
Pawitra Pulbutr
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy Mahasarakham University, Mahasarakham, 44150, Thailand
Klangnapa Thunchomnang
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy Mahasarakham University, Mahasarakham, 44150, Thailand
Karakate Lawa
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy Mahasarakham University, Mahasarakham, 44150, Thailand
Apiwat Mangkhalathon
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy Mahasarakham University, Mahasarakham, 44150, Thailand
Pattarapong Saenubol
Pharmaceutical Chemistry and Natural Product Research Unit, Faculty of Pharmacy Mahasarakham University, Mahasarakham, 44150, Thailand
International Journal of Pharmacology
Year: 2011 | Volume: 7 | Issue: 5 | Page No.: 629-634







ABSTRACT
Zingerone is a non-volatile pungent compound mostly found in ginger which is a herbal medicine used for various purposes. The anti-obesity actions of ginger and zingerone have also been documented. This study was aimed to investigate the effects of zingerone on adipocyte lipolysis in adipocytes isolated from Normal Pellet Diet (NPD)-fed rats and High Fat Diet (HFD)-fed rats. Adipocyte suspensions were prepared from the epididymal fat pads of male Wistar rats by using the collagenase digestion method. The concentrations of Free Fatty Acid (FFA) in the incubation media were used as an index of adipocyte lipolysis. Zingerone at the concentration of 1,000 μM significantly increased both basal lipolysis and isoprenaline (0.l μM)-induced lipolysis in adipocytes derived from the NPD-fed rats with the FFA concentrations of 682.92±37.00 μM mL-1 Packed Cell Volume (PCV)/ h and 1,361.51±62.90 μM mL-1 PCV/ h, respectively (p<0.05; n = 4). In adipocytes derived from the HFD-fed rats, zingerone at any concentration tested had no effect on the basal lipolysis. Zingerone only at the concentration of 100 μM significantly stimulated the isoprenaline-induced lipolysis with the FFA concentration of 1,383.88±36.73 μM mL-1 PCV/ h (p<0.05; n = 4) in adipocytes derived from the HFD-fed rats. In conclusion, zingerone appeared to produce less lipolytic actions in adipocytes derived from the HFD-fed rats. The reason for the lower lipolytic response to zingerone in HFD-derived adipocytes should be investigated further, especially in the molecular aspects.
PDF Abstract XML References Citation
Received: March 31, 2011;
Accepted: May 30, 2011;
Published: July 23, 2011
How to cite this article
Pawitra Pulbutr, Klangnapa Thunchomnang, Karakate Lawa, Apiwat Mangkhalathon and Pattarapong Saenubol, 2011. Lipolytic Effects of Zingerone in Adipocytes Isolated from Normal Diet-Fed Rats and High Fat Diet-Fed Rats. International Journal of Pharmacology, 7: 629-634.
DOI: 10.3923/ijp.2011.629.634
URL: https://scialert.net/abstract/?doi=ijp.2011.629.634
DOI: 10.3923/ijp.2011.629.634
URL: https://scialert.net/abstract/?doi=ijp.2011.629.634
INTRODUCTION
Obesity which is a consequence of an imbalance between energy intake and energy expenditure, is an important risk factor for several diseases such as hypertension, dyslipidemia, type 2 diabetes and several types of cancers (Kopelman, 2000). Over accumulation of adipose tissue in the body also contributes to the pathogenesis of insulin resistance which is implicated in several chronic metabolic and cardiovascular disorders (Samuel et al., 2010). The prevalence of obesity is continuously increasing globally. Sedentary lifestyle and over food consumption, especially a high amount of fat intake, are the major causes of obesity (Biro and Wien, 2010). Adipocyte lipolysis, which is characterized by a breakdown of stored triglyceride in adipocytes, plays a crucial role in the regulation of fat mass (Arner, 2005). Thus, modulation of adipocyte lipolysis is one means to reduce fat mass and ameliorate obesity.
Ginger (rhizome of Zingiber officinale Roscoe) has been used widely as a condiment in oriental foods and beverages. It has also been known as a herbal medicine for the treatment of various disorders such as common cold, gastrointestinal disorders, motion sickness, diabetes (Karim et al., 2011), metabolic disorder and also obesity (Westerterp-Plantenga et al., 2006; Kundu et al., 2009). The anti-obesity actions of ginger extracts have been documented in chemical- and high fat diet-induced obese mice (Goyal and Kadnur, 2006; Han et al., 2005). However, the biologically active components in ginger producing such anti-obesity actions are still not clearly established. Zingerone is one of the non-volatile pungent compounds mostly found in ginger. It was reported that zingerone increased catecholamine secretion from the adrenal medulla (Kawada et al., 1988). Thus, it is suggested that the favorable effect of zingerone on adiposity is partly due to its catecholamine-releasing action. Zingerone was found to inhibit obesity-induced inflammatory responses by suppressing adipose tissue macrophage accumulation and activation and inhibiting release of monocyte chemoattractant protein-1 (MCP-1) from adipocytes (Woo et al., 2007). Additionally, it was evidenced that zingerone produced its anti-obestiy action in ovariectomised rats possibly through an activation of adipocytes lipolysis (Han et al., 2008). It is currently unknown whether zingerone produce the lipolytic stimulating action in adipocytes derived from animals fed with high fat diet which is an established model of diet-induced obesity. This study was aimed to investigate the effects of zingerone on adipocyte lipolysis in isolated adipocytes derived from high fat diet-fed rats and to compare its actions to those found in adipocytes derived from normal diet-fed rats.
MATERIALS AND METHODS
Preparation of adipocytes: Sixteen male Wistar rats weighing 140-160 g were obtained from the National Laboratory Animal Center, Mahidol University, Thailand. The animals were kept at constant temperature (25±1°C) with a 12 h dark-light cycle. After acclimatization period of one week, the rats were randomly divided into two groups. The rats in the first group (n = 8) were fed with Normal Pellet Diet (NPD) (C.P. mice feed food No. 082, Bangkok, Thailand) (NPD-group) whilst the rats in the second group (n = 8) were fed with high fat diet (HFD-group). The fat content in NPD and HFD are approximately 12 and 60% of total energy, respectively. High fat diet was prepared according to the formulation of Srinivasan et al. (2005) with slight modification. The animals were fed ad libitum and freely accessed to water. After three weeks, the rats were sacrificed by CO2 overdose and epididymal fat pads were dissected for preparation of adipocyte suspension. All procedures performed with the animals were approved by the animal research ethic committee, Mahasarakham University, Thailand.
Adipocytes were isolated according to the method of Rodbell (1964) with some modifications. Briefly, the epididymal fat pads were rinsed with Hanks buffer containing 5 mM glucose and 0.5 % bovine serum albumin (Sigma) (BSA, with less than 0.005% (w/w) free fatty acid). Hanks physiological solution containing (in mM): 5.6 KCl, 138 NaCl, 4.2 NaHCO3, 1.2 NaH2PO4, 2.6 CaCl2, 1.2 MgCl2, 10 HEPES, pH 7.4 (with NaOH). The tissues were cut into small pieces and then digested with 0.5 mg mL-1 collagenase type II enzyme at 37°C. Isolated adipocytes were filtered through nylon mesh and then rinsed twice with collagenase free Hanks buffer. After the second wash, the adipocytes were resuspended with Hanks solution into 1:1 (v/v) proportion to give a final volume of the cell suspension at about 8-15 mL, depending on the volume of adipocytes. The cell suspension was transferred to a fresh Nalgene conical flask and stored in the water bath at 37°C before performing experiments. All experiments were conducted at the Faculty of Pharmacy, Mahasarakham University, Thailand during 1st December 2010 to 14th February 2011.
Adipocyte lipolysis: Adipocytes were incubated for 60 min in microfuge tubes at 37°C with Hanks buffer, pH 7.4, containing 5 mM glucose, 0.5% BSA and various concentrations of zingerone (Sigma) at 10 μM, 100 μM and 1,000 μM. For stimulated lipolysis study, the experiments were performed in the presence of isoprenaline (Sigma) at the concentration of 0.1 μM. The final volume of the incubations was adjusted to 1 mL. After 1 h incubation, the cell free incubation media were collected and the concentrations of Free Fatty Acid (FFA) were measured by using non-esterified fatty acid assay kit from WAKO Chemicals (Japan).
Statistical analysis: The results are expressed as mean±SEM. Statistical analysis was performed by using one-way analysis of variance (ANOVA) followed by Bonferroni post hoc test. Statistical tests were performed using SPSS software version 16.0 (SPSS Inc., Chicago, Illinois, USA). The data was considered as significantly different when p-value <0.05.
RESULTS
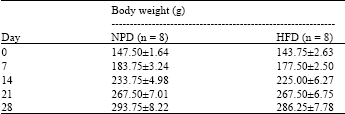
Body weight and epididymal fat pad weight: The body weight of the rats was recorded at the beginning of the experiment (day 0) and every week thereafter (day 7, 14, 21 and 28). There was no difference in the body weight of the rats fed with NPD or HFD (Table 1). However, the epididymal fat weight of HFD-fed rats (2.18±0.18/100 g of body weight, n =8) was significantly higher than that of NPD-fed control rats (1.43±0.11/100 g of body weight n = 8) with the p-value of <0.05.
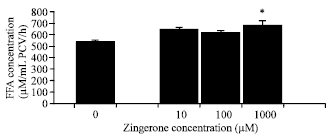
Effects of zingerone in adipocytes derived from the NPD-fed rats (NPD-group): The FFA concentration in the basal condition without adding any lipolytic-stimulating agents was 535.32±14.79 μM mL-1 Packed Cell Volume (PCV)/ h (Fig. 1). Zingerone at the concentration of 1,000 μM significantly increased adipocyte lipolysis in the NPD-group with the FFA concentration of 682.92±37.00 μM mL-1 PCV/ h (p<0.05) (Fig. 1).
| Table 1: | Body weight of NPD-fed rats and HFD-fed rats (Mean±SEM) |
 | |
 | |
| Fig. 1: | Effects of zingerone on basal adipocyte lipolysis in the NPD-group. The results are expressed as the Mean±SEM (n = 4). *p<0.05 when compared with basal lipolysis (One-way ANOVA) |
 | |
| Fig. 2: | Effects of zingerone on isoprenaline-induced lipolysis in the NPD-group. The results are expressed as the Mean±SEM (n = 4). Adipocytes were treated with 0.1 μM isoprenaline under the condition shown. *p<0.05 when compared with isoprenaline-induced lipolysis (One-way ANOVA) |
Zingerone at the lower concentrations used (10 μM and 100 μM) caused a slight increase in the basal lipolysis, with the FFA concentrations of 643.51±18.88 and 614.96±17.21 μM mL-1 PCV/h, respectively (Fig. 1). However, these FFA levels were not significantly different from that of the basal lipolysis.
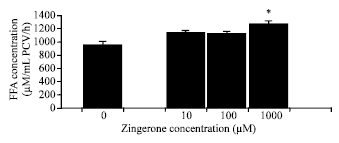
Isoprenaline (0.1 μM) caused a significant lipolytic stimulation in adipocytes derived from the NPD-group, with the FFA concentration of 1,026.18±60.85 μM mL-1 PCV/ h (p<0.05) (Fig. 2). The FFA level in the presence of isoprenaline was approximately 191% of that in the absence of isoprenaline (basal lipolysis). Zingerone at the highest concentration used (1,000 μM) significantly increased the isoprenaline-induced lipolysis (FFA concentration of 1,361.51±62.90 μM mL-1 PCV/ h (p<0.05). When zingerone at the concentrations of 10 μM and 100 μM was added, the isoprenaline-induced lipolysis was slightly increased (FFA concentrations of 1,222.91±40.46 and 1,208.00±42.83 μM mL-1 PCV/ h, respectively). However, these FFA levels did not reach a significant difference when compared to that in the presence of isoprenaline alone (Fig. 2).
 | |
| Fig. 3: | Effects of zingerone on basal adipocyte lipolysis in the HFD-group. The results are expressed as the Mean±SEM (n = 4) |
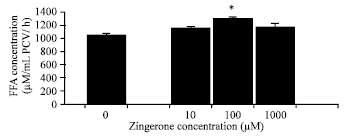 | |
| Fig. 4: | Effects of zingerone on isoprenaline-induced lipolysis in the HFD-group. The results are expressed as the Mean±SEM (n = 4). Adipocytes were treated with 0.1 μM isoprenaline under the condition shown. *p<0.05 when compared with isoprenaline-induced lipolysis (One-way ANOVA) |
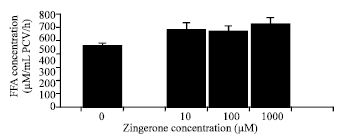
Effects of zingerone in adipocytes derived from the HFD-fed rats (HFD-group): The FFA concentration in the basal lipolytic condition of the HFD-fed group was 562.75±23.92 μM mL-1 PCV/ h. This was not different from the basal lipolytic level in the NPD-group described above. Zingerone at any concentration tested did not significantly increase the basal lipolysis in adipocytes derived from the HFD-fed rats. The FFA concentrations in the presence of zingerone at 10 μM, 100 μM and 1,000 μM were 710.02±63.60, 694.32±49.01 and 762.35±57.47 μM mL-1 PCV/ h, respectively (Fig. 3).
Isoprenaline at the concentration of 0.1 μM significantly increased adipocyte lipolysis in the HFD-group with the FFA concentration of 1,119.88±25.62 μM mL-1 PCV/ h (p<0.05) (Fig. 4). The FFA level in the condition of isoprenaline-induced lipolysis was approximately 199% of the basal lipolysis in adipocytes derived from the HFD-fed rats. The lipolytic action of isoprenaline in the HFD-group was not different from that in the NPD-group described above. Zingerone only at the concentration of 100 μM significantly stimulated the isoprenaline-induced lipolysis in adipocytes derived from the HFD-fed rats, with the FFA concentration of 1,383.88±36.73 μM mL-1 PCV/ h (p<0.05) (Fig. 4). However, at the concentrations of 10 and 1,000 μM, zingerone did not produce a significant increase in the isoprenaline-induced lipolysis in adipocytes derived from the HFD-group (Fig. 4). The FFA concentrations in the presence of isoprenaline and zingerone (10 μM and 1,000 μM) were 1,234.06±20.88 and 1,246.96±70.36 μM mL-1 PCV/ h, respectively (Fig. 4).
DISCUSSION
There was no difference in the body weight of the rats fed with two different types of diet (Table 1). Harris (1991) also showed that after 3 weeks of feeding, the body weight of the rats fed with HFD was not different from that of the rats fed with NPD. Conversely, the weight of epididymal fat pads in the HFD-fed rats was significantly higher than that in the NPD-fed rats. This is in agreement with the results of Naim et al. (1985) which reported the higher amount of epididymal fat pads in the rats fed with HFD. Additionally, Collin et al. (2006) also showed that after 3 weeks, the rats fed with HFD had a significantly higher value in the visceral fat pad weight. From their study, there was also no statistical difference in the body weight between the NPD-fed rats and the HFD-fed rats after 3 weeks.
Zingerone only at the concentration of 1,000 μM significantly increased the basal lipolysis in adipocytes derived from the NPD-fed rats (Fig. 1). This finding is in accordance with the results from the study of Han et al. (2008) which reported that a significant increase in the basal lipolysis was found only when 1,000 μM zingerone was used. From their study, zingerone at the lower concentrations tested (1 μM, 10 μM and 100 μM) slightly increased the basal lipolysis but these were not significantly different. These are similar to the results presented here in this study.
In adipocytes derived from the NPD-fed rats, the FFA level in the presence of isoprenaline (0.1 μM) was approximately 191% of the basal lipolysis. This was not dissimilar from the results reported by Zhang et al. (2009), in which 0.1 μM isoprenaline stimulated lipolysis with the FFA level of approximately 200% of the basal lipolysis. Zingerone only at the concentration of 1,000 μM caused a significant increase in the isoprenaline-induced lipolysis (Fig. 2). The FFA level in the presence of isoprenaline and 1,000 μM zingerone was approximately 132% of the isoprenaline-induced lipolysis. From the study of (Han et al., 2008), zingerone at 1,000 μM significantly increased norepinephrine (1 μg mL-1)-induced lipolysis, with the FFA level of approximately 1,600% of norepinephrine-induced lipolysis. It was reported from their studies that zingerone at the concentrations of 10 and 100 μM also significantly increased norepinephrine-induced lipolysis, with the FFA levels of approximately 200 and 400% of norepinephrine-induced lipolysis, respectively (Han et al., 2008). However, these lower concentrations of zingerone did not cause an increase in the isoprenaline-induced lipolysis in the current study. Han et al. (2008) also showed that the lipolytic stimulating action of zingerone was associated with the translocation of hormone-sensitive lipase enzyme from the cytosol into the lipid droplets in adipocytes. The difference in the lipolytic response of zingerone (in term of percentage of the lipolytic stimulating agent-induced lipolysis) between the studies may be due to the difference in lipolytic stimulating agents used. Norepinephrine is a non-selective α- and β-adrenoceptor agonist whilst isoprenaline is a selective β-adrenoceptor agonist (Griffith, 2008). The EC50s of norepinephrine and isoprenaline for their lipolytic stimulating activity are 1.04x10¯7 and 3.46x10¯8 M, respectively (Farias-Silva et al., 1999). Thus, the lipolytic stimulating potency of isoprenaline is higher than that of norepinephrine. The difference in the levels of lipolytic stimulation induced by isoprenaline and norepinephrine may partly be responsible for the dissimilar actions of zingerone between studies.
The level of basal lipolysis in adipocytes derived from the NPD-fed rats was similar to that in adipocytes derived from the HFD-fed rats. This is in accordance to the results reported by Garcia-Diaz et al. (2009) in which there was no difference in the basal lipolytic responses between adipocytes isolated from the HFD-group and the NPD-group. This is also similar to the results reported earlier by our group (Pulbutr and Rattanakiat, 2010). However, this is contrary to the results from the study of Collin et al. (2006) which reported that after 3 week-feeding, the HFD-derived adipocytes had a higher level of the basal lipolysis compared to that of the NPD-derived adipocytes.
The level of isoprenaline-induced lipolysis in adipocytes derived from the NPD was not different from that in the HFD-fed adipocytes. This is similar to the results of Collin et al. (2006) which showed that after 3 weeks, isoprenaline (0.1 μM)-induced lipolysis was similar between the NPD-group and the HFD-group. However, Portillo et al. (1999) reported the lower lipolytic response to dobutamine (a selective β1-agonist) and BRL37344 (a selective β3-agonist) in adipocytes derived from the HFD-fed rats. The investigators suggested that this may be due to a loss of β1 and β3-adrenoceptors in HFD-derived adipocytes (Portillo et al., 1999). Since isoprenaline can activate all three types of β-adrenoceptor (β1-, β2-, β3-), thus its lipolytic stimulating efficacy is possibly not affected by HFD-feeding.
In adipocytes derived from the rats fed with HFD, the lipolytic response of zingerone was different from that in adipocytes derived from the rats fed with NPD. Zingerone at any concentration tested could not produce a significant increase in the basal lipolysis in the HFD-group (Fig. 3). Additionally, only zingerone at the concentration of 100 μM significantly stimulated the isoprenaline-induced lipolysis in adipocytes derived from the HFD-fed rats (Fig. 4). Collectively, the lipolytic actions of zingerone are likely to be less effective in adipocytes derived from the HFD-fed rats. The effects of HFD on some key components involving in adipocyte lipolysis such as perilipin, hormone-sensitive lipase enzyme, adipocyte triglyceride lipase enzyme, have been documented (Collin et al., 2006; Gaidhu et al., 2010). The alterations of these proteins may partly be the reason for the dissimilar responses to zingerone in adipocytes derived from the different diet-fed rats.
Interestingly, Zhang et al. (2007) reported the depletion of both protein and mRNA expressions of transient receptor potential vanilloid type 1 (TRPV1) in visceral adipose tissue from mice on high fat diet. TRPV1 is a non-selective cation channel presented in various tissues including visceral adipose tissue (Zhang et al., 2007). TRPV1 has been found to be implicated in adipocyte metabolism including adipogenesis and adipocyte lipolysis (Zhang et al., 2007; Motter and Ahern, 2008). Zingerone exerted an agonistic action on TRPV1 with the EC50s of 7.41x10¯5 M and 5.75x10¯4 M in cells expressing rat and human TRPV1, respectively (Witte et al., 2002). The concentrations used in this study are also in the range of TRPV1 activation. If zingerone caused a lipolytic stimulation via the activation of TRPV1, the depletion of TRPV1 expression is thus likely to be involved in the lower lipolytic actions of zingerone in adipocytes derived from the HFD-fed rats. Further experiments are thus needed to prove this speculation.
CONCLUSION
Zingerone at the concentration of 1,000 μM produced a significant increase in both basal lipolysis and isoprenaline-induced lipolysis in adipocytes derived from the NPD-fed rats. Generally, adipocytes derived from the HFD-fed rats are less sensitive to the lipolytic stimulating action of zingerone. Zingerone at any concentration tested did not produce a significant change in the basal lipolysis. The isoprenaline-induced lipolysis was stimulated by zingerone only at the concentration of 100 μM in the HFD-group. Further experiments should be performed to explore the lower responses to zingerone in the HFD-fed derived adipocytes, especially in the molecular aspects.
ACKNOWLEDGMENTS
We thank the Faculty of Pharmacy, Mahasarakham University for the laboratory support. We would like to express our gratitude to Assistant Professor Songsak Jumpawadee, Faculty of Veterinary Medicine and Animal Sciences, Mahasarakham University for preparation of animal diet. We thank Ajarn Thidarat Somdee, Faculty of Public Health, Mahasarakham University for her kind technical support. We are also grateful to Dr. Benjamart Cushnie and Dr. Sakulrat Rattanakiat for their useful comments on the project.
REFERENCES
- Arner, P., 2005. Human fat cell lipolysis: Biochemistry, regulation and clinical role. Best Pract. Res. Clin. Endocrinol. Metab., 19: 471-482.
Direct Link - Biro, F.M. and M. Wien, 2010. Childhood obesity and adult morbidities. Am. J. Clin. Nutr., 91: 1499S-1505S.
CrossRef - Collin, P., N. Chapados, E. Dufresne, P. Corriveau, P. Imbeault and J.M. Lavoie, 2006. Time course of changes in in vitro lipolysis of intra-abdominal fat depots in relation to high-fat diet-induced hepatic steatosis in rats. Br. J. Nutr., 96: 268-275.
PubMed - Gaidhu, M.P., N.M. Anthony, P. Patel, T.J. Hawke and R.B. Ceddia, 2010. Dysregulation of lipolysis and lipid metabolism in visceral and subcutaneous adipocytes by high-fat diet: Role of ATGL, HSL and AMPK. Am. J. Physiol. Cell Physiol., 298: C961-C971.
CrossRef - Garcia-Diaz, D.F., J. Campion, F.I. Milagro, L. Paternain, A. Solomon and J.A. Martinez, 2009. Ascorbic acid oral treatment modifies lipolytic response and behavioural activity but not glucocorticoid metabolism in cafeteria diet-fed rats. Acta. Physiol. (Oxf)., 195: 449-457.
CrossRef - Goyal, R.K. and S.V. Kadnur, 2006. Beneficial effects of Zingiber officinale on goldthioglucose induced obesity. Fitoterapia. J., 77: 160-163.
PubMedDirect Link - Han, L.K., C. Morimoto, Y.N. Zheng, W. Li, E. Asami, H. Okuda and M. Saito, 2008. Effects of zingerone on fat storage in ovariectomized rats. Yakugaku Zasshi, 128: 1195-1201.
PubMed - Han, L.K., X.J. Gong, S. Kawano, M. Saito, Y. Kimura and H. Okuda, 2005. Antiobesity actions of Zingiber officinale Roscoe. Yakugaku Zasshi, 125: 213-217.
Direct Link - Harris, R.B., 1991. Growth measurements in Sprague-Dawley rats fed diets of very low fat concentration. J. Nutr., 121: 1075-1080.
PubMed - Kawada, T., S. Sakabe, T. Watanabe, M. Yamamoto and K. Iwai, 1988. Some pungent principles of spices cause the adrenal medulla to secrete catecholamine in anesthetized rats. Proc. Soc. Exp. Biol. Med., 188: 229-233.
PubMed - Kundu, J.K., H.K. Na and Y.J. Surh, 2009. Ginger-derived phenolic substances with cancer preventive and therapeutic potential. Forum Nutr., 61: 182-192.
CrossRefPubMedDirect Link - Motter, A.L. and G.P. Ahern, 2008. TRPV1-null mice are protected from diet-induced obesity. FEBS Lett., 582: 2257-2262.
PubMed - Naim, M., J.G. Brand, M.R. Kare and R.G. Carpenter, 1985. Energy intake, weight gain and fat deposition in rats fed flavored, nutritionally controlled diets in a multichoice (cafeteria) design. J. Nutr., 115: 1447-1458.
CrossRefPubMedDirect Link - Portillo, M.P., E. Simon, M.A. Garcia-Calonge and A.S. Del Barrio, 1999. Effect of high-fat diet on lypolisis in isolated adipocytes from visceral and subcutaneous WAT. Eur. J. Nutr., 38: 177-182.
PubMed - Pulbutr, P. and S. Rattanakiat, 2010. Lipolytic effects of genistein and daidzein in adipocytes derived from normal diet-fed rats and high fat diet-fed rats. J. Biol. Sci., 10: 754-760.
CrossRef - Rodbell, M., 1964. Metabolism of isolated fat cells. I. Effects of hormones on glucose metabolism and lipolysis. J. Biol. Chem., 239: 375-380.
PubMed - Samuel, V.T., K.F. Petersen and G.I. Shulman, 2010. Lipid-induced insulin resistance: Unravelling the mechanism. Lancet, 375: 2267-2277.
CrossRefDirect Link - Srinivasan, K., B. Viswanad, L. Asrat, C.L. Kaul and P. Ramarao, 2005. Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: A model for type 2 diabetes and pharmacological screening. Pharmacol. Res., 52: 313-320.
CrossRefDirect Link - Westerterp-Plantenga, M., K. Diepvens, A.M. Joosen, S. Berube-Parent and A. Tremblay, 2006. Metabolic effects of spices, teas and caffeine. Physiol. Behav., 89: 85-91.
PubMed - Witte, D.G., S.C. Cassar, J.N. Masters, T. Esbenshade and A.A. Hancock, 2002. Use of a fluorescent imaging plate reader-based calcium assay to assess pharmacological differences between the human and rat vanilloid receptor. J. Biomol. Screen., 7: 466-475.
PubMed - Woo, H.M., J.H. Kang, T. Kawada, H. Yoo, M.K. Sung and R. Yu, 2007. Active spice-derived components can inhibit inflammatory responses of adipose tissue in obesity by suppressing inflammatory actions of macrophages and release of monocyte chemoattractant protein-1 from adipocytes. Life Sci., 80: 926-931.
PubMed - Zhang, L.L., D.Y. Liu, L.Q. Ma, Z.D. Luo and T.B. Cao et al., 2007. Activation of transient receptor potential vanilloid type-1 channel prevents adipogenesis and obesity. Circ. Res., 100: 1063-1070.
PubMed - Zhang, T., J. He, C. Xu, L. Zu and H. Jiang et al., 2009. Mechanisms of metformin inhibiting lipolytic response to isoproterenol in primary rat adipocytes. J. Mol. Endocrinol., 42: 57-66.
PubMed - Karim, A., M.N. Sohail, S. Munir and S. Sattar, 2011. Pharmacology and phytochemistry of Pakistani herbs and herbal drugs used for treatment of diabetes. Int. J. Pharmacol., 7: 419-439.
CrossRef