
Saeideh Momtaz
Department of Plant Science, Faculty of Natural and Agricultural Science, University of Pretoria, Pretoria, South Africa
Mohammad Abdollahi
Pharmaceutical Sciences Research Center, Tehran University of Medical Sciences, Tehran, Iran
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 1 | Page No.: 1-19







ABSTRACT
Aging is an undeniable fact shadowing animal life but while it comes to human, aging affects all aspects of their physical and emotional abilities. Aging occurs by multi-stimuli which is affected by both environmental and internal conditions. During the past decade, remarkable recognition of cellular senescence and its related pathways has happened. The increased production and/or ineffective scavenging of oxidants in particular, Reactive Oxygen Species (ROS) play a critical role in cellular senescence. High reactivity of ROS determines chemical changes in virtually all cellular components, leading to senescence mostly via DNA damage. Present paper discusses the oxidative stress as the main cause of aging and the benefit of antioxidants. So far, more effective explorations are needed to decrease the deleterious effects of oxidative-stress-induced aging. This approach could be achieved through manipulations of antioxidant enzyme systems, development of drug therapies, manipulation of aging related genes and genetic manipulation of plant-based antioxidant resources.
PDF Abstract XML References Citation
Received: August 10, 2011;
Accepted: November 08, 2011;
Published: December 23, 2011
How to cite this article
Saeideh Momtaz and Mohammad Abdollahi, 2012. A Comprehensive Review of Biochemical and Molecular Evidences from Animal and Human Studies on the Role of Oxidative Stress in Aging: An Epiphenomenon or the Cause. Asian Journal of Animal and Veterinary Advances, 7: 1-19.
DOI: 10.3923/ajava.2012.1.19
URL: https://scialert.net/abstract/?doi=ajava.2012.1.19
DOI: 10.3923/ajava.2012.1.19
URL: https://scialert.net/abstract/?doi=ajava.2012.1.19
INTRODUCTION
Aging is an undeniable fact shadowing animal life but while it comes to human, aging affects all aspects of their physical and emotional abilities. To compromise with aging-induced deteriorations, humans always were thoughtful to find the solutions that either prevent or cure various symptoms of aging. Various natural or synthetic agents, hormones, enzymes, genetical and physiological ways were examined to approach this goal. Amongst natural resources, plants in particular have shown very optimistic effects as anti-aging. Present study discusses the oxidative stress as the main cause of aging and the benefit of antioxidants.
WHAT IS THE REAL CONCEPT OF AGING?
In general, aging refers to ‘an accumulation of changes in a cell, tissue or an organ over time which leads to lose its routine function and vitality, undergoing mortality’ (Atwood et al., 2005). Masoro (1995) delightfully expressed aging as: deteriorative changes with time during postmaturational life that underlie an increasing vulnerability to challenges, thereby decreasing the ability of an organism to survive. Causes of this deterioration may be found in; accumulation of genetic damage or mutations in genes; the deposition of lipofuscin and advanced glycation endproducts in many cell and tissue locations; the abnormal modifications of proteins and the accumulation of insoluble aggregates; damage by ROS in many contexts; loss of immune functions and autoimmunity; osteoporosis and osteoarthritis; inflammatory damage to tissues; hormone imbalance; the loss or gain of DNA methylation and finally incidence of tumors (Holliday, 2006). Despite the public perception, aging is not a universal phenomenon though plenty studies mentioned some plants or animals with negligible senescence.
MOLECULAR BIOLOGY OF SENESCENCE
The word of ‘senescence’ is derived from the Latin word ‘senex’, meaning old age or advanced age. In biology, senescence states deteriorative processes of aging that follow development and maturation. Senescence has engaged human thoughts along history, nevertheless, it is well understood nowadays that multi-stimuli lead to senescence, of which changes in chromosome structures and functions were recognized as roots of this progress. Senescence could be defined for an individual, an organ, a tissue and lastly a cell.
Senescence may impact on aging through two mechanisms; accumulation of senescent cells in tissues may reach a point that compromises functionality and senescence may also limit the regenerative potential of adult stem cells (a limitation that may be produced as well by quiescence or apoptosis of stem cells) (Collado et al., 2007). Leonard Hayflick and Paul Moorhead in 1961 discovered that human fibroblasts derived from embryonic tissues could only divide a finite number of times in culture, usually around 50 Cumulative Population Doublings (CPDs). This phenomenon herein called Replicative Senescence’ (RS) (also known as ‘Hayflick phenomenon’). They presciently hypothesized the existence of cellular factors, whose loss through consecutive cell divisions limited the proliferation of normal cells (Campisi and d'Adda di Fagagna, 2007).
Cellular senescence is a signal transduction program leading to irreversible cell cycle arrest. This growth arrest can be triggered by many different mechanisms including recognition by cellular sensors of DNA double-strand breaks leading to the activation of cell cycle checkpoint responses and recruitment of DNA repair foci. To picture the main causes of senescence, Campisi and d'Adda di Fagagna (2007) categorized them as; telomerase dependent senescence, DNA-damage initiated senescence, senescence caused by chromatin perturbation, oncogene-induced senescence and stress-induced senescence (Fig. 1).
Senescence is initiated by the shortening of telomeres (telomeric senescence or replicative senescence) or by other endogenous and exogenous acute and chronic stress signals (STASIS: stress or aberrant signaling-induced senescence) (Shay and Roninson, 2004). Briefly, ‘replicative senescence’ is defined to senescence that occurs following extended proliferation, presumably triggered by a cell-intrinsic mechanism. Stress-induced premature senescence, accelerated senescence or extrinsic senescence is referred to rapid senescence triggered by non-telomeric signals or extrinsic stress (Ben-Porath and Weinberg, 2005) of which oxidative stress plays a fundamental role (Campisi, 2001; Campisi and d'Adda di Fagagna, 2007; Harman, 1981). In Mammals, cellular senescence is confined to mitotic cells, able to proliferate unlike the post-mitotic cells which permanently lose the ability to divide owing to differentiation. The senescent cells often share similar features known as ‘senescent phenotype’ which include; resistance to cell-death signals (apoptosis resistance), changes in gene expression (altered gene expression) and stopped proliferation (growth arrest).
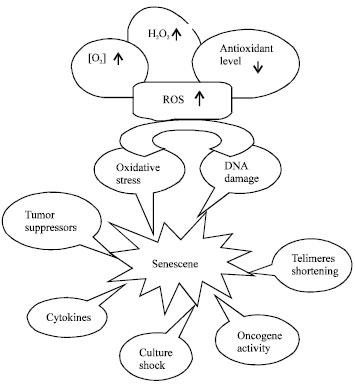 | |
| Fig. 1: | Multi-stimuli induce cellular senescence. High levels of ambient oxygen concentration and H2O2, low level of antioxidants lead to ROS formation. Consequently, DNA damages occur and redox pathways are activated. Intrinsic senescence triggers by telomere shortening while DNA damaging, oncogenic signaling, oxidative stress, overexpression of tumor suppressors, cytokines and undefined stress signals (culture shock) induce extrinsic senescence |
The features and stringency of the senescence growth arrest vary depending on the species and the genetic background of the cell (Campisi and d'Adda di Fagagna, 2007).
Suppression of genes required for cell cycle progression and upregulation of growth inhibitory genes may lead to senescence. Cells may undergo senescence by overexpression of certain oncoproteins (e.g., RAS-RAF-MEK signaling cascade) or while the DNA damage is irreversible and/or irreparable. Premature senescence may appear due to the activation or overexpression of major tumor suppressor genes including p16INK4a Cyclin-Dependent Kinase Inhibitor (CDKI), p21WAF1 and p27 (McConnell et al., 1998).
Extensive investigations have shown some oncogenes such as activated Ras (H-RasV12) trigger a permanent arrest that might induce replicative senescence. The tumor suppressor p16INK4a which activates p53 together with general cell cycle regulators pRb, p107 and p130, plays a key roles in Ras-induced senescence (Serrano et al., 1997). Various oncogenesis pathways were shown to cause premature senescence.
Overall, telomere uncapping can trigger a DNA damage response that results in senescence-like growth arrest. It was found that telomerase shortening-induced senescence acts as a DNA damage response which is mediated by the ATM/ATR-p53-p21 pathway (Herbig et al., 2004). In other way, telomerase dysfunction activates DNA damage resulting in cell cycle growth arrest. Thereby, the agents that trigger DNA damage might manipulate the cellular genome and induce senescence. It is believed that inactivation of p53 could inhibit the DNA-damage induced senescence (Ongusaha et al., 2003). The p53 protein is stabilized and proceeds to p21CIP1/WAF leading to premature senescence (Ben-Porath and Weinberg, 2005). P53 and pRb were found to be essential for the initiation and maintenance of the senescent state. These findings suggested a linear signaling model; therefore a stress signal activates p53 which in turn activates pRb. The p21WAF1 protein, an inhibitor of cyclin E/Cdk2 complexes which is a direct transcriptional target of p53 was proposed to link these two pathways. However, p21 undergo RAS-induced senescence (Pantoja and Serrano, 1999). In general, p53 plays a prominent role in mediating the response to telomere dysfunction, DNA damage and oxidative stress.
Cells might undergo premature senescence by overexpression of viral oncoproteins such as the SV40 large T-antigen and the human papillomavirus E6 and E7 proteins (Ben-Porath and Weinberg, 2005; Wright and Shay, 2001), germline homologous recombination in the mouse and somatic homologous recombination in human cells, nuclear injection of antibodies dominant-negative forms of these proteins and RNAi expression knockdown methods (Ben-Porath and Weinberg, 2005).
ANTI-AGING PRODUCTS; FACT OR FICTION?
In general, an anti-aging product expected to prevent, reduce or treat age-related features along with maximizing the quality and quantity of the normal life-span of an individual. Anti-aging products (medicines) with their multiple claims cover mainly two broad categorizes; ‘cosmeceutical age related products’ and ‘anti-aging drugs’. Cosmeceuticals are referred to substances that exerted both cosmetic and therapeutic benefits. Statistical shows cosmeceuticals and anti-aging products were sold more than $16 billion by 2010 in USA (Rivers, 2008). Anti-aging drugs are associated with age-dependent degenerative diseases such as; diabetes, atherosclerosis, hypertension, macular degeneration, cancer, Alzheimer's disease and osteoarthritis.
There are scientific controversy regarding anti-aging nutritional supplementation and medicine. Scientist’s legitimate efforts emphasize to clearly discriminate between the anti-aging quackery and those valuable efforts that lead to recover or delay aging processes. Wick (2002) claimed in addition to questionable and even harmful effects of anti-aging products, many of them were found not to meet the quantitative and/or qualitative criteria given on the label. Olshansky et al. (2002) have criticized the anti-aging industry in response to what they see as unscrupulous profiteering from the sale of unproven anti-aging supplements. Recommendation of anti-aging products may damage the credibility of experimental gerontology as the scientific discipline that focuses on studies of the aging process to understand the fundamental of aging and subsequently use this knowledge to extend the quantity, while maintaining the quality, of human life (Wick, 2002).
Proper diet and calorie intake as well as regular exercise and good environmental conditions are known to contribute to anti aging. In addition to certain super foods, some herbal preparations have been shown to have anti-oxidant property contributing to delaying the aging process (Dhar, 2009).
AGING AND OXIDATIVE STRESS
What is oxidative stress? In Sies (1985); oxidative stress has been defined as ‘a disturbance in the pro-oxidant/antioxidant balance in favour of the former’. Thus oxidative stress is essentially an imbalance between the production of various reactive species and the ability of the organism’s natural protective mechanisms to cope with these reactive compounds and prevent adverse effects. The oxidative stress concept was redefined as ‘an imbalance between oxidants and antioxidants in favor of the oxidants leading to a disruption of redox signaling and control and/or molecular damage’ (Veskoukis et al., 2012). Instate, an antioxidant characterizes a substance that, when present at low concentrations compared with those of an oxidizable substrate, significantly prevents or delays a pro-oxidant initiated oxidation of the substrate (Abdollahi et al., 2004). A pro-oxidant is a toxic substance that causes oxidative damage to lipids, proteins and nucleic acids, resulting in various pathologic events and/or diseases. Pro-oxidant is a synonym for reactive species (Prior and Cao, 1999).
Antioxidants can be categorized into nonenzymatic and enzymatic ROS scavengers. Nonenzymatic antioxidants include dietary compounds, such as vitamins (C and E) and minerals (selenium and zinc) and also glutathione, uric acid, tocopherol, retinol and ubiquinol. Superoxide Dismutase (SOD), Catalase (CAT), γ-Glutamyl Transpeptidase (GT), glutathione reductase and glutathione peroxidase (GPX) are the main enzymatic antioxidants. The sum of all known and unknown endogenous and exogenous antioxidants in a medium is usually called Total Antioxidant Capacity (TAC) and gives a holistic view of antioxidant status (Rezaie et al., 2007).
ROS formation: Reactive species are unstable, highly reactive structures. They may be free radicals with half-lives vary from a few nanoseconds to seconds and hours. They trigger chain reactions resulting in the oxidation of macromolecules in order to reach a steady state. They are divided into four main categories based on their central atom; ROS, Reactive Nitrogen Species (RNS), reactive sulfur species and reactive chloride species (Halliwell and Gutteridge, 2007).
ROS are oxygen-derived small molecules, including oxygen radicals [superoxide (O2-●), hydroxyl (HO●), peroxyl (RO2●) and alkoxyl (RO●)] and certain non-radicals that are either oxidizing agents and/or are easily converted into radicals, such ashypochlorous acid (HOCl), Ozone (O3), singlet oxygen (1O2) and hydrogen peroxide (H2O2) (Bedard and Krause, 2007). The concentration of reactive species is a key factor in their effect. If the concentration is excessively high, they may become very harmful for macromolecules (Veskoukis et al., 2012).
ROS are generated in multiple compartments and by multiple enzymes within the cell and external pro-oxidant stimuli, such as radiations, pathogen infections, herbicides/pesticides, toxins and Ultra Violet (UV). ROS are formed via proteins within the plasma membrane (e.g., NADPH oxidases); lipid metabolism within the peroxisomes; activation of various cytosolic enzymes (e.g., cyclooxygenases). The majority of cellular ROS (approximately 90%) is generated within the mitochondria describing its crucial role to accelerate premature senescence (mitochondrial theory of aging) (Balaban et al., 2005; Burdon, 1995; Droge, 2002).
Oxidative phosphorylation generates a potential energy for protons (ΔΨ) across the mitochondrial inner membrane via the oxidation of NADH or FADH (Balaban et al., 2005). The generated ATP is coupled with a reaction in which O2 is reduced to H2O. It is believed under certain conditions, O2 can also be reduced to H2O via superoxide anion (O2-●) and H2O2. It is recognized that the cellular production of (O2-●) and H2O2 favors the formation of other reactive oxygen and nitrogen species (i.e., (HO●) and peroxynitrite (ONOO¯) and excessive production of these species causes oxidative stress (Lopez-Lazaro, 2007).
Bedard and Krause (2007) postulated ROS generation as a cascade of reactions that starts with the production of superoxide. Thereafter, (O2-●) rapidly dismutates to (HO●) either spontaneously or catalyzed by SOD. Afterward, superoxide reacts with nitric oxide to form (ONOO¯), the peroxidase-catalyzed formation of hypochlorous acid from hydrogen peroxide and the iron-catalyzed Fenton reaction leading to the generation of hydroxyl radical. During Fenton reactions, hydrogen peroxide is partially converted into the extremely reactive (HO●), which immediately attack surrounding macromolecules, including DNA, proteins and lipids. Hydroxyl radicals exert their pathogenic activity also indirectly, by reacting with unsaturated fatty acids under the formation of organic peroxides, which decompose to aldehydes. The latter are responsible for a number of toxic effects, including the formation of protein–protein aldehyde bridges (Terman and Brunk, 2006).
ROS and aging: Harman (1956) suggested ROS formation induces macromolecular damages leading to progressive deleterious changes that were termed aging or senescence. The “oxidative stress theory” defines a progressive and irreversible accumulation of ROS-induced oxidative damage influencing the aging process (Kregel and Zhang, 2007). A key assumption of the free radical hypothesis of aging is that the endogenous antioxidant defenses of aerobic cells are deficient and the residual prooxidants exert a certain level of oxidative stress even under normal physiological conditions (Sohal and Orr, 1992).
There are undeniable relevant between the multi-stimuli of aging. Increases in interacellular ROS level lead to DNA damage through erroneous repair or replication, which has the possibility to be converted into mutations. Mutations are well recognized to cause cancer and have also been proposed to initiate senescence (Vijg, 2000). It is apparent that ROS implicate in both cellular senescence and in organismal aging (Beckman and Ames, 1998). Indeed, mice that are genetically deficient for the p66SHC gene are less sensitive to the toxic effects of ROS at both the cellular and organismal level and such mutant mice live 30% longer than their wild-type littermates, indicating that in these mice the process of organismal aging is delayed (Migliaccio et al., 1999; Lundberg et al., 2000).
Oxidants might non-specifically induce a spectrum of damage to cellular components (e.g., DNA) that directly leads to senescence or this damage might induce senescence by secondarily activating important intracellular pathways, such as the DNA damage response (Lu and Finkel, 2008). Oxidative stress may activate numerous intracellular signaling pathways via ROS-mediated modulation of various enzymes and critical transcription factors. Increases of ROS level activate transcription factors or oxidative damage is transferred to the nucleus within a cell and bind to promoter regions of particular genes. Consequently, the stress-activated pathways might impact the gene expression and the cell might undergo apoptosis, proliferation or cytokines (Kregel and Zhang, 2007; Abdollahi et al., 2005; Kajbaf et al., 2007; Najafi et al., 2009).
Oxidants may directly activate certain redox-sensitive pathways linked to senescence. The activation of redox-sensitive transcriptional factors by age-related oxidative stress causes the upregulation of pro-inflammatory gene expression. As a result, various pro-inflammatory molecules are generated, leading to an inflammation cascade associated with different age related pathologies such as cancer, cardiovascular diseases, arthritis and several neurodegenerative diseases (Chung et al., 2006; Kregel and Zhang, 2007). Recently, the efficacy of different antioxidants in various diseases like diabetes (Afshari et al., 2004; Hasani-Ranjbar et al., 2009, 2010a, 2011; Radfar et al., 2005; Rahimi et al., 2005), Langerhans islet transplantation (Larijani et al., 2011; Mohammadi et al., 2011; Mohseni-Salehi-Monfared et al., 2009), Alzheimer’s disease (Nunomura et al., 2006), inflammatory and oxidant-related diseases (Hasani-Ranjbar et al., 2009, 2010b; Rahimi et al., 2010; Malekirad et al., 2011; Mohammadirad et al., 2011; Momtaz and Abdollahi, 2010; Mousavi et al., 2010), rheumatoid arthritis (Abdollahi et al., 2005; Mirshafiey and Mohsenzadegan, 2008), cancers (Izzo and Ernst, 2009; Shadnia et al., 2005) and inflammatory bowel disease (Rahimi et al., 2009; Rezaie et al., 2007; Rastegarpanah et al., 2011) has been systematically reviewed. Antioxidants have been also proven to reduce the harmful effects of xenobiotic exposures (Abdollahi et al., 2004; Karami-Mohajeri and Abdollahi, 2010; Soltaninejad and Abdollahi, 2009; Mohammadirad and Abdollahi, 2011; Shadnia et al., 2011).
One of the effective factors on cellular senescence has been recognized to be ambient oxygen concentration (Parrinello et al., 2003). Different studies have clarified, since intracellular oxidants increase by altering ambient oxygen concentrations or lowering antioxidant levels, this accelerates the onset of senescence while lowering ambient oxygen or increasing ROS scavenging appears to delay senescence (Lu and Finkel, 2008). For instance, human fibroblasts experienced premature senescence when cultured in high ambient oxygen conditions (40-50%), while their proliferative lifespan significantly extended when grown in low ambient oxygen (2-3%) more likely to physiological oxygen levels (Chen et al., 1995). Even, telomere shortening is also dependent on ambient oxygen concentration. Senescence induced by oxidative stress might be as result of a telomere-dependent and -independent way. Exposure to mild oxidative stress leads to faster telomere shortening, whereas maintenance of telomere length improves under low oxidative stress (Von Zglinicki, 2002). The rate of telomere shortening is accelerated in fibroblasts grown in high oxygen conditions (Forsyth et al., 2003).
The concentration of hydrogen peroxide appears important as very high concentrations of ROS appear to trigger apoptosis while lower concentrations appear to favor senescence (Colavitti and Finkel, 2005). The cells treated with hydrogen peroxide induced a predominant G1 arrest with an increase in p53 protein levels and increased p53 activity, including the subsequent overexpression of p21 (Chen et al., 1998). Another study exhibited the reduction of ambient oxygen level decreases the proportion of pre-senescent p21-expressing cells without any effect on p16-expressing cells. This explains that oxidative stress-induced senescence, like telomere dysfunction operates through the p53-p21-Rb axis (Itahana et al., 2003).
In normal diploid fibroblasts, Ras expression leads to overexpression of p16 and p21 which results a telomere-independent growth arrest. More interestingly, in primary cells, increased Ras expression enhances intracellular ROS levels which appears to be important for cellular senescence. Since culturing Ras expressing cells in either low oxygen or treating these cells with a hydrogen peroxide scavenging agent (e.g., N-acetylcysteine (NAC)) blocked Ras-induced senescence (Lee et al., 1999; Serrano et al., 1997).
It was reported that p16INK4a/pRb pathway could increase intracellular ROS levels through activation of PKCδ (Protein Kinase C δ), which, in turn seems to lead to further production of ROS by activating NADPH oxidase, thus establishing a self-sustained positive feedback loop. In addition, the activation of PKCδ results in depletion of WARTS protein (WTS/large tumour-suppressor 1 mitotic kinase) expression, the kinase essential in cytokinesis and G2M cell cycle arrest which act as a second barrier ensuring irreversibility of senescence (Takahashi et al., 2006). Other studies indicate that oxidative stress can induce p16, through the action of p38 MAPK, a member of the Stress-Activated Protein Kinase family (SAPK) (Iwasa et al., 2003). Another study implicated enhancement of ROS level in H2O2-induced senescence cells causes the activation of p38 by increasing the expression of a subset of p38 MAPK-dependent genes (Zdanov et al., 2006).
Antioxidants and aging: Any substance that delays, prevents or removes oxidative damage to a target molecule is called an antioxidant.
| Table 1: | Plant species with anti-aging effects |
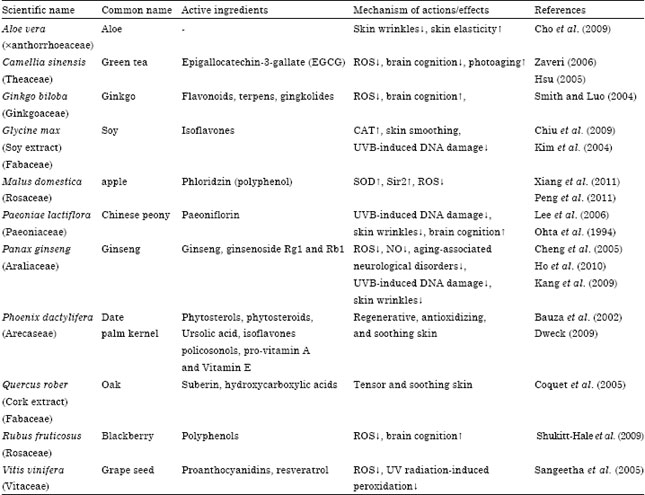 | |
Since oxidative stress counts as a fundamental reason inducing both cellular and organismal senescence, thus antioxidants could be considered as either senescence inhibitors or even as treatment of aging. As mentioned earlier, animals are dealing with different endogenous and exogenous barriers, consequently are equipped with defense mechanisms. In between, animal body faces oxidative stress through both enzymatic and nonenzymatic systems. Regarding their origin, various antioxidants such as glutathione, uric acid, CAT and SOD can be synthesized in vivo, whereas others, namely; polyphenols and β-carotene, are obtained from food. Based on their physical properties, antioxidants can be divided into water soluble antioxidants such as uric acid, glutathione and polyphenols or lipid-soluble antioxidants such as vitamins A and E and lipoic acid. Due to the inadequacy of antioxidant defenses, cells are under a certain level of oxidative stress even under normal physiological conditions. Following sections will describe the enzymatic antioxidants, later, polyphenols and the important plant species/components correlated with aging process will introduce (Table 1).
SOD, GPX and CAT: An imbalance in the ratio of SOD to GPX and CAT results in the accumulation of H2O2 which may participate in the Fenton reaction, resulting in the formation of noxious hydroxyl radicals. This obviously lead to oxidative stress and results in premature cellular senescence. It has been observed that enhancement of lipid peroxidation in the brain of aging mice could change Cu/Zn-superoxide dismutase (SOD1)/(GPX1 plus CAT) ratio. Therefore, it was proposed the balance in the activity of the SOD to GPX plus CAT ratio (SOD/(GPX plus CAT)) is an important determinant of cellular aging (De Haan et al., 1995).
SOD catalyzes dismutation of superoxide anions to hydrogen peroxide. SOD1 and SOD2 are counted as two intracellular SOD enzymes which convert O2¯ into H2O2 that is then further deactivated by CAT to water and oxygen or by the various glutathione peroxidases (GPXs) to reduced glutathione and water. Serra et al. (2003) reported increasing the level of SOD expanded the life span of primary fibroblasts as well as decreasing the rate of telomere shortening. Another study showed reduction of SOD resulted in the induction of p53 which is essential for senescence to happen (Balaban et al., 2005).
GPX catalyzes the degradation of H2O2 and hydroperoxides originating from unsaturated fatty acids at the expense of reduced glutathione. Among the enzymatic antioxidants, GPX is considered one of the most important metabolites and is the first line of defense against reactive species. Lipid hydroperoxides are destroyed by glutathione peroxidase (Ames et al., 1993). Glutathione is important for the regeneration of antioxidant vitamins E and C. GPX participates in several different biochemical processes; in the regulation of protein and DNA synthesis and as an essential cofactor of many enzymes (Halliwell and Gutteridge, 2007; Veskoukis et al., 2012).
CAT converts H2O2 into molecular oxygen and water. In a study, CAT activity increased with age suggests an increase in H2O2 formation in erythrocytes (Inal et al., 2001). Conversely, Goth (1989) found low CAT activity in the aging process. Hydrogen peroxide is removed by CAT and GPX activities in erythrocytes (Andersen et al., 1997).
Polyphenolic compounds: Polyphenolics cover broad classes of compounds with decisive antioxidant properties. As antioxidants, polyphenols may protect cell constituents against oxidative damage and, therefore, limit the risk of various degenerative diseases associated to oxidative stress. Plentiful evidences indicate that polyphenols might exert several other specific biological effects such as; the inhibition or reduction of different enzymes such as telomerase, cycloxygenase, the interaction with signal transduction pathways and cell receptors (D’Archivio et al., 2007).
Polyphenolics are plant secondary metabolites (a part of plant’s defense system) which have at least one aromatic ring in their molecule and usually exist in the form of glycosides. These compounds may be classified into different groups as a function of the number of phenol rings that they have. Distinctions are thus made between the phenolic acids, flavonoids, stilbenes and lignans (Manach et al., 2004).
In addition to the protective effects of endogenous enzymatic antioxidant defenses, consumption of dietary antioxidants appears to be of great importance. Fruits and vegetables, the main source of antioxidants in the diet, are associated with a lowered risk of degenerative aging diseases (Ames et al., 1993). Galli et al. (2002) showed increasing dietary intake of polyphenols, can retard and even reverse age-related declines of brain function, cognition and motor performance in rats. Antioxidants dietary supplementation can decrease the vulnerability to oxidative stress in vivo by reductions in neuronal signaling.
Flavonoids and aging: The general flavonoid structure is a flavan nucleus, 15 carbon atoms arranged in three rings (A, B, C). The various classes differ in their level of oxidation and in their pattern of substitution of the c-ring. Anti-aging potential of polyphenolics of which flavonoids considered as a central group was studied extensively. Flavonoids were shown to activate key enzymes in mitochondrial respiration and to protect neuronal cells by acting as antioxidants, thus breaking the vicious cycle of oxidative stress and tissue damage. Flavonoids possess a highly reactive hydroxyl group that gets oxidized by electron-donation, thus stabilizing the radical to a less reactive molecule. An investigation revealed flavonoids efficiently attenuate the deleterious effect of free radicals and ROS/RNS (Schmitt-Schillig et al., 2005). Many flavonoids are known to possess anti-aging properties in terms of prevention or treatment. In between some compounds attracted more attentions; catechins of green tea, resveratrol of grape extract, anthocyanins of berry extracts and Vitamin E.
Resveratrol: Resveratrol (3,5,4'-trihydroxystilbene) was first isolated from the roots of white hellebore (Veratrum grandiflorum O. Loes) in 1940 and later was found in grapes and red wine. Resveratrol exhibited beneficial effects in mammals to prevent or delay the onset of cancer, heart disease, ischaemic and chemically induced injuries, diabetes, pathological inflammation, viral infection and possibly might extend the lifespans of various organisms from yeast to vertebrates (Baur and Sinclair, 2006). Majority of evidences confirm resveratrol might perform its life expanding activity through SIR family and mimics the effects of CR on life extending (Alarcon de la Lastra and Villegas, 2005; Denu, 2003; Lin et al., 2000). Recently, gene expression profiling suggested both CR and resveratrol may retard some aspects of aging through alterations in chromatin structure and transcription in rat (Barger et al., 2008).
Catechins: Japanese and Chinese are the first populations consuming green tea. Originally, green tea is the unfermented leaves of Camellia sinensi that are steamed and dried to inactivate the polyphenol oxidase enzyme, a process that essentially maintains the polyphenols in their monomeric forms. Several polyphenolic catechins were isolated from green tea; -(-) Epicatechin (EC), (-) Epicatechin-3-gallate (ECG), (-) Epigallocatechin (EGC), (-) Epigallocatechin-3-Gallate (EGCG), (+) catechin and (+) Gallocatechin (GC). The main antioxidant properties of green tea are attributed to catechins particularly EGCG, which is the most abundant with about 65% of the total catechin content (Zaveri, 2006). Green tea catechins are rapidly absorbed and are distributed mainly into the mucous membranes of the small intestine, liver and the plasma. The antioxidative capacity of human plasma is increased by consumption of green tea constantly (Kimura et al., 2002).
Consumption of green tea catechin provokes antioxidative enzymes (Khan et al., 1992). Unno et al. (2004) reported intake of green tea catechin partially improves the morphologic and functional alterations that occur naturally in the brains of aged mice with accelerated senescence (SAMP10 mice). Catechins exhibited protective effect on cognitive dysfunction and suppressed cerebral atrophy in these animals. Furthermore, green tea also decreased the levels of 8-oxo-deoxyguanosine (8-oxodG), a marker of oxidative DNA damage, in mice kidney, liver and cerebrum, suggesting that green tea polyphenols may have a beneficial effect on aging-induced damage (Zaveri, 2006).
Anthocyanins: Anthocyanins differ from other natural flavonoids in the range of colors that can be derived from them and by their ability to form resonance structures by changes in pH. Anthocyanins serve as the major components of different berries. Anthocyanins maintain DNA integrity and exhibit potential antioxidant properties. Studies have shown that supplementations with berries rich in anthocyanins were effective in reducing oxidative stress associated with aging and were beneficial in reversing age-related neuronal and behavioral changes. Supplementation with anthocyanins for 6-8 months retarded age-related declines in neuronal and cognitive function by improving antioxidant status (Bagchi et al., 2004).
Vitamin E and aging: Vitamin E is a ubiquitous natural compound derived from plant kingdom. The term vitamin E embraces all tocopherols and tocotrienols showing the biological activity of the isomer RRR-α-tocopherol. Chemically, tocopherol is 6-chromanol derivative. There are four forms of tocopherol esteroisomers (α, β, δ, γ) depending on the different substituent positions on its chromane ring. There is also a synthetic form of vitamin E derived from phytol that is called all-rac-α-tocopherol with lower biological activity than the natural one (Nachbar and Korting, 1995). The antioxidative activity of Vitamin E has been confirmed in numerous in vitro and in vivo experiments (Huang et al., 1988; McCall and Frei, 1999; Roob et al., 2000). Among the natural forms of tocopherol having Vitamin E activity, α-tocopherol has the greatest physiological effects, including suppression of lipid peroxidation (Bjorneboe et al., 1990). Vitamin E is recognized to be the most natural accruing lipid-soluble antioxidant in human tissue (Chow, 1991; Furuse, 1987; Nachbar and Korting, 1995).
Vitamin E is believed to be a food component that may have an anti-aging effect (Meydani, 1992). To be considered as an anti-aging, vitamin E has raised various controversies. The photoperotective effect of Vitamin E has been proven years ago (Werninghaus et al., 1991; Kagan et al., 1992; Mayer, 1993; Nachbar and Korting, 1995). Thereby, scientific reasonable documents on its anti-aging properties have not been found yet.
Ginseng and aging: Ginseng, the root and rhizome of Panax ginseng C A Meyer., has been known as a most precious medicine in China, Japan and Korea for centuries. Modern medicine also identified different pharmacological properties for Ginseng as; increasing immune function, improvement of brain function, enhance central cholinergic system function, inhibition of free radical and NO generation and promotion of proliferation of rodent progenitor cells in vitro and in vivo (Cheng et al., 2005; Choi, 2008). Ginseng, ginsenoside Rg1 and Rb1 (isolated compounds), have the ability to interfere almost all of the pathways that accelerate aging process (Cheng et al., 2005). Pharmacologically, polyphenols are the active antioxidant constituents of Ginseng which might benefit aging-related diseases.
Ginkgo biloba and aging: Ginkgo biloba is believed to be the oldest tree on earth, more than 200 million years antiquity. Scientific data strongly support the efficacy of Ginkgo for treatment of age-related disorders (Gaby, 1996). Ginkgo portraits great biological activities of which, improvement of brain activity and blood circulation, absorb the most attention. Two major classes of compounds; flavonoids and terpens are responsible for unique properties of this species. The antioxidant effect of its flavonoids may be achieved by direct attenuation of reactive oxygen species, thereby decreasing tissue levels of ROS and inhibition of membrane lipid peroxidation (Smith and Luo, 2004).
Oken et al. (1998) estimated Ginkgo extract is capable to improve cognition function of brain in Alzheimer Disease (3% difference). Gingkolides the most important isolated compound from Ginkgo biloba demonstrates antioxidant, neuroprotective and cholinergic activities relevant to Alzheimer's disease mechanisms. In addition, the therapeutic efficacy of Ginkgo extracts in Alzheimer's disease reported to be similar to currently prescribed drugs such as tacrine or donepezil with minimal side effects in clinical trials (Perry et al., 1999). Another study showed Ginkgo extract (EGb 761), increased stress resistance and extends organism’s life span in Caenorabditis elegans (Wu et al., 2002).
β-carotene and aging: β-carotene is one of more than 600 carotenoids known to exist in nature. About 50 of the naturally occurring carotenoids can potentially yield vitamin A and are thus referred to as provitamin A. β-carotene is the most abundant and most efficient provitamin A and is known to quench singlet oxygen. Some authors concluded β-carotene dose not possess antioxidant protective effects against chronic disease as great as Vitamin E. Probably, the majority of β-carotene's beneficial in vivo, is not derived from its antioxidant properties but rather it might grow from its effect on a number of biochemical systems (Pryor et al., 2000). Eicker et al. (2003) reported β-carotene is capable of protecting fibroblast from the induction of photoaging-associated mitochondrial DNA (mtDNA) deletions. They also exhibited in a dose dependent manner β-carotene interacts with UVA radiation in the cell and shows protective properties from the induction of a photoaging-associated mtDNA mutation.
CONCLUSION
Up to date studies indicate that genetic, nutrition, physical activity and environmental conditions could affect lifespan. A better understanding of the causes of senescence or age-related dysfunctions might open new ways to slow aging. As a result, remarkable aging risk factors could be distinguished, avoided and their harmful effects could be minimized. Additionally, the more sensitive body parts to degenerative effects of aging might be recognized. Up to date, animal models have shown promising differences in lifespan while genetically modified. However, the relevance of some animal models to human aging is unclear. Literature review suggests that ROS participate in aging process, therefore many investigations have been conducted to delay aging by reducing free radical damages. So far, more effective explorations are needed to decrease the deleterious effects of ROS-induced senescence. This approach could be achieved through manipulations of antioxidant enzyme systems, development of drug therapies, manipulation of senescence related genes and genetic manipulation of plant-based antioxidant resources.
ACKNOWLEDGMENT
This study is the outcome of an in-house non-financially supported study.
REFERENCES
- De la Lastra, C.A. and I. Villegas, 2005. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res., 49: 405-430.
CrossRefDirect Link - Izzo, A.A. and E. Ernst, 2009. Interactions between herbal medicines and prescribed drugs: An updated systematic review. Drugs, 69: 1777-1798.
CrossRefPubMedDirect Link - Atwood, C.S., S.V. Meethal, T. Liu, A.C. Wilson, M. Gallego, M.A. Smith and R.L. Bowen, 2005. Dysregulation of the hypothalamic-pituitary-gonadal axis with menopause and andropause promotes neurodegenerative senescence. J. Neuropathol. Exp. Neurol., 64: 93-103.
PubMedDirect Link - Balaban, R.S., S. Nemoto and T. Finkel, 2005. Mitochondria, oxidants and aging. Cell, 120: 483-495.
PubMed - Bedard, K. and K.H. Krause, 2007. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev., 87: 245-313.
CrossRefDirect Link - Ben-Porath, I. and R.A. Weinberg, 2005. The signals and pathways activating cellular senescence. Int. J. Biochem. Cell. B., 37: 961-976.
PubMedDirect Link - Bjorneboe, A., G.E. Bjorneboe and C.A. Drevon, 1990. Absorption, transport and distribution of vitamin E. J. Nutr., 120: 233-242.
PubMedDirect Link - Campisi, J. and F. d'Adda di Fagagna, 2007. Cellular senescence: When bad things happen to good cells. Nat. Rev. Mol. Cell. Biol., 8: 729-740.
CrossRefDirect Link - Chen, Q., A. Fischer, J.D. Reagan, L.J. Yan and B.N. Ames, 1995. Oxidative DNA damage and senescence of human diploid fibroblast cells. Proc. Natl. Acad. Sci. USA, 92: 4337-4341.
Direct Link - Chen, Q.M., J.C. Bartholomew, J. Campisi, M. Acosta, J.D. Reagan and B.N. Ames, 1998. Molecular analysis of H2O2-induced senescent-like growth arrest in normal human fibroblasts: p53 and Rb control G1 arrest but not cell replication. Biochem. J., 332: 43-50.
Direct Link - Cheng, Y., L.H. Shen and J.T. Zhang, 2005. Anti-amnestic and anti-aging effects of ginsenoside Rg1 and Rb1 and its mechanism of action. Acta Pharmacol. Sin., 26: 143-149.
CrossRefPubMedDirect Link - Cho, S., S. Lee, M.J. Lee, D.H. Lee, C.H. Won, S.M. Kim and J.H. Chung, 2009. Dietary aloe vera supplementation improves facial wrinkles and elasticity and it increases the type I procollagen gene expression in human skin in vivo. Ann. Dermatol., 21: 6-11.
CrossRefPubMedDirect Link - Choi, K.T., 2008. Botanical characteristics, pharmacological effects and medicinal components of Korean Panax ginseng C A Meyer. Acta Pharmacol. Sinica, 29: 1109-1118.
CrossRefPubMedDirect Link - Chung, H.Y., B. Sung, K.J. Jung, Y. Zou and B.P. Yu, 2006. The molecular inflammatory process in aging. Antioxid. Redox Signaling, 8: 572-581.
CrossRefPubMedDirect Link - Colavitti, R. and T. Finkel, 2005. Reactive oxygen species as mediators of cellular senescence. IUBMB Life, 57: 277-281.
CrossRefPubMedDirect Link - Collado, M., M.A. Blasco and M. Serrano, 2007. Cellular senescence in cancer and aging. Cell, 130: 223-233.
CrossRefPubMedDirect Link - Coquet, C., E. Bauza, G. Oberto, A. Berghi and A.M. Farnet et al., 2005. Quercus suber cork extract displays a tensor and smoothing effect on human skin: An in vivo study. Drugs Exp. Clin. Res., 31: 89-99.
PubMedDirect Link - D'Archivio, M., C. Filesi, R. Di Benedetto, R. Gargiulo, C. Giovannini and R. Masella, 2007. Polyphenols, dietary sources and bioavailability. Ann. Ist. Super. Sanita, 43: 348-361.
PubMedDirect Link - De Haan, J.B., F. Cristiano, R.C. Iannello and I. Kola, 1995. Cu/Zn-superoxide dismutase and glutathione peroxidase during aging. Biochem. Mol. Biol. Int., 35: 1281-1297.
PubMedDirect Link - Denu, J.M., 2003. Linking chromatin function with metabolic networks: Sir2 family of NAD+-dependent deacetylases. Trends Biochem. Sci., 28: 41-48.
Direct Link - Dweck, A.C., 2009. The internal and external use of medicinal plants. Clin. Dermatol., 27: 148-158.
CrossRefPubMedDirect Link - Eicker, J., V. Kurten, S. Wild, G. Riss, R. Goralczyk, J. Krutmann and M. Berneburg, 2003. Betacarotene supplementation protects from photoaging-associated mitochondrial DNA mutation. Photochem. Photobiol. Sci., 2: 655-659.
PubMedDirect Link - Forsyth, N.R., A.P. Evans, J.W. Shay and W.E. Wright, 2003. Developmental differences in the immortalization of lung fibroblasts by telomerase. Aging Cell, 2: 235-243.
CrossRefPubMedDirect Link - Furuse, K., 1987. Vitamin E: Biological and clinical aspects of topical treatment. Cosmet. Toiletries, 102: 99-116.
Direct Link - Galli, R.L., B. Shukitt-Hale, K.A. Youdim and J.A. Joseph, 2002. Fruit polyphenolics and brain aging: Nutritional interventions targeting age-related neuronal and behavioral deficits. Ann. N.Y. Acad. Sci., 959: 128-132.
CrossRefPubMedDirect Link - Goth, L., 1989. Effect of age, sex and smoking on serum catalase activity. Acta Biol. Hung., 40: 395-399.
PubMedDirect Link - Hasani-Ranjbar, S., B. Larijani and M. Abdollahi, 2011. Recent update on animal and human evidences of promising antidiabetic medicinal plants: A mini-review of targeting new drugs. Asian J. Anim. Vet. Adv., 6: 1271-1275.
CrossRefDirect Link - Herbig, U., W.A. Jobling, B.P.C. Chen, D.J. Chen and J.M. Sedivy, 2004. Telomere shortening triggers senescence of human cells through a pathway involving ATM, p53 and p21CIP1, but not p16INK4a. Mol. Cell, 14: 501-513.
CrossRefPubMedDirect Link - Holliday, R., 2006. Aging is no longer an unsolved problem in biology. Ann. N. Y. Acad. Sci. USA., 1067: 1-9.
CrossRefPubMedDirect Link - Hsu, S., 2005. Green tea and the skin. J. Am. Acad. Dermatol., 52: 1049-1059.
CrossRefPubMedDirect Link - Huang, N., B. Lineberger and M. Steiner, 1988. α-tocopherol, a potent modulator of endothelial cell function. Thrombosis Res., 50: 547-557.
PubMedDirect Link - Itahana, K., Y. Zou, Y. Itahana, J.L. Martinez and C. Beausejour et al., 2003. Control of the replicative life span of human fibroblasts by p16 and the polycomb protein Bmi-1. Mol. Cell Biol., 23: 389-401.
PubMed - Iwasa, H., J. Han and F. Ishikawa, 2003. Mitogen-activated protein kinase p38 defines the common senescence-signalling pathway. Genes Cells, 8: 131-144.
CrossRef - Kagan, V., E. Witt, R. Goldman, G. Scita and L. Packer, 1992. Ultraviolet light-induced generation of vitamin E radicals and their recycling. A possible photosensitizing effect of vitamin E in skin. Free Radic. Res. Commun., 16: 51-64.
PubMedDirect Link - Kang, T.H., H.M. Park, Y.B. Kim, H. Kim and N. Kim et al., 2009. Effects of red ginseng extract on UVB irradiation-induced skin aging in hairless mice. J. Ethnopharmacol., 123: 446-451.
CrossRefPubMedDirect Link - Kim, S.Y., S.J. Kim, J.Y. Lee, W.G. Kim, W.S. Park, Y.C. Sim and S.J. Lee, 2004. Protective effects of dietary soy isoflavones against UV-induced skin-aging in hairless mouse model. J. Am. Coll. Nutr., 23: 157-162.
CrossRefPubMedDirect Link - Kimura, M., K. Umegaki, Y. Kasuya, A. Sugisawa and M. Higuchi, 2002. The relation between single/double or repeated tea catechin ingestions and plasma antioxidant activity in humans. Eur. J. Clin. Nutr., 56: 1186-1193.
PubMedDirect Link - Kregel, K.C. and H.J. Zhang, 2007. An integrated view of oxidative stress in aging: Basic mechanisms, functional effects and pathological considerations. Am. J. Physiol. Regul. Integr. Comp. Physiol., 292: R18-R36.
CrossRefDirect Link - Lee, A.C., B.E. Fenster, H. Ito, K. Takeda and N.S. Bae, 1999. Ras proteins induce senescence by altering the intracellular levels of reactive oxygen species. J. Biol. Chem., 274: 7936-7940.
Direct Link - Lee, S., J.M. Lim, M.H. Jin, H.K. Park and E.J. Lee et al., 2006. Partially purified paeoniflorin exerts protective effects on UV-induced DNA damage and reduces facial wrinkles in human skin. J. Cosmet. Sci., 57: 57-64.
PubMed - Lin, S.J., P.A. Defossez and L. Guarente, 2000. Requirement of NAD and SIR2 for Life-span extension by calorie restriction in Saccharomyces cerevisiae. Science, 289: 2126-2128.
CrossRef - Meydani, M., 1992. Protective role of dietary vitamin E on oxidative stress in aging. Age, 15: 89-93.
CrossRef - Mirshafiey, A. and M. Mohsenzadegan, 2008. The role of reactive oxygen species in immunopathogenesis of rheumatoid arthritis. Iran J. Allergy Asthma Immunol., 7: 195-202.
PubMed - Mousavi, S., M. Mojtahedzadeh and M. Abdollahi, 2010. Place of iron chelators like desferrioxamine and deferasirox in management of hyperoxia-induced lung injury: A systematic review. Int. J. Pharmacol., 6: 326-337.
CrossRefDirect Link - Momtaz, S. and M. Abdollahi, 2010. An update on pharmacology of Satureja species; From antioxidant, antimicrobial, antidiabetes and anti-hyperlipidemic to reproductive stimulation. Int. J. Pharmacol., 6: 346-353.
CrossRefDirect Link - Nachbar, F. and H.C. Korting, 1995. The role of vitamin E in normal and damaged skin. J. Mol. Med. (Berl), 73: 7-17.
PubMed - Ohta, H., K. Matsumoto, M. Shimizu and H. Watanabe, 1994. Paeoniflorin attenuates learning impairment of aged rats in operant brightness discrimination task. Pharmacol. Biochem. Behav., 49: 213-217.
CrossRef - Oken, B.S., D.M. Storzbach and J.A. Kaye, 1998. The efficacy of ginkgo biloba on cognitive function in alzheimer disease. Arch. Neurol., 55: 1409-1415.
CrossRefPubMedDirect Link - Olshansky, S.J., L. Hayflick and B.A. Carnes, 2002. Position statement on human aging. J. Gerontol. A Biol. Sci. Med. Sci., 57: B292-B297.
PubMed - Ongusaha, P.P., T. Ouchi, K.T. Kim, E. Nytko and J.C. Kwak et al., 2003. BRCA1 shifts p53-mediated cellular outcomes towards irreversible growth arrest. Oncogene, 22: 3749-3758.
PubMed - Pantoja, C. and M. Serrano, 1999. Murine fibroblasts lacking p21 undergo senescence and are resistant to transformation by oncogenic Ras. Oncogene, 18: 4974-4982.
PubMed - Parrinello, S., E. Samper, A. Krtolica, J. Goldstein, S. Melov and J. Campisi, 2003. Oxygen sensitivity severely limits the replicative lifespan of murine fibroblasts. Nat. Cell. Biol., 5: 741-747.
PubMed - Peng, C., H.Y. Chan, Y. Huang, H. Yu and Z.Y. Chen, 2011. Apple polyphenols extend the mean lifespan of Drosophila melanogaster. J. Agric. Food Chem., 59: 2097-2106.
PubMed - Pryor, W.A., W. Stahl and C.L. Rock, 2000. Beta carotene: From biochemistry to clinical trials. Nutr. Rev., 58: 39-53.
CrossRefDirect Link - Rivers, J.K., 2008. The role of cosmeceuticals in antiaging therapy. Skin Ther. Lett., 13: 5-9.
PubMed - Roob, J.M., G. Khoshsorur, A. Tiran, J.H. Horina, H. Holzer and B.M. Winklhoffer-Roob, 2000. Vitamin E attenuates oxidative stress induced by intravenous iron in patients on hemodialysis. J. Am. Soc. Nephrol., 11: 539-549.
PubMed - Shukitt-Hale, B., V. Cheng and J.A. Joseph, 2009. Effects of blackberries on motor and cognitive function in aged rats. Nutr. Neurosci., 12: 135-140.
PubMed - Smith, J.V. and Y. Luo, 2004. Studies on molecular mechanisms of Ginkgo biloba extract. Applied Microbiol. Biotechnol., 64: 465-472.
CrossRef - Takahashi, A., N. Ohtani, K. Yamakoshi, S. Iida and H. Tahara et al., 2006. Mitogenic signalling and the p16INK4a-Rb pathway cooperate to enforce irreversible cellular senescence. Nat. Cell Biol., 8: 1291-1297.
PubMed - Terman, A. and U. Brunk, 2006. Oxidative stress, accumulation of biological Garbage and aging. Antioxid. Redox Signal., 8: 197-204.
PubMed - Unno, K., F. Takabayashi, T. Kishido and N. Oku, 2004. Suppressive effect of green tea catechins on morphologic and functional regression of the brain in aged mice with accelerated senescence (SAMP10). Exp. Gerontol., 39: 1027-1034.
CrossRef - Veskoukis, A.S., A.M. Tsatsakis and D. Kouretas, 2012. Dietary oxidative stress and antioxidant defense with an emphasis on plant extract administration. Cell Stress Chaperones, 17: 11-21.
CrossRefDirect Link - Werninghaus, K., R.M. Handjani and B.A. Gilchrest, 1991. Protective effect of alpha-tocopherol in carrier liposomes on ultraviolet-mediated human epidermal cell damage in vitro. Photodermatol Photoimmunol Photomed. 8: 236-242.
PubMed - Wick, G., 2002. Anti-aging medicine: Does it exist? A critical discussion of anti-aging health products. Exp. Gerontol., 37: 1137-1140.
CrossRef - Wu, Z., J.V. Smith, V. Paravasivam, P. Buiko, I. Khan, J.R. Cypser and Y. Luo, 2002. Ginkgo biloba extract EGb 761 increases stress resistance and extends life span of Caenorhabditis elegans. Cell. Mol. Biol., 48: 725-731.
PubMed - Xiang, L., K. Sun, J. Lu, Y. Weng, A. Taoka, Y. Sakagami and J. Qi, 2011. Anti-aging effects of phloridzin an apple polyphenol, on yeast via the SOD and Sir2 genes. Biosci Biotechnol Biochem., 75: 854-858.
PubMed - Zdanov, S., F. Debacq-Chainiaux, J. Remacle and O. Toussaint, 2006. Identification of p38MAPK-dependent genes with changed transcript abundance in H2O2- induced premature senescence of IMR-90 hTERT human fibroblasts. FEBS Lett., 580: 6455-6463.
PubMed - Schmitt-Schillig, S., S. Schaffer, C.C. Weber, G.P. Eckert and G.P. Muller, 2005. Flavonoids and the aging brain. J. Physiol. Pharmacol., 56: 23-36.
PubMedDirect Link - Afshari, M., B. Larijani, A. Rezaie, A. Mojtahedi and M.J. Zamani et al., 2004. Ineffectiveness of allopurinol in reduction of oxidative stress in diabetic patients; a randomized, double‐blind placebo‐controlled clinical trial. Biomed. Pharmacother., 58: 546-550.
CrossRefDirect Link - Ames, B.N., M.K. Shigenaga and T.M. Hagen, 1993. Oxidants, antioxidants and the degenerative diseases of aging. Proc. Nat. Acad. Sci. USA, 90: 7915-7922.
CrossRefDirect Link - Andersen, H.R., J.B. Nielsen, F. Nielsen and P. Grandjean, 1997. Antioxidative enzyme activities in human erythrocytes. Clin. Chem., 43: 562-568.
Direct Link - Bauza, E., C. Dal Farra, A. Berghi, G. Oberto, D. Peyronel and N. Domloge, 2002. Date palm kernel exhibits antiaging properties and significantly reduces skin wrinkles. Int. J. Tissue React., 24: 131-136.
PubMedDirect Link - Burdon, R.H., 1995. Superoxide and hydrogen peroxide in relation to mammalian cell proliferation. Free Radic. Biol. Med., 18: 775-794.
CrossRefPubMedDirect Link - Chow, C.K., 1991. Vitamin E and oxidative stress. Free Radic. Biol. Med., 11: 215-232.
CrossRefDirect Link - Droge, W., 2002. Free radicals in the physiological control of cell function. Phsyol. Rev., 82: 47-95.
CrossRefPubMedDirect Link - Harman, D., 1956. Aging: A theory based on free radical and radiation chemistry. J. Gerontol., 11: 298-300.
Direct Link - Hasani-Ranjbar, S., B. Larijani and M. Abdollahi, 2009. A systematic review of the potential herbal sources of future drugs effective in oxidant-related diseases. Inflamm. Allergy Drug Targets, 8: 2-10.
CrossRefPubMedDirect Link - Hasani-Ranjbar, S., N. Nayebi, B. Larijani and M. Abdollahi, 2010. A systematic review of the efficacy and safety of Teucrium species; from anti-oxidant to anti-diabetic effects. Int. J. Pharmacol., 6: 315-325.
CrossRef - Hasani-Ranjbar, S., H. Vahidi, S. Taslimi, N. Karimi, B. Larijani and M. Abdollahi, 2010. A systematic review on the efficacy of herbal medicines in the management of human drug-induced hyperprolactinemia; Potential sources for the development of novel drugs. Int. J. Pharmacol., 6: 691-695.
CrossRefDirect Link - Ho, Y.S., K.F. So and R.C.C. Chang, 2010. Anti-aging herbal medicine-How and why can they be used in aging-associated neurodegenerative diseases?. Ageing Res. Rev., 9: 354-362.
CrossRef - Inal, M.E., G. Kanbak and E. Sunal, 2001. Antioxidant enzyme activities and malondialdehyde levels related to aging. Clin. Chim. Acta, 305: 75-80.
CrossRef - Karami-Mohajeri, S. and M. Abdollahi, 2011. Toxic influence of organophosphate, carbamate and organochlorine pesticides on cellular metabolism of lipids, proteins and carbohydrates: A systematic review. Hum. Exp. Toxicol., 30: 1119-1140.
CrossRefDirect Link - Khan, S.G., S.K. Katiyar, R. Agarwal and H. Mukhata, 1992. Enhancement of antioxidant and phase II enzymes by oral feeding of green tea polyphenols in drinking water to SKH-1 hairless mice: Possible role in cancer prevention. Cancer Res., 52: 4050-4052.
Direct Link - Larijani, B., M. Salimi, N. Pourkhalili, A. Mohammadirad, M. Baeeri, A. Nili-Ahmadabadi and M. Abdollahi, 2011. Positive response of isolated rat pancreatic islets to IMOD; hopes for better transplant outcome and graft function. Asian J. Anim. Vet. Adv., 6: 1019-1025.
CrossRefDirect Link - Malekirad, A.A., N. Hosseini, M. Bayrami, T. Hashemi, K. Rahzani and M. Abdollahi, 2011. Benefit of lemon verbena in healthy subjects, targeting diseases associated with oxidative stress. Asian J. Anim. Vet. Adv., 6: 953-957.
CrossRefDirect Link - Manach, C., A. Scalbert, C. Morand, C. Remesy and L. Jimenez, 2004. Polyphenols: Food sources and bioavailability. J. Clin. Nutr., 79: 727-747.
CrossRefDirect Link - McCall, M.R. and B. Frei, 1999. Can antioxidant vitamins materially reduce oxidative damage in humans? Free Radic. Biol. Med., 26: 1034-1053.
CrossRefDirect Link - Mohammadi, M., S. Atashpour, N. Pourkhalili, A. Nili-Ahmadabadi and M. Baeeri et al., 2011. Comparative improvement in function of isolated rat langerhans islets by various phosphodiesterase 3, 4 and 5 inhibitors. Asian J. Anim. Vet. Adv., 6: 1233-1240.
CrossRef - Mohammadirad, A. and M. Abdollahi, 2011. A systematic review on oxidant/antioxidant imbalance in aluminum toxicity. Int. J. Pharmacol., 7: 12-21.
CrossRefDirect Link - Mohammadirad, A., H.R. Khorram-Khorshid, F. Gharibdoost and M. Abdollahi, 2011. Setarud (IMODTM) as a multiherbal drug with promising benefits in animal and human studies: A comprehensive review of biochemical and cellular evidences. Asian J. Anim. Vet. Adv., 6: 1185-1192.
CrossRefDirect Link - Monfared, S.S.M.S., B. Larijani and M. Abdollahi, 2009. Islet transplantation and antioxidant management: A systematic review. World J. Gastroenterol., 15: 1153-1161.
CrossRefPubMedDirect Link - Najafi, A., M. Mojtahedzadeh, A. Mahmoodpoor, M. Aghamohammadi and A. Ahmadi et al., 2009. Effect of N-acetylcysteine on microalbuminuria in patients with acute respiratory distress syndrome. Arch. Med. Sci., 5: 408-414.
Direct Link - Nunomura, A., R.J. Castellani, X. Zhu, P.I. Moreira, G. Perry and M.A. Smith, 2006. Involvement of oxidative stress in Alzheimer disease J. Neuropathol. Exp. Neurol., 65: 631-641.
CrossRefPubMedDirect Link - Perry, E.K., A.T. Pickering, W.W. Wang, P.J. Houghton and N.S. Perry, 1999. Medical plants and Alzheimers disease: From ethnobotany to phytotherapy. J. Pharmacy Pharmacol., 51: 527-534.
CrossRef - Prior, R.L. and G. Cao, 1999. In vivo total antioxidant capacity: Comparison of different analytical methods. Free Radic. Biol. Med., 27: 1173-1181.
CrossRefPubMedDirect Link - Radfar, M., B. Larijani, M. Hadjibabaie, B. Rajabipour, A. Mojtahedi and M. Abdollahi, 2005. Effects of pentoxifylline on oxidative stress and levels of EGF and NO in blood of diabetic type-2 patients: A randomized, double-blind placebo-controlled clinical trial. Biomed. Pharmacother., 59: 302-306.
Direct Link - Rahimi, R., S. Ghiasi, H. Azimi, S. Fakhari and M. Abdollahi, 2010. A review of the herbal phosphodiesterase inhibitors: Future perspective of new drugs. Cytokine, 49: 123-129.
CrossRefPubMedDirect Link - Rahimi, R., S. Mozaffari and M. Abdollahi, 2009. On the use of herbal medicines in management of inflammatory bowel diseases: A systematic review of animal and human studies. Dig. Dis. Sci., 54: 471-480.
CrossRefDirect Link - Rahimi, R., S. Nikfar, B. Larijani and M. Abdollahi, 2005. A review on the role of antioxidants in the management of diabetes and its complications. Biomed. Pharmacother., 59: 365-373.
PubMedDirect Link - Rastegarpanah, M., N. Omidzohour, H. Vahedi, R. Malekzadeh, F. Hashemian, T. Safarnavadeh and M. Abdollahi, 2011. Management of human ulcerative colitis by SaturexTM: A randomized controlled trial. Int. J. Pharmacol., 7: 516-521.
CrossRefDirect Link - Sangeetha, P., M. Balu, D. Haripriya and C. Panneerselvam, 2005. Age associated changes in erythrocyte membrane surface charge: Modulatory role of grape seed proanthocyanidins. Exp. Gerontol., 40: 820-828.
CrossRef - Serra, V., T. von Zglinicki, M. Lorenz and G. Saretzki, 2003. Extracellular superoxide dismutase is a major antioxidant in human fibroblasts and slows telomere shortening. J. Biol. Chem., 278: 6824-6830.
PubMedDirect Link - Shadnia, S., S. Ashrafivand, S. Mostafalou and M. Abdollahi, 2011. N-acetylcysteine a novel treatment for acute human organophosphate poisoning. Int. J. Pharmacol., 7: 732-735.
CrossRef - Soltaninejad, K. and M. Abdollahi, 2009. Current opinion on the science of organophosphate pesticides and toxic stress: A systematic review. Med. Sci. Monit., 15: RA75-RA90.
Direct Link - Zaveri, N.T., 2006. Green tea and its polyphenolic catechins: Medicinal uses in cancer and noncancer applications. Life Sci., 78: 2073-2080.
CrossRefPubMedDirect Link - Vijg, J., 2000. Somatic mutations and aging: A re-evaluation. Mutat. Res., 447: 117-135.
PubMedDirect Link - Beckman, K.B. and B.N. Ames, 1998. The free radical theory of aging matures. Physiol. Rev., 78: 547-581.
PubMed - Barger, J.L., T. Kayo, J.M. Vann, E.B. Arias and J. Wang et al., 2008. A low dose of dietary resveratrol partially mimics caloric restriction and retards aging parameters in mice. Plos One, Vol. 3.
CrossRef - Abdollahi, M., S. Mostafalou, S. Pournourmohammadi and S. Shadnia, 2004. Oxidative stress andcholinesterase inhibition in saliva and plasma of rats following subchronic exposure to malathion. Comparative Biochem. Physiol. Part C: Toxicol. Pharmacol., 137: 29-34.
CrossRef - Abdollahi, M., B. Larijani, R. Rahimi and P. Salari, 2005. Role of oxidative stress in osteoporosis. Therapy, 2: 787-796.
CrossRef - Shadnia, S., E. Azizi, R. Hosseini, S. Khoei and S. Fouladdel et al., 2005. Evaluation of oxidative stress and genotoxicity in organophosphorus insecticide formulators. Hum. Exp. Toxicol., 24: 439-445.
CrossRef - Kajbaf, F., M. Mojtahedzadeh and M. Abdollahi, 2007. Mechanisms underlying stress-induced hyperglycemia in critically ill patients. Therapy, 4: 97-106.
CrossRefDirect Link