
Thinnakorn Lao-Ong
Research Group for Pharmaceutical Activities of Natural Products using Pharmaceutical Biotechnology, Faculty of Pharmaceutical Sciences, National Research University-Khon Kaen University, Khon Kaen, 40002, Thailand
Waranya Chatuphonprasert
Research Group for Pharmaceutical Activities of Natural Products using Pharmaceutical Biotechnology, Faculty of Pharmaceutical Sciences, National Research University-Khon Kaen University, Khon Kaen, 40002, Thailand
Kanokwan Jarukamjorn
Research Group for Pharmaceutical Activities of Natural Products using Pharmaceutical Biotechnology, Faculty of Pharmaceutical Sciences, National Research University-Khon Kaen University, Khon Kaen, 40002, Thailand
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 4 | Page No.: 271-276







ABSTRACT
Berberine is a major active constituent in several medicinal plants such as Coscinium fenestratum, Berberis aristata, Coptis japonica and Coptis chinensis. Berberine possesses a wide range of biochemical and pharmacological activities including anti-inflammation, anti-microorganism, anti-cancer activities and anti-diabetes. In this study, we determined the effect of berberine, in combination with β-naphthoflavone (BNF, a pro-carcinogen), on the expression of sex-hormone synthesis genes at mRNA level including CYP17, CYP19, 3β-HSD, 17β-HSD1, 17β-HSD3 in mouse testes. The single treatment of berberine up-regulated the expression of testicular CYP17, 3β-HSD and 17β-HSD1 mRNA. BNF up-regulated only 17β-HSD1 mRNA. Moreover, the combination of berberine and BNF up-regulated the expression of CYP17 and 3β-HSD. These observations suggested that berberine might disturb sex-hormone synthesis pathway; consequently possibly resulted in modification of estrogen or testosterone synthesis. Therefore, a caution should be noted for the use of berberine as an alternative medicine, especially at high dose or long period.
PDF Abstract XML References Citation
Received: March 08, 2013;
Accepted: March 25, 2013;
Published: July 10, 2013
How to cite this article
Thinnakorn Lao-Ong, Waranya Chatuphonprasert and Kanokwan Jarukamjorn, 2013. Berberine Disturbs the Expression of Sex-hormone Regulated Genes in β-naphthoflavone-induced Mice. Journal of Biological Sciences, 13: 271-276.
DOI: 10.3923/jbs.2013.271.276
URL: https://scialert.net/abstract/?doi=jbs.2013.271.276
DOI: 10.3923/jbs.2013.271.276
URL: https://scialert.net/abstract/?doi=jbs.2013.271.276
INTRODUCTION
In steroidogenesis (Fig. 1), the first step is converting cholesterol into pregnenolone by cytochrome P450 (CYP11α) encodes cytochrome P450scc (cholesterol side-chain cleavage enzyme) (Miller, 1988; Stocco, 2000) Afterwards, pregnenolone is converted to progesterone by 3β-hydroxysteroid dehydrogenase (3β-HSD). The human cytochrome P450c17α enzyme (CYP17) possesses two enzymatic actions namely 17α-hydroxylase and 17,20-lyase which participate in two different catalytic steps in the steroid hormone synthesis pathway (Zuber et al., 1986). Pregnenolone and progesterone are converted by 17α-hydroxylase to 17-hydroxypregnenolone and 17-hydroxyprogesterone, respectively. After that, the 17,20-lyase turns both 17-hydroxypregnenolone and 17-hydroxyprogesterone into dehydroepiandrosterone (DHEA) and androstenedione, respectively. The expression of human CYP17 is constitutively expressed in female ovarian theca cells and adrenal cortex. Rare mutations in the coding region of CYP17 have been found in patients with 17α-hydroxylase/17,20-lyase deficiency, resulting in various clinical profiles such as congenital adrenal hyperplasia, abnormal sexual development, osteoporosis and irregular menstruation (Yanase et al., 1991; Yanase, 1995). The 3β-hydroxysteroid dehydrogenase-isomerase enzyme (3β-HSD) functions by converting pregnenolone to progesterone. Moreover, 3β-HSD can convert progesterone and DHEA to androstenedione (Bates et al., 2005). The aromatase P450 enzyme (CYP19) is a catalyzer for the conversion of androstenedione to estrone, or modification of testosterone to estrogen in many tissues (Morishima et al., 1995). The reproductive systems of rats, mice and human are affected by aromatase (CYP19) inhibition. Deficiency of CYP19 enzyme in male mice severely disturbed spermatogenesis resulted in infertility and consequently deteriorated ability to breed pups. In females, CYP19 deficiency likewise resulted in lack of estrogen followed by pseudohermaphroditism and masculinization in adolescence (Carani et al., 1997).
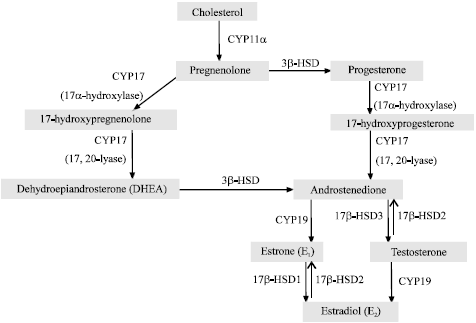 | |
| Fig. 1: | Sex-hormone synthesis pathway |
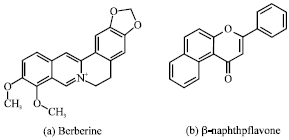 | |
| Fig. 2: | Structure of (a) Berberine and (b) β-naphthoflavone |
The latter pathway culminates in the formation of testosterone via conversion of androstenedione by 17β-hydroxysteroid via conversion of androstenedione by 17β-hydroxysteroid dehydrogenase type 3 (17β-HSD3) while 17β-hydroxysteroid dehydrogenase type 1 (17β-HSD1) converts the formation of estrone to estradiol (Sha et al., 1996).
Berberine (Fig. 2) is a major plant alkaloid present in several Indian and Chinese herbal plants; Coscinium fenestratum, Berberis aristata, Berberis vulgaris, Hydrastis Canadensis, Tinospora cordifolia, Coptis japonica and Coptis chinensis (Rojsanga et al., 2006; Sato and Yamada, 1984). The extensive researches demonstrated a number of biological and pharmacological benefits of berberine, i.e., anti-inflammation (Kuo et al., 2004), anti-microorganism (Schmeller et al., 1997), anti-cancer activities (Wu et al., 1999; Letasiova et al., 2006; Piyanuch et al., 2007) and anti-diabetes (Zhang et al., 2008; Zhang et al., 2011).
Our previous study noted the effects of berberine on β-naphthoflavone (BNF)-induced CYP1A expression in both primary mouse hepatocytes and mouse livers (Chatuphonprasert et al., 2011). The BNF-induced expressions of mouse CYP1A mRNA and protein and the related enzyme activities were significantly suppressed by berberine. In addition, berberine significantly lowered the level of BNF-induced lipid peroxidation in mouse hepatic microsome (Chatuphonprasert et al., 2011). The study suggested that the use of berberine as an alternative medicine might bring more advantages due to its ability to decrease the risk of carcinogenesis from induction of CYP1A expression and inhibitory effect on lipid peroxidation activity. In this study, we determined the effect of berberine, in combination with BNF (as pro-carcinogen), on the expression of sex-hormone synthesis genes at mRNA level including CYP17, CYP19, 3β-HSD, 17β-HSD1, 17β-HSD3 in mouse testes. Completeness of the study provided more detail of utilization of berberine. In contrast, if there is any negative effect found, precaution of berberine would be concerned.
MATERIALS AND METHODS
Chemicals: Berberine choride (Ber), β-Naphthoflavone (BNF) and corn oil were supplied by Sigma-Aldrich Chemical (St. Louis, MO). ReverTraAce was a product of Toyobo (Osaka, Japan). Trizol® reagent, random primers (hexadeoxyribonuclotide mixture), Taq DNA polymerase, RNase inhibitor and dNTP mixtures were products of InvitrogenTM (Carlsbad, CA). All other laboratory chemicals were of the highest purity available from commercial suppliers. Forward and reverse primers of mouse CYP17, CYP19, 3β-HSD, 17β-HSD1, 17β-HSD3 and GAPDH genes were synthesized by Bio Basic, Inc. (Markham Ontario, Canada). The primers of each gene are shown in Table 1.
| Table 1: | Primers sequences for PCR |
 | |
| Table 2: | Conditions of PCR cycles |
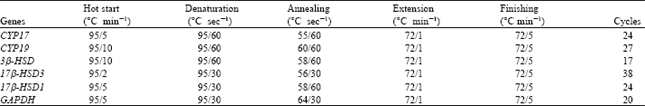 | |
Animals and treatments: Male C57BL/6 mice were supplied by National Laboratory Animal Center, Mahidol University, Nakhon Pathom, Thailand. All mice were housed in the Animal Unit of Faculty of Pharmaceutical Sciences, Khon Kaen University. Mice were housed in the Northeast Laboratory Animal Center (Khon Kaen University, Khon Kaen, Thailand) under the supervision of a certified laboratory veterinarian. They were treated according to a research protocol approved by the Animal Ethics Committee for Use and Care of Khon Kaen University, Khon Kaen, Thailand (AEKKU 06/2553). At all times, the mice were housed on wood chip bedding in polysulfone cages with water and commercial animal diet supplied ad libitum and were acclimated for at least 7 days in housing with a 12 h dark/light cycle under controlled temperature (23±2°C) and humidity (45±2%) before dosing. Seven week-old mice were orally fed daily with 7.5 mg/kg/day of berberine (Ber) for 7 days and/or intraperitoneally given 30 mg/kg/day of BNF in the last 3 days. The control group was orally given corn oil daily for 7 days. The mice were sacrificed at the 24 h after the last treatment and the livers were excised immediately for preparation of total RNA from testes.
Semi-quantitative reverse transcription polymerase chain reaction: Mouse CYP17, CYP19, 3β-HSD, 17β-HSD3, 17β-HSD1 and GAPDH mRNAs were semi-quantified by RT-PCR. Testicular total RNA was reverse-transcribed using random primer and ReverTraAce®, then cDNA was amplified. The conditions of PCR cycle were followed by the method of Degawa et al. (2006), Sha et al. (1996), Udomsuk et al. (2011) and Chatuphonprasert et al. (2009) with some modifications (Table 2). After separation of the PCR products by 2% agarose gel electrophoresis, the target cDNA bands were detected under ultraviolet light in the presence of ethidium bromide and semi-quantified by Gel documentation (Ingenius syngene bio-imagimg, model: IngeniusL) and Gene Tool Match program (Syngene, Lab Focus Co. Ltd., Cambridge, UK). The mRNA levels of the target genes were normalized to that of a house keeping gene, glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
RESULTS AND DISCUSSION
Modification of sex-hormone related genes including CYP17, 3β-HSD, CYP19, 17β-HSD3 and 17β-HSD1 and GAPDH, at the transcriptional level after treatment with berberine and/or BNF were examined in testes. The single treatment of berberine and the co-treatment with BNF significantly up-regulated expression of CYP17 and 3β-HSD mRNA (Fig. 3). Polymorphism of CYP17 gene influences breast cancer in young women and up-regulated transcription of CYP17 might affect the synthesis of estrogen and testosterone (Bergman-jungestrom et al., 1999). In addition, the retinoids stimulated mRNA abundance and promoter function of CYP17 was found in polycystic ovary syndrome theca cell (Wickenheisser et al., 2005). Hence, the enhancing effect of berberine and the co-treatment with BNF might be aware due to CYP17 induction effect. 3β-HSD involves in several step of sex-hormone synthesis pathway (Zuber et al., 1986; Bates et al., 2005). Therefore, berberine might disturb sex-hormone synthesis, consequently possibly resulted in modification of circulating pattern of estrogen or testosterone in the body. Moreover, the expression of 17β-HSD1 mRNA was significantly up-regulated after single treatment with berberine or BNF but there was no change in co-treatment group (Fig. 4). The up-regulated expression of 17β-HSD1 might be involved in estradiol metabolism pathway by an increase the conversion of estradiol.
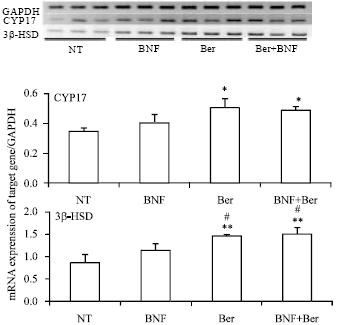 | |
| Fig. 3: | Expression pattern of testicular CYP17 and 3β-HSD mRNA in male mice Seven week-old mice were orally fed daily with 7.5 mg/kg/day day of berberine (Ber) for 7 days and/or intraperitoneally given 30 mg/kg/day day of β-naphthoflavone (BNF) in the last 3 days. The control group (NT) was orally given corn oil daily for 7 days. The relative mRNA expression levels were normalized by that of GAPDH. The data are presented as the Mean±SD (n = 3-4) from at least 2 independent experiments. A significant difference was examined by ANOVA with Tukey post hoc test. *p<0.05, **p<0.001 compared to NT group; #p<0.05 compared to BNF group |
Hepatic cytochrome P450 1A1 (CYP1A1) and cytochrome P450 1B1 (CYP1B1) play key roles in catalyze estrogen to 2 and 4-hydroxyestradiol and convert to methoxyestradiol by catechol-O-methyltransferase (Liehr, 2000; Dawling et al., 2003). The previous study showed that BNF induced CYP1A1 and CYP1B1 activities. Activations of these enzymes lead to increase estradiol metabolism and resulted in up-regulated expression of 17β-HSD1. The unchange of 17β-HSD1 by the co-treatment resulted from the feedback inhibition of estradiol metabolism by methoxyestradiol (Liehr, 2000; Dawling et al., 2003). The results in estradiol accumulation accordance with 17β-HSD1 recovered to nearly the same level as the normals (Fig. 4). The expressions of CYP19 and 17β-HSD3 were not significantly modified by all treatments (Fig. 5).
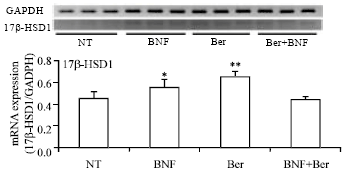 | |
| Fig. 4: | Expression pattern of testicular 17β-HSD1 mRNA in male mice, Seven week-old mice were orally fed daily with 7.5 mg/kg/day of berberine (Ber) for 7 days and/or intraperitoneally given 30 mg/kg/day of β-naphthoflavone (BNF) in the last 3 days. The control group (NT) was orally given corn oil daily for 7 days. The relative mRNA expression levels were normalized by that of GAPDH. The data are presented as the Mean±SD (n = 3-4) from at least 2 independent experiments. A significant difference was examined by ANOVA with Tukey post hoc test. * p<0.05, ** p<0.001 compared to NT group; # p<0.05 compared to BNF group |
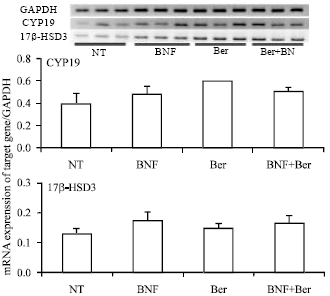 | |
| Fig. 5: | Expression pattern of testicular CYP19 and 17β-HSD3 mRNA in male mice, Seven week-old mice were orally fed daily with 7.5 mg/kg/day day of berberine (Ber) for 7 days and/or intraperitoneally given 30 mg/kg/day day of β-naphthoflavone (BNF) in the last 3 days. The control group (NT) was orally given corn oil daily for 7 days. The relative mRNA expression levels were normalized by that of GAPDH. The data are presented as the Mean±SD (n = 3-4) from at least 2 independent experiments. A significant difference was examined by ANOVA with Tukey post hoc test. *p<0.05, **p<0.001 compared to NT group;#p<0.05 compared to BNF group |
CONCLUSION
In the conclusion, the single treatment of berberine up-regulated the expression of testicular CYP17, 3β-HSD and 17β-HSD1 mRNA. The single treatment of BNF up-regulated only 17β-HSD1 mRNA expression. While the combination of berberine and BNF up-regulated in expression of CYP17 and 3β-HSD. These observations suggested that berberine disturbed sex hormone synthesis pathway; consequently possibly resulted in modification of circulating estrogen pattern in the body. Therefore, a caution should be noted for the use of berberine as an alternative medicine. However, a further study to affirm the evidence of sex hormone modification by berberine in illness is still required.
REFERENCES
- Bates, K.A., A.R. Harvey, M. Carruthers and R.N. Martins, 2005. Alzheimer's disease: The impact of age-related changes in reproductive hormones. Cell. Mol. Life Sci., 62: 281-292.
CrossRefDirect Link - Bergman-Jungestrom, M., M. Gentile, A.C. Lundin, South-East Breast Cancer Group and S. Wingren, 1999. Association between cyp17 gene polymorphism and risk of breast cancer in young women. Int. J. Cancer, 84: 350-353.
CrossRefDirect Link - Carani, C., K. Qin, M. Simoni, M. Faustini-Fustini and S. Serpente et al., 1997. Effect of testosterone and estradiol in a man with aromatase deficiency. N. Engl. J. Med., 337: 91-95.
CrossRefDirect Link - Chatuphonprasert, W., K. Jarukamjorn, S. Kondo and N. Nemoto, 2009. Synergistic increases of metabolism and oxidation-reduction genes on their expression after combined treatment with a CYP1A inducer and andrographolide. Chem. Biol. Interact., 182: 233-238.
CrossRef - Chatuphonprasert, W., T. Sangkawat, N. Nemoto and K. Jarukamjorn, 2011. Suppression of beta-naphthoflavone induced CYP1A expression and lipid-peroxidation by berberine. Fitoterapia, 82: 889-895.
CrossRefDirect Link - Dawling, S., N. Roodi and F.F. Parl, 2003. Methoxyestrogens exert feedback inhibition on cytochrome P450 1A1 and 1B1. Cancer Res., 63: 3127-3132.
PubMedDirect Link - Degawa, M., K. Hanaki and M. Sekimoto, 2006. A hepatocarcinogenic tryptophan-pyrolyzate component, Trp-P-1, decreases serum total testosterone level and induces hepatic Cyp1a2 in male mice. Cancer Sci., 97: 32-37.
CrossRefPubMedDirect Link - Kuo, C.L., C.W. Chi and T.Y. Liu, 2004. The anti-inflammatory potential of berberine in vitro and in vivo. Cancer Lett., 203: 127-137.
CrossRefPubMedDirect Link - Letasiova, S., S. Jantova, L. Cipak and M. Muckova, 2006. Berberine-antiproliferative activity in vitro and induction of apoptosis/necrosis of the U937 and B16 cells. Cancer Lett., 239: 254-262.
CrossRefPubMedDirect Link - Liehr, J.G., 2000. Is estradiol a genotoxic mutagenic carcinogen? Endocr. Rev., 21: 40-54.
CrossRefPubMedDirect Link - Miller, W.L., 1988. Molecular biology of steroid hormone synthesis. Endocr. Rev., 9: 295-318.
CrossRefPubMedDirect Link - Morishima, A., M.M. Grumbach, E.R. Simpson, C. Fisher and K. Qin, 1995. Aromatase deficiency in male and female siblings caused by a novel mutation and the physiological role of estrogens. J. Clin. Endocrinol. Metab., 80: 3689-3698.
CrossRefPubMedDirect Link - Rojsanga, P., W. Gritsanapan and L. Suntornsuk, 2006. . Determination of berberine content in the stem extracts of Coscinium fenestratumby TLC densitometry. Med. Princ. Pract., 15: 373-378.
Direct Link - Sato, F. and Y. Yamada, 1984. High berberine producing cultures of Coptis japonica. Phytochemistry, 23: 281-285.
CrossRef - Schmeller, T., B. Latz-Bruning and M. Wink, 1997. Biochemical activities of berberine, palmatine and sanguinarine mediating chemical defence against microorganisms and herbivores. Phytochemistry, 44: 257-266.
CrossRefPubMedDirect Link - Sha, J., P. Baker and P.J. O'Shaughnessy, 1996. Both reductive forms of 17β-Hydroxysteroid dehydrogenase (types 1 and 3) are expressed during development in the mouse testis. Biochem. Biophys. Res. Commun., 222: 90-94.
CrossRefPubMedDirect Link - Stocco, D.M., 2000. Intramitochondrial cholesterol transfer. Biochimica Biophysica Acta (BBA)-Mol. Cell Biol. Lipids, 1486: 184-197.
CrossRefPubMedDirect Link - Udomsuk, L., T. Juengwatanatrakul, W. Putalun and K. Jarukamjorn, 2011. Down regulation of gene related sex hormone synthesis pathway in mouse testes by miroestrol and deoxymiroestrol. Fitoterapia, 82: 1185-1189.
CrossRefPubMedDirect Link - Wickenheisser, J.K., V.L. Nelson-DeGrave, K.L. Hendricks, R.S. Legro, J.F. Strauss and J.M. McAllister, 2005. Retinoids and retinol differentially regulate steroid biosynthesis in ovarian theca cells isolated from normal cycling women and women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab., 90: 4858-4865.
CrossRefPubMedDirect Link - Wu, H.L., C.Y. Hsu, W.H. Liu and B.Y.M. Yung, 1999. Berberine-induced apoptosis of human leukemia HL-60 cells is associated with down-regulation of nucleophosmin/B23 and telomerase activity. Int. J. Cancer, 81: 923-939.
CrossRefDirect Link - Yanase, T., 1995. 17α-Hydroxylase/17,20-lyase defects. J. Steroid Biochem. Mol. Biol., 53: 153-157.
CrossRefPubMedDirect Link - Yanase, T., E.R. Simpson, M.R. Waterman, 1991. 17α-hydroxylase/17,20-lyase deficiency: From clinical investigation to molecular definition. Endocr. Rev., 12: 91-108.
CrossRefPubMedDirect Link - Zhang, Q., X. Xiao, K. Feng, T. Wang and W. Li et al., 2011. Berberine moderates glucose and lipid metabolism through multi pathway mechanism. Evid. Based Complement. Alternat. Med., Vol. 2011.
CrossRefDirect Link - Zhang, Y., X. Li, D. Zou, W. Liu and J. Yang et al., 2008. Treatment of type 2 diabetes and dyslipidemia with the natural plant alkaloid berberine. J. Clin. Endocrinol. Metab., 93: 2559-2565.
CrossRefPubMedDirect Link - Zuber, M.X., E.R. Simpson and M.R. Waterman, 1986. Expression of bovine 17α-hydroxylase cytochrome P450 cDNA in nonsteroidogenic (Cos 1) cells. Science, 234: 1258-1261.
CrossRefPubMedDirect Link