia ranged from 0.67-0.80 mg mL-1. Autohydrolysis of fish viscera for 47 h under initial pH of 6.2 and 4.2 led to a considerable increase of ammonia concentrations of 1.37 and 2.93 mg mL-1, respectively.
Another considerable increase in ammonia concentration (1.37 mg mL-1) was reported in Bolti fish samples hydrolysated under initial pH 6.2 for 18 h.
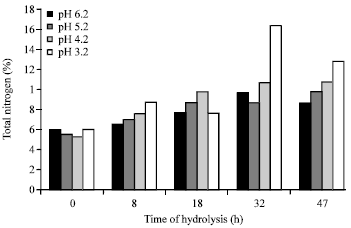 | |
| Fig. 1: | Total nitrogen content (%) in Bolti fish viscera autohydrolyzed under various pH for various times |
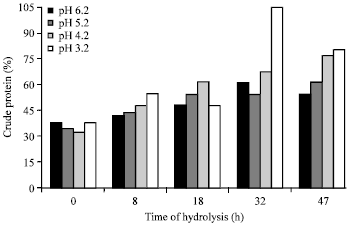 | |
| Fig. 2: | Crude protein content (%) in Bolti fish viscera autohydrolyzed under various pH for various times |
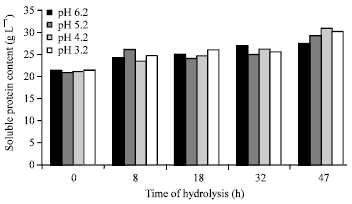 | |
| Fig. 3: | Soluble protein content (g L-1) in Bolti fish viscera autohydrolyzed under various pH for various times |
Amino acid profile: The results showed that the amino acid profiles were generally higher in essential amino acids compared to the suggested pattern of requirement by FAO/WHO (1990) for adult humans and amino acid requirements of juvenile common carp, as listed by NRC (1993).
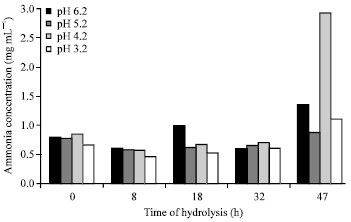 | |
| Fig. 4: | Total free ammonia (mg mL-1) in autohydrolyzed Bolti fish viscera under various pH for various times |
| Table 1: | Amino acid composition of Bolti fish viscera hydrolysated under various pH for 47 h |
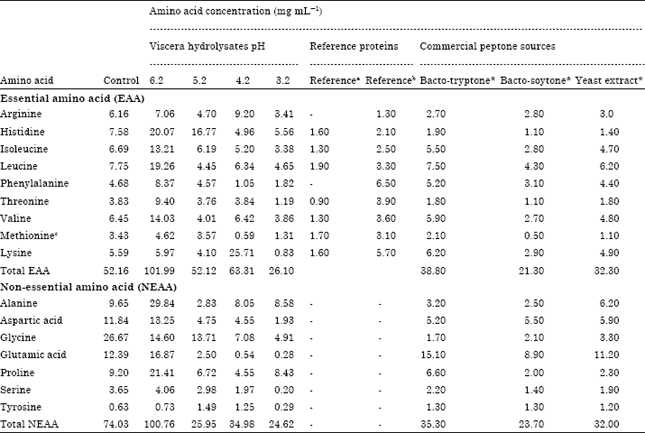 | |
| aSuggested profile of essential amino acid requirements for adults (FAO/WHO) (NRC, 1993). b Essential amino acid requirements of common carp according to NRC (1993). cMethionine+cysteine. *Amino acid compositions as described in supplier datasheets: BactoTMTryptone from casein (Beckton Dickinson, BD), BactoTMSoytone from soybean (BD), Yeast extract (Oxoid) | |
The amino acid composition of fish hydrolysates after 47 h of hydrolysis under initial pH of 6.2 were several orders of magnitude higher concentration in most of the amino acids compared to amino acid compositions of Bacto-Tryptone, Bacto-Soytone and yeast extract (Table 1). The highest amino acids concentrations were shown for alanine, glycine, proline, histidine, leucine, glutamic acid, valine and isoleucine with values of 29.84, 26.67, 21.41, 20.07, 19.26, 16.87, 14.03 and 13.21 mg mL-1, respectively. For nonessential amino acids aromatic serine showed the lowest level (0.20 mg mL-1).
The fish viscera hydrolyzed for 47 h at pH 3.2 showed comparable total concentration of essential (26.1) and non-essential (24.62 mg mL-1) amino acids, respectively. In both cases, the total amino acid yield was slightly higher to the total amino acid concentration of Bacto-Soytone which contained 21.30 mg mL-1 of essential amino acids and 23.70 mg mL-1 of non-essential amino acids.
The comparison between concentrations of essential amino acids of hydrolysate with initial pH 4.2 and commercial peptones (Bacto-Tryptone, Bacto-Soytone and yeast extract) showed almost two fold concentrations in hydrolysate. The first, second and third highest concentrations of essential amino acids were lysine (25.71 mg mL-1), histidine (20.07 mg mL-1) and leucine (19.26 mg mL-1). The level of most of the essential amino acids in hydrolysates was found to be adequate compared to those of reference protein (FAO/WHO, 1990).
SDS electrophoresis: Figure 5 shows the electrophoretic protein profile of the digested fish viscera under various initial pH of 6.2, 5.2, 4.2 and 3.2 for 47 h.
The electrophoretic patterns of various hydrolysates showed the presence of one intensive band with a molecular mass of less than 14.2 kDa in fish peptone hydrolysates with initial pH 6.2 (lanes 2 and 3) and with initial pH 3.2 (lanes 7 and 8) after 8 and 18 h, respectively.
The four hydrolysates also showed a couple of bands in the range of 150 and 200 kDa which might be proteins that not totally hydrolyzed by the endogenous enzyme. From the analysis of the gels it can be observed that no characteristic band can be individualized for each hydrolysate and, at the same time, greater variation was found only within the hydrolysate digested for 47 h with initial pH 4.2 (lane 11).
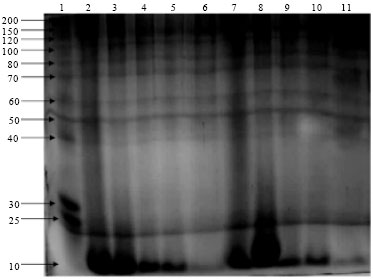 | |
| Fig. 5: | SDS-PAGE of fish hydrolysates. A total of 7.5 mL of viscera hydrolysate was applied to each well. Lane 1: Protein molecular marker (molecular masses are indicated). Lanes 2-5: Viscera hydrolysates produced with pH 6.2 for 8, 18, 32 and 47 h, respectively. Lane 6: Viscera hydrolysates produced under pH 5.2 for 47 h. Lanes 7-10: Viscera hydrolysates produced under pH 3.2 for 8, 18, 32 and 47 h, respectively. Lane 11: Viscera hydrolysates produced under pH 4.2 for 47 h |
DISCUSSION
There are two types of processes for the production of fish protein hydrolysates, namely autolysis and accelerated hydrolysis process (Mohr, 1977). Accelerated hydrolysis is generally a more complex process and the cost of enzymes involved may influence economical aspects and commercial viability of the process. Autolysis via digestive enzymes is an industrial method for preparing fish protein concentrate as described and documented in the literature (Gildberg et al., 1989; Batista and Nunes, 1997; Kim et al., 1999; Mohr, 1980). The autolytic process depends on the action of digestive enzymes of the fish itself.
Based on the nature of catalytic site, digestive proteinases from marine animals are further classified into four categories as acid or aspartate proteinases, serine proteinases, thiol or cysteine proteinases, or metalloproteinase’s. The acid or aspartyl proteinases are a group of endopeptidases characterized by high activity and stability at acidic pH such as pepsin, chymosin and gastricsin (Simpson, 2000). Furthermore, digestive cysteine or thiol proteinases are most active at close to neutral to acidic pH (6.0-3.0) and inactive at alkaline pH. Common examples of digestive thiol proteins from marine animals are cathepsin B, cathepsin L and cathepsin S (Simpson, 2000; Wasswa et al., 2007). In this study, we have utilized a great range of environmental conditions in which these proteinases are active which can explain the differences in amino acid concentrations obtained after 47 h with various initial pH.
Prepared media fortified by visceral homogenates of Bolt fish resulted in higher growth rates and biomass yields than those obtained with costly commercial media, specifically media recommended for culturing and large-scale production of lactic acid bacteria (Deraz et al., 2011). One possible explanation for the good performance of the fish peptones could be their amino acid composition with high levels of branched amino acids such as, leonine, valise and isoleucine which are essential for L. sakei strain (Aspmo et al., 2005).
Amino acid composition did not display a homogeneous trend; levels fluctuated compared to control. A similar result was observed by Morales-Ulloa and Oetterer (1997) and Arason (1994). The nutritive value of a protein depends primarily on the capacity to satisfy the needs from nitrogen and essential amino acids (Pellet and Young, 1980). According to Strom and Eggum (1981), lysine, cystine and methionine are the most important amino acids for fish from a nutritional viewpoint. From the results presented, all fish viscera hydrolysates satisfy amino acid requirements according to FAO and Nile Tilapia standards (NRC, 1993; Kamara et al., 2011). Furthermore, when considering amino acid limiting level, the 30% minimum requirement in fish diets rather than FAO standards (Tacon, 1994); this study shows that all hydrolysates fulfilled this specific requirement. In addition, our study revealed the practical relevance of Bolti viscera hydrolysates as a good source of desirable quality essential and non essential amino acids in food, feed or bacterial growth medium applications. Similar results were obtained by Goda et al. (2007) from hydrolysis of muscles and gonads of wild and captive Nile tilapia.
Ammonia is a product of hydrolysis or anaerobic degradation of protein-rich substrates such as fish sludge. Ammonia concentrations below 200 mg L-1 are beneficial to anaerobic process (Chen et al., 2008) but when present at high concentrations, it can cause growth inhibition. There are two forms of ammoniacal nitrogen in aqueous solution, ammonium (NH4+) and Free Ammonia (FA). FA is reported to be the main cause of inhibition since it is highly membrane-permeable and can therefore diffuse passively into the cell causing proton imbalance and/or potassium deficiency (Gallert et al., 1998). Therefore, ammonia is one of the important products of protein degradation which should be controlled especially for application in fish peptone hydrolysates in growth medium.
The word peptone principally denotes a protein hydrolysis, whereas, several commercial peptones and certainly the fish hydrolysates discussed in this study contains much more than just peptides and amino acids and are, as such, more than just a source of nitrogen (Shapawi et al., 2011). It is also important to note that some of the substrates used in this study are insoluble particles containing complexes of bimolecular. Hydrolysis of one of the major compounds in these complexes, i.e. protein, will liberate much more to the solution than just peptides and amino acids. Therefore, further chemical composition studies are going to determine other component in the current hydrolysates. More extensive research to identify the most specific and promising enzymes in Bolt fish viscera and determination of the optimal conditions for their use is of utmost importance.
CONCLUSION
The endogenous enzymes of Bolti fish viscera are capable to hydrolyze viscera waste to form nitrogen-rich products under mild reaction conditions. Hydrolysis yield can be substantially improved by using a suitable initial pH in a period of a few hours. Namely, several amino acids were present with considerable amounts in the obtained hydrolysate, among them aspartic acid, alanine, glutamic acid, glycine, lysine, arginine, valine and leucine. Literature comparison with three commercial peptones described for bacterial growth media indicates that hydrolysates have potential to be used as peptone source in formulating microbiological media, specifically recommended for culturing and large-scale production of lactic acid bacteria which requires diversified peptidic sources. The obtained hydrolysates contained most of the common amino acids and can serve as a starting material for various industrial applications such as upgrading these byproducts for food or feed purposes. In fact the results of such research would provide the incentive for commercial developments leading to large-scale and cost-effective production of fish peptone hydrolysates rich with essential amino acids from Bolti fish viscera.
REFERENCES
- Aspmo, S.I., S.J. Horn and V.G.H. Eijsink, 2005. Hydrolysates from Atlantic cod (Gadus morhua L.) viscera as components of microbial growth media. Process Biochem., 40: 3714-3722.
CrossRef - Bhaskar, N., V.K. Modi, K. Govindaraju, C. Radha and R.G. Lalitha, 2007. Utilization of meat industry by products: Protein hydrolysate from sheep visceral mass. Bioresour. Technol., 98: 388-394.
PubMed - Chen, Y., J.J. Cheng and K.S. Creamer, 2008. Inhibition of anaerobic digestion process: A review. Bioresour. Technol., 99: 4044-4064.
CrossRefDirect Link - Clausen, E., A. Gildberg and J. Raa, 1985. Preparation and testing of an autolysate of fish viscera as growth substrate for bacteria. Applied Environ. Microbiol., 50: 1556-1557.
Direct Link - Deraz, S.F., G.F. El-Fawal, S.A. Abd-Ellatif and A.A. Khalil, 2011. Autohydrolysed Tilapia nilotica fish viscera as a peptone source in bacteriocin production. Indian J. Microbiol., 51: 171-175.
CrossRef - Dufosse, L., D. de la Broise and F. Guerard, 2001. Evaluation of nitrogenous substrates such as peptones from fish: A new method based on gompertz modeling of microbial growth. Curr. Microbiol., 42: 32-38.
CrossRef - Faid, M., A. Zouiten, A. Elmarrakchi and A. Achkari-Begdouri, 1997. Biotransformation of fish waste into a stable feed ingredient. Food Chem., 60: 13-18.
CrossRefDirect Link - Foh, M.B.K., M.T. Kamara, I. Amadou, B.M. Foh and X. Wenshui, 2011. Chemical and physicochemical properties of tilapia (Oreochromis niloticus) fish protein hydrolysate and concentrate. Int. J. Bot., 6: 21-36.
CrossRefDirect Link - Foh, M.B.K., I. Amadou, M.T. Kamara, B.M. Foh and W. Xia, 2011. Effect of enzymatic hydrolysis on the nutritional and functional properties of nile tilapia (Oreochromis niloticus) proteins. Am. J. Biochem. Mol. Biol., 1: 54-67.
CrossRefDirect Link - Frokjaer, S., 1994. Use of hydrolysates for protein supplementation. Food Technol., 48: 86-88.
Direct Link - Gallert, C., S. Bauer and J. Winter, 1998. Effect of ammonia on the anaerobic degradation of protein by a mesophilic and thermophilic biowaste population. Applied Microbiol. Biotechnol., 50: 495-501.
PubMed - Ghorbani, M. and H.Z. Mirakabad, 2010. Factors influencing on trout production in Khorasan Razavi Province. Trends Agric. Econ., 3: 19-27.
CrossRef - Ghorbel, S., N. Souissi, Y. Triki-Ellouz, L. Dufosse, F. Guerard and M. Nasri, 2005. Preparation and testing of Sardinella protein hydrolysates as nitrogen source for extracellular lipase production by Rhizopus oryzae. World J. Microbiol. Biotechnol., 21: 33-38.
CrossRef - Goda, A.M.A.S., O.M. El-Husseiny, G.M. Abdul-Aziz, A. Suloma and Y.H. Ogata, 2007. Fatty acid and free amino acid composition of muscles and gonads from wild and captive tilapia Oreochromis niloticus (L.) (Teleostei: Perciformes): An approach to development broodstock diets. J. Fish. Aquatic Sci., 2: 86-99.
CrossRefDirect Link - Guerard, F., L. Dufosse, D.D.L. Broise and A. Binet, 2001. Enzymatic hydrolysis of proteins from yellowfin tuna Thunnus albacares wastes using Alcalase. J. Mol. Catal. B. Enzym., 11: 1051-1059.
CrossRef - Herpandi, N. Huda and F. Adzitey, 2011. Fish bone and scale as a potential source of halal gelatin. J. Fish. Aquat. Sci., 6: 379-389.
CrossRefDirect Link - Kamara, M.T., I. Amadou, Z. Kexue, M.B. Kelfala Foh and Z. Huiming, 2011. The influence of debittering and desalting on defatted foxtail millet (Setaria italica L.) protein hydrolysate. Am. J. Biochem. Mol. Biol., 1: 39-53.
CrossRefDirect Link - Kristinsson, H.G. and B.A. Rasco, 2000. Fish protein hydrolysates: Production, biochemical and functional properties. Crit. Rev. Food Sci. Nutr., 40: 43-81.
CrossRefPubMedDirect Link - Kurbanoglu, E.B. and O.F. Algur, 2002. The influence of ram horn hydrolyzate on the crop yield of the mushroom Agaricus bisporus. Sci. Horticult., 94: 351-357.
CrossRef - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mohr, V., 1980. Enzymes technology in the meat and fish industries. Process Biochem., 15: 18-23.
Direct Link - Nurdiyana, H. and M.K.S. Mazlina, 2009. Optimization of protein extraction from fish waste using response surface methodology. J. Applied Sci., 9: 3121-3125.
CrossRefDirect Link - Ouellet, D.R., J.R. Seoane, D.M. Veira and J.G. Proulx, 1997. Effects of supplementation with fish meal or fish protein hydrolysate on growth, nutrient digestibility and rumen fermentation of growing cattle fed grass silage. Anim. Feed Sci. Technol., 68: 307-326.
Direct Link - Pastoriza, L., G. Sampedro, M.L. Cabo, J.J.R. Herrera and M. Bernardez, 2004. Solubilisation of proteins from rayfish residues by endogenous and commercial enzymes. J. Sci. Food Agri., 84: 83-88.
CrossRef - Raa, J. and A. Gildberg, 1976. Autolysis and proteolytic activity of cod viscera. Int. J. Food Sci. Technol., 11: 619-628.
CrossRef - See, S.F., L.L. Hoo and A.S. Babji, 2011. Optimization of enzymatic hydrolysis of Salmon (Salmo salar) skin by Alcalase. Int. Food Res. J., 18: 1359-1365.
Direct Link - Shapawi, R., S. Mustafa and W.K. Ng, 2011. A comparison of the growth performance and body composition of the humpback grouper, Cromileptes altivelis fed on farm-made feeds, commercial feeds or trash fish. J. Fish. Aquat. Sci., 68: 523-534.
CrossRefDirect Link - Strom, T. and B.O. Eggum, 1981. Nutritional value of fish viscera silage. J. Sci. Food Agric., 32: 115-120.
CrossRef - Triki-Ellouz, Y., B. Ghorbel, N. Souissi, S. Kammoun and M. Nasri, 2003. Biosynthesis of protease by Pseudomonas aeruginosa MN7 grown on fish substrate. World J. Microbiol. Biotechnol., 19: 41-45.
CrossRef - Wasswa, J., T. Jian and G. Xiao-Hong, 2007. Desalting fish skin protein hydrolysates using macroporous adsorption resin. Am. J. Food Technol., 2: 406-413.
CrossRefDirect Link - Vecht-Lifshitz, S.E., K.A. Almas and E. Zomer, 1990. Microbial growth on peptones from fish industrial wastes. Lett. Applied Microbiol., 10: 183-186.
CrossRef