
Shumaila Shahid
Department of Plant Protection, Faculty of Agricultural Sciences, Aligarh Muslim University, 202002 Aligarh, India
Mujeebur Rahman Khan
Department of Plant Protection, Faculty of Agricultural Sciences, Aligarh Muslim University, 202002 Aligarh, India
Plant Pathology Journal
Year: 2016 | Volume: 15 | Issue: 2 | Page No.: 27-39







ABSTRACT
Present study was carried out to examine the biological potential of different fungal and bacterial antagonists viz., Trichoderma harzianum, T. reesei, Aspergillus niger and Bacillus subtilis in management of root-rot disease caused by Macrophomina phaseolina and on the growth parameters of mungbean under pot condition. From the results, it was evident that the microbial antagonists significantly suppressed the root-rot disease (27-51%) and improved the growth and yield of mungbean. However, the root-rot fungus caused excessive rotting of roots and stunting of plant growth in the untreated plants. Among the tested biological control agents against M. phaseolina, T. harzianum was found to be the most effective against the fungus and resulted in 51% decrease in the root-rot severity and 31% increase in the yield, followed by B. subtilis. Root-rot infection significantly reduced functional root nodules by 58%. Biochemical components viz., total chlorophyll, total carotenoid and leghaemoglobin content was also decreased by 48, 38 and 25%, respectively, in the root-rot fungus infected plants. However, treatment with the biological control agents significantly increased the biochemical components as well as root nodulation in the order T. harzianum>B. subtilis>T. reesei>A. niger. Soil population of M. phaseolina in untreated pots increased exponentially but the increase was 30-66% less in the pots treated with biological control agents. The study has clearly demonstrated that these microbial antagonists specially, T. harzianum and B. subtilis significantly managed the root-rot disease as well as improved the plant growth and yield of mungbean.
PDF Abstract XML References Citation
Received: February 01, 2016;
Accepted: February 29, 2016;
Published: March 15, 2016
How to cite this article
Shumaila Shahid and Mujeebur Rahman Khan, 2016. Biological Control of Root-rot on Mungbean Plants Incited By Macrophomina phaseolina Through Microbial Antagonists. Plant Pathology Journal, 15: 27-39.
DOI: 10.3923/ppj.2016.27.39
URL: https://scialert.net/abstract/?doi=ppj.2016.27.39
DOI: 10.3923/ppj.2016.27.39
URL: https://scialert.net/abstract/?doi=ppj.2016.27.39
INTRODUCTION
Foods which provide high quality protein and are cheap in price are a demand for the poor living in developing countries like India where a large population is vegetarian (Saravanakumar et al., 2007). For this purpose, pulses are always the first preference for them due to protein rich diet, low cost and long time storage (Thirumaran and Seralathan, 1988). In India, the production of pulses is far below the requirement to meet even the minimum level of per capita consumption which requires that agricultural scientists should evolve the strategy to improve its production to meet the protein requirement of increasing population of the country. The productivity of pulses in India is considerably low when compared with the average global mean productivity of 496.4 kg ha–1 (FAOSTAT., 2014).
Mungbean or green gram, Vigna radiata (L.) Wilczek, an important pulse crop, is an excellent source of low cost and high quality protein (Taylor et al., 2005) contributes about 14% of the total protein of average diet of an Indian. It is rich in vitamin A, B, C, niacin and minerals such as calcium, phosphorus and potassium which are necessary for human body (Rattanawongsa, 1993). Since mungbean roots fix atmospheric nitrogen through symbiosis with nitrogen-fixing rhizobia, this crop is valuable both economically as well as nutritionally and is widely used in different cropping systems (Yaqub et al., 2010).
The low productivity and poor quality of mungbean in India can be attributed to several biotic and abiotic constraints of which diseases caused by fungi are of great importance (Khan and Khan, 2001). Of these, the root-rot disease caused by the soil and seed-borne fungus M. phaseolina (Tassi) Goid is a major limiting factor in the mungbean production (Raguchander et al., 1993). It is a serious disease of many crops, inflicting up to 100% yield losses in mungbean under dry and hot conditions. It is known to infect more than 500 plant species worldwide (Wyllie, 1993) and causes root-rot disease in several agronomically important crops. However, it is economically more important in the tropical and sub tropical areas of the world having arid climate (Wrather et al., 1997). The disease is enhanced by warm dry growing conditions and drought stress (Wrather et al., 1997; Dhingra and Sinclair, 1978). Mungbean is susceptible to M. phaseolina at various growth stages (Wheeler, 1975). All parts of plant i.e., root, stem, branches, petioles, leaves, pods and seeds are attacked by the fungus. Roots infected by M. phaseolina show necrotic lesions (Bouhot, 1967). At pre-emergence stage, the pathogen may inhibit seed germination and heavily infected plants die prematurely. Seed infection of Rhizoctonia bataticola (M. phaseolina) ranges from 2.2-15.7% which causes a reduction of 10.8% in grain yield and 12.3% in protein content of seed in mungbean (Kaushik and Chand, 1987). The fungus is soil as well as seed-borne in nature which helps it to survive under adverse climatic conditions and makes it difficult to manage.
Considering the importance of the disease, various management strategies have been adopted viz., cultural, physical, biological and chemical control methods. When the disease occurs in severe form, farmers mostly apply fungicides which do not prove much effective. Apart from the high costs, chemical control may have deleterious effects on environment and human. Moreover, several researchers have also pointed out that management of M. phaseolina through chemicals is not profitable nor advisable (Khalko et al., 2006) because there is greater variability among isolates of M. phaseolina and more chances of development of resistance in the population of fungal pathogens to most of the efficacious fungicides (Guizzardi et al., 1995). This has provided a strong impetus for consideration of use of ecofriendly methods in plant disease management (Hajieghrari et al., 2008). Hence, biological management offers an economic and ecologically viable approach towards disease control in the present agriculture as it is easier, safer to human beings, environment and non target organisms (Heungens and Parke, 2000).
Biological control using antagonistic microorganisms offers low cost ecofriendly technology to suppress the activity of phytopathogens (Yang et al., 2014). Various biocontrol agents e.g., Trichoderma spp. (Pan et al., 2013), Bacillus subtilis (Kumar et al., 2011) and Aspergillus niger (Khan and Anwar, 2007) have been evaluated for the management of root-rot pathogens. Trichoderma spp., are among the most effective mycoparasites in the management of soil-borne fungal pathogens as it has greater ability to survive under a wide range of temperatures (Pan and Bhagat, 2008). Bacillus spp., has excellent colonization capacity and versatility to protect plants from plant pathogenic fungi (Siddiqui et al., 2001; Zhao et al., 2014). The A. niger has also been proved to be a potential microbial antagonist against several plant diseases (Sen et al., 1995; Mondal et al., 2000).
The aim of the present study was to assess the effectiveness of microbial antagonists (Trichoderma harzianum, T. reesei, Aspergillus niger and Bacillus subtilis) on root-rot disease in mungbean in terms of disease severity, growth and biochemical parameters under pot culture condition.
MATERIALS AND METHODS
Isolation and identification of the pathogen: Macrophomina phaseolina was isolated from diseased mungbean plants showing symptoms of wilting and rotting collected from an infected farmer’s field in Aligarh. The infected samples were packed in sterilized poly bags and brought to the laboratory. The fungus was isolated from the infected roots using the method described by Dubey et al. (2009). Infected roots showing typical symptoms were washed under running tap water. Thereafter, the roots were surface sterilized with 0.1% mercuric chloride for a min and then washed three times with distilled water to remove the toxic sterilant. After drying on filter paper, the roots are cut into small pieces of 1 cm each and with the help of sterilized forcep, the pieces were transferred aseptically on Potato Dextrose Agar (PDA) in petri plates under a laminar flow. The plates were incubated at 25±2°C for 6-8 days. Thereafter, the mycelial growth of the fungus on the plates were observed and examined to confirm the association of M. phaseolina with the mungbean roots. The root-rot fungus, M. phaseolina was identified on the basis of cultural and morphological characters (Dhingra and Sinclair, 1978).
Mass culture of the pathogen: The root-rot fungus was mass cultured on sorghum seeds in an incubator at 25±2°C for 7 days. The sorghum seeds were soaked in 5% sucrose solution for 12 h. The sucrose provides supplimentary carbon for the growth of the pathogen. The conical flasks filled with the sorghum seeds (two third) were autoclaved at 15 kg cm‾2 pressure at 121°C for 15-20 min. Thereafter, the seeds were inoculated with 3 mm diameter mycelial discs of 5 days old culture of M. phaseolina (3 discs/flask) under laminar flow and incubated at 25±2°C for 7 days. During incubation period, the flasks were shaken daily manually for a few minutes in order to facilitate uniform colonization of the pathogen on the seeds. After the incubation, estimation of the pathogen population grown on sorghum seeds was done by using dilution plate method (Waksman, 1922). About 10 g fungus colonized seeds were macerated in 100 mL distilled water and serially diluted to 10–6 dilutions. About 0.3 mL suspension from the final dilution was spread over the solidified PDA in petri plates. The plates were incubated at 25±2°C for 5 days and Colony Forming Units (CFU) load of the fungus was determined by counting the colonies under a colony counter.
Isolation and mass culture of biocontrol agents: Four isolates of biocontrol agents (BCAs) of which three were fungal BCAs viz., T. harzianum, T. reesei and A. niger and one bacterial BCA viz., B. subtilis were isolated from the rhizospheric soil of mungbean. The biocontrol fungi were subcultured on PDA in petri plates and mass cultured on Potato Dextrose Broth (PDB) in 500 mL conical flasks and incubated at 25±2°C for 15 days. After incubation, the mycelial mat was harvested, filtered to separate spores from hyphae and ground with distilled water in an electric blender to make its homogenous suspension for uniform treatment of the seeds. The CFU load of the biocontrol fungi in the mycelial mat was determined using dilution plate method (Waksman, 1922). About 1 g mycelial mat was added with 10 mL Double Distilled Water (DDW) in a test tube and agitated for 5 min. About 1 mL of the suspension was then transferred to a test tube containing 9 mL DDW. This procedure was repeated until a dilution of 10–5 was achieved. 0.3 mL of suspension from the final dilution was pippeted and spread over solidified media in a petri plate with the help of a glass spreader. Three replicates were maintained for each biocontrol fungi.
The biocontrol bacteria was subcultured on nutrient agar in petri plates. It was mass cultured in the nutrient broth in conical flasks and incubated at 28±2°C for 2 days. After incubation the culture broth was centrifuged at 5000 rpm for 15 min. The pellet of bacterial cells was mixed in DDW. Similarly, the CFU load of the biocontrol bacteria in the suspension was then adjusted with DDW to achieve a dilution of 10–8 which was determined using the dilution plate method.
Seed treatment with biological control agents: Seeds of mungbean cv., T-44 were procured from an authorized seed dealer in Aligarh and were surface sterilized with HgCl2 solution (0.1%) for a min and then rinsed three times with sterililized water. Mungbean seeds were first applied with the sticker (5% molasses) so that the biocontrol agents get properly adhered to the seeds. The seeds were first treated with biological control agents (4 g kg–1 seeds) and then treated with commercial Rhizobium (20 g kg–1 seeds).
Plant culture: Earthen pots (25×25 cm dimension) were filled with 2 kg mixture of autoclaved loam soil and farmyard manure in the ratio of 3:1. About 10 g fungus colonized sorghum seeds were homogenized with 100 mL distilled water in an electric blender (1000 rpm). About 20 mL suspension containing 10 g fungus colonized sorghum seeds was mixed in 2 kg soil filled in the pots in order to achieve inoculums level at 5 g colonized seeds per kilogram soil. The inoculation was done 2 days prior to seed sowing. The BCA treated seeds were then sown in pots (5 seeds per pot) and after 10 days of sowing, thinning was done to retain only one plant per pot. The pots were arranged in a completely randomized block design consisting of 10 treatments and 3 replications. The plants were watered regularly. For estimation of root nodulation, three additional pots (replicates) were maintained for each treatment.
Nodulation i.e., number of functional and non-functional nodules/root system, nodular dry weight as well as leghaemoglobin content of nodules, total chlorophyll and carotenoid content of leaves was determined on 2 month old plants. At harvesting time i.e., 3.5 month after sowing, each plant was uprooted from the pot to determine the root-rot index on 0-5 scale (0: No rotting, 1: 1-20% rotting, 2: 21-40% rotting, 3: 41-60% rotting, 4: 61-80% rotting and 5: 81-100% rotting). The parameters of plant growth and yield of mungbean was determined at harvesting time and statistically analyzed. The pods were collected from three individual harvests at 2.5, 3.0 and 3.5 months age and were summed up to determine the yield.
Leghaemoglobin content in nodules: The leghaemoglobin content in fresh nodules was estimated by using the methodology of Sadasivam and Macickam (1992). Fresh nodules (200 mg) were mixed with of 0.1 M phosphate buffer (1.5 mL) and the mixture was macerated in an electric blender and then filtered through two layers of cheese cloth. The turbid reddish brown filtrate was centrifuged for 10-30 min at 10,000×g and 1.5 mL of pyridine reagent was added to 1.5 mL of extract. On mixing, the solution becomes greenish yellow because of the formation of haemochrome. The solution was equally divided into two test tubes. To one test tube, a few crystals of potassium hexacyanoferrate were added to oxidise the haemochrome and the reading of the mixture was taken at 539 nm on spectrophotometer, whereas to the other test tube a few crystals of sodium dithionate were added to reduce the haemochrome which was read at 556 nm after an interval of 2-5 min, against a reagent blank. The leghaemoglobin content (Lb) was calculated by using the following equation:
where, D is initial dilution.
Total chlorophyll and total carotenoid content (Leaf pigments): The chlorophyll content in fresh leaves was estimated following method worked out by Arnon (1949). About 1 g of finely cut fresh leaves was ground to a fine paste using a mortar and pestle after pouring 20 mL of 80% acetone. The mixture was centrifuged at 5000 rpm for 5 min. The supernatant was collected in 100 mL volumetric flask. The residue was washed three times using 80% acetone. Each washing was collected in the same volumetric flask and the volume was made up to the mark, using 80% acetone. The absorbance was read at 645 and 663 nm against the blank on spectrophotometer. The total chlorophyll content present in the extract was calculated by using following Arnon equation (Arnon, 1949):
For estimation of total carotenoid content, the absorbance was read at 480, 645 and 663 nm against the blank on spectrophotometer. The total carotenoid content present in the extract was calculated following the formula given by Hendry and Price (1993):
| A | = | Absorbance at specific wavelengths |
| V | = | Final volume of leaf extract in 80% acetone |
| W | = | Fresh weight of leaf tissue, used for extraction |
Soil population of M. phaseolina and biological control agents: Soil population of M. phaseolina and biological control agents in terms of Colony Forming Units (CFU) was determined at planting, 2, 3 and 3.5 months (harvesting time) using the dilution plate method (Waksman, 1922). Rhizosphere soil (10 g) was carefully collected from the root zone (at 1-2 cm depth) in order to determine population of the pathogen at planting time for other months populations, the rhizosphere soil was collected at 3-6 cm depth of each of the three pots. The soil was dried in shade and then sieved separately. The soil was transferred to a conical flask to which 100 double distilled water was added. The soil suspension was shaken for 5 min and 1 mL of the suspension was transferred to a test tube containing 9 mL of sterile double distilled water. The procedure was repeated until a dilution of 10–6 (fungi) and 10–8 (bacteria) was achieved. About 0.3 mL suspension from the final dilution was spread over solidified PDA (fungi) or NA (bacteria) under laminar flow. Three plates were maintained for each treatment and the plates were incubated at 25±2°C (for fungi) for 5 days and 30±2°C for 2 days (for bacteria). After incubation, the colonies of the pathogen and biological control agents were counted under a colony counter.
Statistical analysis: Observations taken from three replicates for each treatment were averaged to calculate means. Data was analyzed with the help of statistical software R (R Development Core Team, 2011) and Least Significant Difference (LSD) was calculated at p≤0.05 (Dospehov, 1984) for all the variables to compare individual treatments. Single factor analysis of variance (ANOVA) was performed for the data on root-rot severity and soil population. The data on plant growth, yield, nodulation and biochemical parameters were subjected to two factor ANOVA. The F-values were also calculated. Standard errors has been calculated and marked in figures.
RESULTS AND DISCUSSION
The present study revealed the antagonistic effects of various biological control agents against M. phaseolina. All the isolates has reduced the disease severity and have shown significantly beneficial effects over the growth attributes, leaf pigment contents, nodulation, leghaemoglobin content and yield of mungbean.
Disease symptoms and severity: From the above experiment, it was revealed that the inoculation of mungbean cv., T-44 with M. phaseolina showed its susceptibility to the pathogen in the form of stunted growth with sudden wilting and drying. The infected leaves became yellowish brown or chlorotic and papery in texture. Severely affected leaves have fallen off prematurely. Withering has been observed from seedling to maturing stage which may be the result of necrosis of roots. The pathogen restricted the growth of lateral roots. The tap root was dark brown and hard and lacked fine root hairs. On average, severity of the disease in terms of root-rot index was 4.5 on 0-5 scale (Fig. 1). The severity however, decreased to a great extent on seed treatment with biological control agents. Etebarian (2006) also reported significant disease severity due to M. phaseolina in melon which was effectively reduced in the presence of microbial antagonist. Of the different microbial antagonists used, T. harzianum showed most effective results in decreasing the root-rot severity (51%) in comparison to control. Same finding was found by Hussain et al. (1990) who observed that T. harzianum and Gliocladium virens were effective in controlling the infection of M. phaseolina in mungbean. Khan et al. (2011a) found that T. harzianum and T. virens effectively decreased the severity of botrytis gray mold disease and increased the yield of chickpea. Trichoderma spp., are opportunistic, avirulent plant symbionts which acts as antagonists of various plant pathogenic fungi (Ayed et al., 2006; Mohiddin et al., 2010; Reshu and Khan, 2012).
Next in effectiveness was B. subtilis, which also caused significant (p≤0.05) reduction in the root-rot (38%). The B. subtilis is highly promising for the control of phytopathogenic diseases among bacterial biological control agents (Choudhary and Johri, 2009; Hobley et al., 2013). The T. reesei was same in significance as B. subtilis and suppressed the disease severity by 41%. Javaid et al. (2014) also reported pronounced antagonistic behavior of T. reesei and T. harzianum under in vitro experiment resulting in 55-65% suppression in the mycelial growth of M. phaseolina. Trichoderma spp., caused colonization of lateral roots like mycorrhizae which may acts as a barrier for the invasion and colonization by the root-rot fungus (Guenoune et al., 2001). Treatment with A. niger was found to be least effective and reduced root-rot severity by 27% in comparison to control sets (p≤0.05, Fig. 1).
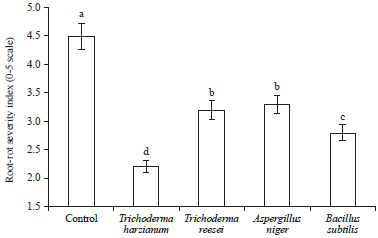 | |
| Fig. 1: | Effect of biological control agents on the severity of root-rot of mungbean caused by Macrophomina phaseolina. Error bars show standard error. Bars labelled by the same alphabets are not significantly different at p≤0.05 |
Seed treatment is an attractive and effective method to introduce antagonists into the environment of soil root since it gives protection to the seed from seed-borne as well as soil-borne pathogens subsequently decrease the disease severity and in addition, it enable the seed to germinate and become established as a healthy seedling (Chang and Kommedahl, 1968).
Plant growth: Application of biocontrol agents in uninoculated pots influenced the growth of mungbean plants, the effect, however, was non-significant when compared to control. Treatment with T. harzianum was found superior over other biocontrol agents and increased the shoot and root length by 4 and 5%, respectively, over uninoculated control. Trichoderma spp., produces plant growth promoting factors and secondary metabolites which may act as auxin like compound due to which enhancement of plant growth occurs (Vinale et al., 2008). The increase in shoot and root length by B. subtilis was 3 and 4% over control. The B. subtilis is considered as a Plant Growth Promoter Rhizobacteria (PGPR) (Turner and Backman, 1991).
Inoculation with M. phaseolina significantly suppressed shoot length (38%) and root length (32%) over uninoculated control (p≤0.05, Table 1). The M. phaseolina inhibits germination and growth of mungbean due to production of a toxin known as phaseolinon (Bhattacharya et al., 1994). Treatment with biocontrol agents checked the suppressive effects of root-rot fungus leading to 15-30% and 10-27% increase in shoot and root lengths of M. phaseolina infected plants. Maximum increase in root and shoot length was recorded in the plants treated with T. harzianum (27-30%). The B. subtilis showed the next maximum increase in shoot length (24%) but root length was equally increased with B. subtilis as with T. harzianum (27%). Bacillus subtilis has the capacity to colonize plant roots and does not leave enough roots for the invasion by pathogenic hyphae. Siddiqui et al. (2001) also reported the same finding that B. subtilis significantly suppressed root-rot infection in mungbean and enhanced plant growth and yield when used as seed dressing under greenhouse conditions. The A. niger was found to be least effective in increasing the root and shoot lengths (10-15%) over control (p≤0.05, Table 1). Aspergillus niger may cause suppression of plant pathogens through mycoparasitism, antibiosis and by stimulating plant defense reaction. Sen (2000) have also reported the antagonistic behaviour of A. niger and found that application of A. niger killed M. phaseolina, Rhizoctonia solani, Pythium spp. and Sclerotinia sclerotiorum.
Plant dry weight: The mungbean plants produced greatest dry matter when were applied with T. harzianum and resulted in 12-17% increase in the dry weight of shoot and root, respectively over uninoculated control. Trichoderma spp., actively colonize in soil (Akrami et al., 2009) and are antagonistic to M. phaseolina (Khan and Gupta, 1998). Bacillus subtilis, T. reesei and A. niger were similar in significance in increasing dry weight of shoot (4-10%) over control (p≤0.05, Table 1). The M. phaseolina inoculated plants showed 45 and 37% reduction in shoot and root dry weight over inoculated control. However, treatment with biocontrol agents checked the reduction significantly. Highest increase in shoot dry weight was recorded with T. harzianum (29%), followed by B. subtilis (21%). Trichoderma enhanced uptake of nutrients due to promotion of root growth (Harman et al., 2004). The mechanism of biocontrol by Bacillus activated was due to production of antibiotics, siderophores and hydrogen cyanide (Weller and Thomashow, 1993). Trichoderma reesei and A. niger were found equally effective in increasing the dry weight of root (13%) in comparison to inoculated control. The present study is in conformity with Mondal et al. (2000) who found that A. niger significantly increased root and shoot length and biomass of potato against M. phaseolina.
| Table 1: | Effect of biological control agents on the plant growth of mungbean grown in the soil inoculated or not inoculated with Macrophomina phaseolina |
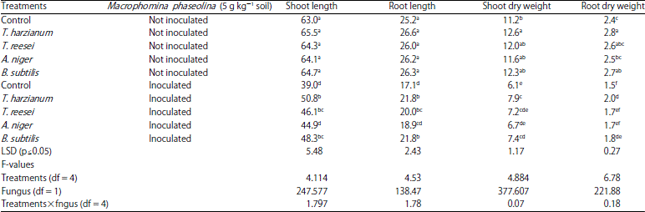 | |
| Each value is mean of three replicates, values within a column labelled by the same alphabets are not significantly different at p≤0.05 | |
Application of A. niger has been reported to improve growth and development of root, crop productivity, the uptake and use of nutrients and resistance to abiotic stresses (Sen, 2000). Trichoderma reesei has been reported to show effective antagonism against B. cinerea (El-Naggar et al., 2008).
Yield: Treatment with biocontrol agents in the non-infested pots enhanced the pod formation by 5-15% over control. Infestation with M. phaseolina caused a marked reduction in the pod formation (33%) over control (p≤0.05, Table 2). However, treatment with the biocontrol agents checked the suppressive effect of the fungus, leading to improvement in the number of pods per plant that varied with the treatment regime, T. harzianum being highly effective (32%). Trichoderma spp., produces various antibiotics e.g., trichodermin, viridin, gliotoxins, certain biologically active heat stable metabolites like ethyl acetate (Mohiddin et al., 2010) and cell wall degrading enzymes (Bruckner and Przybylski, 1984) which inhibits activity of soil-borne pathogens leading to the suppression of the disease and subsequently, increase in yield (Chet and Baker, 1981; Khan et al., 2011b).
Seed formation per pod was also significantly decreased in inoculated pots by 22%. The enhancement in the number of seeds per pod was 21% with T. harzianum and 19% with B. subtilis in comparison to inoculated control (p≤0.05, Table 2).
Non-infested pots treated with biocontrol agents significantly influenced the yield (weight of seeds per plant) of mungbean being maximum with T. harzianum (15%). Inoculation with M. phaseolina remarkedly reduced the yield by 36%. There was a significant improvement in the yield when biocontrol agents were incorporated to the pathogen infested pots (p≤0.05, Table 2). Maximum increase in yield was recorded in T. harzianum (31%) due to corresponding increase in the pod and seed formation with the biocontrol agent. Trichoderma spp., are considered as efficacious mycoparasites and they have strong capacity to control root-rot of mungbean and enhance its yield (Dubey et al., 2009). Next in effectiveness was B. subtilis and produced an increment of 24% in the yield. Bacillus spp., produces a large number of peptide antibiotics, hydrogen cyanide and low molecular weight volatile compounds which have specific activities against phytopathogenic fungi (Sarhan and Shehata, 2014; Chaves-Lopez et al., 2015). Abeysinghe (2009) reported that T. harzianum and B. subtilis showed significant control of damping-off disease of Solanum melongena and Capsicum annuum caused by R. solani. Minimum significant increase of 11% has been recorded with A. niger treatment. Vassilev et al. (2006) reported up to 47% increase in the plant growth production of tomato can after the addition of A. niger. Dawar et al. (2008) found that seed dressing with A. niger showed significant but lesser increase than T. harzianum in enhancing the plant height, weight and reducing the infection of root rot fungi in okra and sunflower. This is in conformity to the present study.
Nodulation, nodular dry weight and leghaemoglobin content in nodules: Nodulation on roots of mungbean was luxuriant in pots not inoculated with M. phaseolina. Infection with root-rot fungus however, caused a significant reduction in the number of functional (58%), whereas it increased the number of non-functional nodules by 45% (p≤0.05, Table 3). The M. phaseolina colonized the nodules and caused their rotting due to which there is a remarkable decrease in the functional nodules and increase in the non-functional nodules.
| Table 2: | Effect of biological control agents on the yield of mungbean grown in the soil inoculated or not inoculated with Macrophomina phaseolina |
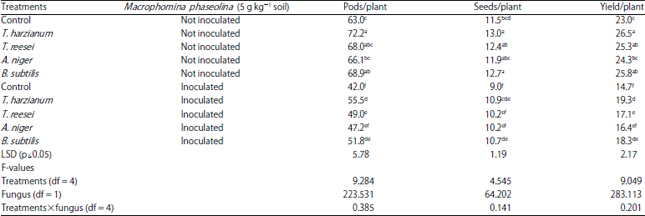 | |
| Each value is mean of three replicates, values within a column labelled by the same alphabets are not significantly different at p≤0.05 | |
Besides colonizing the nodules, M. phaseolina might have produced some metabolites which restricted the nodule formation. The present study is in conformity with Khan et al. (2001) who reported that M. phaseolina infection on roots caused rotting and emergence of lateral roots as a result of which nodule formation is reduced. Root nodulation in mungbean was greatly reduced due to infection by R. solani (Khan and Khan, 2001).
A significant (p≤0.05) increase in the number of functional (59%) and decrease in the number of non-functional nodules (32%) were recorded in the inoculated pots treated with T. harzianum (Table 3). Trichoderma improves nitrogen fixation by increasing nodulation on roots. Dry weight of nodules was found to be 43% lower in plants inoculated with the pathogen. A corresponding increase in the nodular dry weight was recorded with the biocontrol agents in uninoculated (4-27%) as well as inoculated (34-53%) pots (p≤0.05, Table 3). Choudhary and Sindhu (2015) reported that inoculation of B. subtilis enhanced the nodule number, nodule fresh weight and plant dry weight as compared to uninoculated control plants and also suppressed the root rot disease under pot house conditions. Since root-rot fungus caused rotting of lateral roots, therefore only limited roots were available for nodulation, hence lesser number of nodules were formed on the roots which can also be attributed to corresponding decrease in the nodular dry weight. However, due to suppression of the pathogen by the biological control agents, lesser root-rot and root decay occurred resulting in more availability of root hairs for Rhizobium colonization and hence, more root nodule formation.
It is clear from Table 3 that there was a marked decrease (26%) in the leghaemoglobin content of nodules in pathogen inoculated plants. Application of biocontrol agents led to increase in the leghaemoglobin content, however variations occurred in different treatments. A significant increase of 17 and 24% was recorded with T. harzianum in uninfested and infested pots due to more antagonism of the pathogen, whereas B. subtilis and T. reesei were significantly similar in causing the increase in uninfested (11-13%) and infested (20%) pots (p≤0.05, Table 3). Pathogen infection disturbs the functioning of nodules by altering its host nutrition and leghaemoglobin content gets decreased on infection due to improper functioning of nodules. Since leghaemoglobin regulates the supply of oxygen which affects the nitrogenase enzyme activity required for the reduction of atmospheric nitrogen to ammonia, therefore a decrease in the leghaemoglobin content due to pathogen infection would lead to a decrease in the nitrogen fixation. Altomare et al. (1999) reported that Trichoderma improved nitrogen fixation increased availability and absorption efficiency of solubilized micronutrients such as Fe, Cu and Mn etc., which subsequently, improved the plant growth.
Leaf pigments (Total chlorophyll and total carotenoid content): Data pertaining to leaf pigments depicted that the treatment with biocontrol agents in the uninoculated plants improved the total chlorophyll (11-21%) and total carotenoid (11-18%) content of the leaves. Infection with the pathogen reduced the total chlorophyll and total carotenoid content in the leaves by 48 and 38% (p≤0.05, Fig. 2 and 3). The T. harzianum and T. reesei were the most effective in increasing the total chlorophyll (40 and 28%) and total carotenoid content in the leaves of inoculated plants (35 and 26%).
| Table 3: | Effect of biological control agents on the root nodulation (number/root system), nodular dry weight and leghaemoglobin content of nodules of mungbean grown in the soil inoculated or not inoculated with Macrophomina phaseolina |
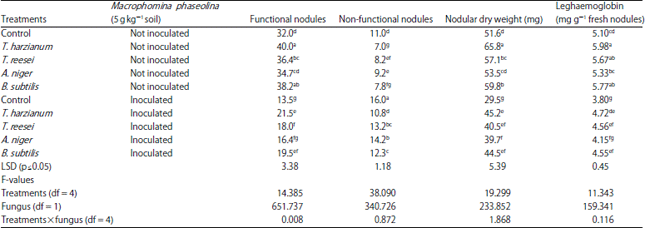 | |
| Each value is mean of three replicates, values within a column labelled by the same alphabets are not significantly different at p≤0.05 | |
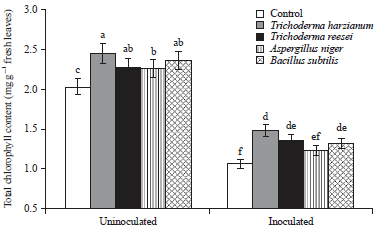 | |
| Fig. 2: | Effect of biological agents on the total chlorophyll content of mungbean grown in the soil inoculated or not uninoculated with Macrophomina phaseolina. Error bars show standard error. Bars labelled by the same alphabets are not significantly different at p≤0.05 |
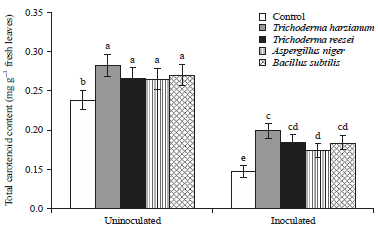 | |
| Fig. 3: | Effect of biological agents on the total carotenoid content of mungbean grown in the soil inoculated or not uninoculated with Macrophomina phaseolina. Error bars show standard error. Bars labelled by the same alphabets are not significantly different at p≤0.05 |
Trichoderma spp., increases availability and absorption efficiency of solubilized micronutrients such as Fe etc., which are the building blocks of chlorophyll. Uptake of nutrients improves due to application of Trichoderma which improves plant growth (Srivastava et al., 2006). Treatment with B. subtilis and A. niger showed comparatively less but significant effectiveness in enhancing total chlorophyll (16-24%) and total carotenoid (18-25%) content of the leaves (p≤0.05, Fig. 2 and 3). Chlorophyll content is significantly influenced by plant growth promoting rhizobacteria (El-Sherif et al., 2007).
Soil population of M. phaseolina and biocontrol agents: Soil population of root-rot fungus in the rhizosphere of plants increased with time and attained its peak in third month but declined at harvest. This may be due to non availability of host roots or hardening and rotting of the roots at the harvesting stage of the crop. The soil population increased from 2×105 to 10.5×105 CFUs g–1 soil up to 3 months which was 425% greater than the initial population and then at harvest, 7% decline was recorded in the fungus population from previous month population (p≤0.05, Fig. 4). Maximum decline in soil population was recorded with T. harzianum suppressing the build up of pathogen population up to 50% (first month) to 66% (harvesting time) over respective month control (p≤0.05, Fig. 4). Comparitively greater reduction by Trichoderma spp. might be due to better colonization and multiplication of the biocontrol fungus. The B. subtilis was next in effectiveness and caused a decrease of 54% in the M. phaseolina population till harvesting time. The A. niger was found least effective and caused a decline of 16-30% over control population.
Soil population of the biocontrol agents increased rapidly over initial population but the increase varied with the biocontrol agents (Fig. 5). The greatest increase in the soil population was recorded for T. harzianum (940%).
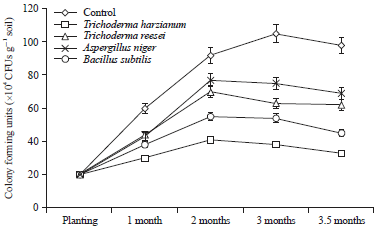 | |
| Fig. 4: | Effect of biological control agents on soil population of Macrophomina phaseolina in the rhizosphere of mungbean. Error bars show standard error |
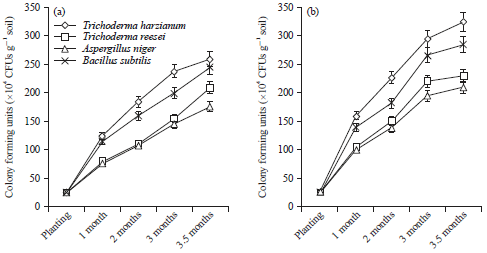 | |
| Fig. 5(a-b): | Soil population of biological control agents in the soil inoculated or not inoculated with Macrophomina phaseolina in the rhizosphere of mungbean. Error bars show standard error |
In the M. phaseolina infested pots, the population of biocontrol agents build up further over respective month population in the uninfested pots and caused an increase up to 300-540% in the first month, 456-808% in the second month, 680-1080% in the third month and 740-1200% at harvesting time in comparison to the initial (planting) population (p≤0.05, Fig. 5). Seed treatment with biocontrol agents provided an initial barrier against the pathogenic fungus through multiplication and colonization of seed by the biocontrol agents which in turn, would have restricted the entry, establishment and multiplication of root-rot fungus. Increase in the population of biocontrol agents in the inoculated pots can be attributed to the availability of host fungus on which the antagonists colonize and multiply rapidly (Jeffries and Young, 1994).
CONCLUSION
The results of the present pot study demonstrated the potentialities of microbial antagonists in the management of root-rot disease and improvement in the yield of mungbean which offers a promising and better alternative to costly and hazardous chemicals besides being safe for the environment as well to the consumers. The order of effectiveness of the tested biological control agents were T. harzianum> B. subtilis>T. reesei>A. niger. This is an ecofriendly as well as environmentally sustainable approach for the management of root-rot disease and can be used for field trials in naturally infected fields for the management of the disease in order to validate the present findings before its commercial use.
ACKNOWLEDGMENT
The financial assistance provided by the Department of Science and Technology (DST), New Delhi during the present study is greatly acknowledged. There is no conflict of interest between the authors. There is no financial interests or connections involved in the information during the manuscript.
REFERENCES
- Saravanakumar, D., S. Harish, M. Loganathan, R. Vivekananthan, L. Rajendran, T. Raguchander and R. Samiyappan, 2007. Rhizobacterial bioformulation for the effective management of Macrophomina root rot in mungbean. Arch. Phytopathol. Plant Protect., 40: 323-337.
CrossRefDirect Link - Taylor, R.S., D.B. Weaver, C. Wood and E. van Santen, 2005. Nitrogen application increases yield and early dry matter accumulation in late-planted soybean. Crop Sci., 45: 854-858.
CrossRefDirect Link - Yaqub, M., T. Mahmood, M. Akhtar, M.M. Iqbal and S. Ali, 2010. Induction of mungbean [Vigna radiata (L.) Wilczek] as a grain legume in the annual rice-wheat double cropping system. Pak. J. Bot., 42: 3125-3135.
Direct Link - Khan, N. and M.R. Khan, 2001. Screening of mungbean cultivars against increasing inoculum levels of Rhizoctonia solani. Tests Agrochem. Cultivars, 22: 36-37.
Direct Link - Raguchander, T., R. Samiappan and G. Arjunan, 1993. Biocontrol of Macrophomina root rot of mungbean. Indian Phytopathol., 46: 379-382.
Direct Link - Wrather, J.A., T.R. Anderson, D.M. Arsyad, J. Gai and L.D. Ploper et al., 1997. Soybean disease loss estimates for the top 10 soybean producing countries in 1994. Plant Dis., 81: 107-110.
CrossRefDirect Link - Dhingra, O.D. and J.B. Sinclair, 1978. Biology and Pathology of Macrophomina phaseolina. Imprensa Universitaria, Universidade Federal de Vicosa, Vicosa, Brazil, Pages: 166.
Direct Link - Kaushik, C.D. and J.N. Chand, 1987. Seedborne nature of Rhizoctonia bataticola causing leaf blight of mung bean. Indian J. Mycol. Plant Pathol., 17: 154-157.
Direct Link - Khalko, S., S. Pan and S. Bhagat, 2006. Development of tolerance and cross-tolerance in Macrophomina phaseolina against fungicides. Indian J. Plant Protect., 34: 233-235.
Direct Link - Guizzardi, M., D.R.L. Caccioni and G.C. Pratella, 1995. Resistance monitoring of Monilinia laxa (Aderh. et Ruhl.) honey to benzimidazoles and dicarboximides in postharvest stage. J. Plant Dis. Protect., 102: 86-90.
Direct Link - Hajieghrari, B., M. Torabi-Giglou, M.R. Mohammadi and M. Davari, 2008. Biological potantial of some Iranian Trichoderma isolates in the control of soil borne plant pathogenic fungi. Afr. J. Biotechnol., 7: 967-972.
Direct Link - Heungens, K. and J.L. Parke, 2000. Zoospore homing and infection events: Effects of the biocontrol bacterium Burkholderia cepacia AMMDR1 on two oomycete pathogens of pea (Pisum sativum L.). Applied Environ. Microbiol., 66: 5192-5200.
CrossRefDirect Link - Yang, Q.Y., K. Jia, W.Y. Geng, R.J. Guo and S.D. Li, 2014. Management of cucumber wilt disease by Bacillus subtilis B006 through suppression of Fusarium oxysporum f. sp. cucumerinum in rhizosphere. Plant Pathol. J., 13: 160-166.
CrossRefDirect Link - Pan, S., R. Mukherji and S. Bhagat, 2013. Evaluation of Trichoderma spp. against soil borne plant pathogens. Ann. Plant Protect. Sci., 21: 197-198.
Direct Link - Khan, M.R. and A.M. Anwer, 2007. Molecular and biochemical characterization of soil isolates of Aspergillus niger aggregate and an assessment of their antagonism against Rhizoctonia solani. Phytopathologia Mediterranea, 46: 304-315.
Direct Link - Pan, S. and S. Bhagat, 2008. Characterization of antagonistic potential of Trichoderma spp. against some soil borne plant pathogens. J. Biol. Control, 22: 43-49.
Direct Link - Siddiqui, I.A., S. Ehetshamul-Haque and S.S. Shaukat, 2001. Use of rhizobacteria in the control of root rot-root knot disease complex of mungbean. J. Phytopathol., 149: 337-346.
CrossRefDirect Link - Zhao, P., C. Quan, Y. Wang, J. Wang and S. Fan, 2014. Bacillus amyloliquefaciens Q-426 as a potential biocontrol agent against Fusarium oxysporum f. sp. spinaciae. J. Basic Microbiol., 54: 448-456.
CrossRefDirect Link - Sen, B., K. Mukherjee, C. Chattopadhyay, A.K. Patibanda and J. Sharma, 1995. Aspergillus niger, a potential biocontrol agent for soil-borne plant pathogens. Proceeding of the Global Conference on Advances in Research on Plant Conference and their Management Society of Mycology and Plant Pathology, February 12-17, 1995, Udaipur, pp: 161.
- Mondal, G., P. Dureja and B. Sen, 2000. Fungal metabolites from Aspergillus niger AN27 related to plant growth promotion. Indian J. Exp. Biol., 38: 84-87.
PubMedDirect Link - Dubey, S.C., R. Bhavani and B. Singh, 2009. Development of Pusa 5SD for seed dressing and Pusa Biopellet 10G for soil application formulations of Trichoderma harzianum and their evaluation for integrated management of dry root rot of mungbean (Vigna radiata). Biol. Control, 50: 231-242.
CrossRefDirect Link - Waksman, S.A., 1922. A method for counting the number of fungi in the soil. J. Bacteriol., 7: 339-341.
Direct Link - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Etebarian, H.R., 2006. Evaluation of Streptomyces strains for biological control of charcoal stem rot of Melon caused by Macrophomina phaseolina. Plant Pathol. J., 5: 83-87.
CrossRefDirect Link - Hussain, S., A. Ghaffar and M. Aslam, 1990. Biological control of Macrophomina phaseolina charcoal rot of sunflower and mung bean. J. Phytopathol., 130: 157-160.
CrossRefDirect Link - Khan, M.R., M.A. Anwer and S. Shahid, 2011. Management of gray mold of chickpea, Botrytis cinerea with bacterial and fungal biopesticides using different modes of inoculation and application. Biol. Control, 57: 13-23.
CrossRefDirect Link - Ayed, F., M. Daami-Remadi, F. Jabnoun-Khiareddine and M. El Mahjoub, 2006. Potato vascular Fusarium wilt in Tunisia: Incidence and biocontrol by Trichoderma sp. Plant Pathol. J., 5: 92-98.
CrossRefDirect Link - Mohiddin, F.A., M.R. Khan, S.M. Khan and B.H. Bhat, 2010. Why Trichoderma is considered super hero (super fungus) against the evil parasites? Plant Pathol. J., 9: 92-102.
CrossRefDirect Link - Reshu and M.M. Khan, 2012. Role of different microbial-origin bioactive antifungal compounds against Alternaria spp. causing leaf blight of mustard. Plant Pathol. J., 11: 1-9.
CrossRefDirect Link - Choudhary, D.K. and B.N. Johri, 2009. Interactions of Bacillus spp. and plants-with special reference to Induced Systemic Resistance (ISR). Microbiol. Res., 164: 493-513.
CrossRefPubMedDirect Link - Hobley, L., A. Ostrowski, F.V. Rao, K.M. Bromley and M. Porter et al., 2013. BslA is a self-assembling bacterial hydrophobin that coats the Bacillus subtilis biofilm. Proc. Nat. Acad. Sci. usa., 110: 13600-13605.
CrossRefDirect Link - Javaid, A., L. Afzal, A. Bashir and A. Shoaib, 2014. In vitro screening of Trichoderma species against Macrophomina phaseolina and Fusarium oxysporum f. sp. lycopersici. Pak. J. Phytopathol., 26: 39-43.
Direct Link - Guenoune, D., S. Galili, D.A. Phillips, H. Volpin, I. Chet, Y. Okon and Y. Kapulnik, 2001. The defense response elicited by the pathogen Rhizoctonia solani is suppressed by colonization of the AM-Fungus Glomus intraradices. Plant Sci., 160: 925-932.
CrossRefPubMedDirect Link - Vinale, F., K. Sivasithamparam, E.L. Ghisalberti, R. Marra and M.J. Barbetti et al., 2008. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol., 72: 80-86.
CrossRefDirect Link - Turner, J.T. and P.A. Backman, 1991. Factors relating to peanut yield increases after seed treatment with Bacillus subtilis. Plant Dis., 75: 347-353.
CrossRefDirect Link - Bhattacharya, D., T.K. Dhar, K.A.I. Siddiqui and E. Ali, 1994. Inhibition of seed germination by Macrophomina phaseolina is related to phaseolinone production. J. Applied Bacteriol., 77: 129-133.
CrossRefDirect Link - Akrami, M., A.S. Ibrahimov, D.M. Zafari and E. Valizadeh, 2009. Control Fusarium rot of bean by combination of by Trichoderma harzianum and Trichoderma asperellum in greenhouse condition. Agric. J., 4: 121-123.
Direct Link - Khan, M.R. and J. Gupta, 1998. Antagonistic efficacy of Trichoderma species against Macrophomina phaseolina on eggplant. J. Plant Dis. Protect., 105: 387-393.
Direct Link - Harman, G.E., C.R. Howell, A. Viterbo, I. Chet and M. Lorito, 2004. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol., 2: 43-56.
CrossRefPubMedDirect Link - Weller, D.M. and L.S. Thomashow, 1993. Use of rhizobacteria for biocontrol. Curr. Opin. Biotechnol., 4: 306-311.
CrossRefDirect Link - El-Naggar, M., G.J. Kovics and E. Sandor L. Irinyi, 2008. Mycoparasitism and antagonistic efficiency of Trichoderma reesei against Botrytis spp. Contribuţii Botanice, 43: 141-147.
Direct Link - Bruckner, H. and M. Przybylski, 1984. Isolation and structural characterization of polypeptide antibiotics of the peptaibol class by high-performance liquid chromatography with field desorption and fast atom bombardment mass spectrometry. J. Chromatogr. A, 296: 263-275.
CrossRefDirect Link - Chet, I. and R.R. Baker, 1981. Isolation and biocontrol potential of Trichoderma hamatum from soil naturally suppressive to Rhizoctonia solani. Phytopathology, 71: 286-290.
CrossRefDirect Link - Khan, M.R., S. Majid, F.A. Mohiddin and N. Khan, 2011. A new bioprocess to produce low cost powder formulations of biocontrol bacteria and fungi to control fusarial wilt and root-knot nematode of pulses. Biol. Control, 57: 130-140.
CrossRefDirect Link - Sarhan, E.A.D. and H.S. Shehata, 2014. Potential plant growth-promoting activity of Pseudomonas spp. and Bacillus spp. as biocontrol agents against damping-off in alfalfa. Plant Pathol. J., 13: 8-17.
CrossRefDirect Link - Chaves‐Lopez, C., A. Serio, A. Gianotti, G. Sacchetti and M. Ndagijimana et al., 2015. Diversity of food-borne Bacillus volatile compounds and influence on fungal growth. J. Applied Microbiol., 119: 487-499.
CrossRefDirect Link - Abeysinghe, S., 2009. Effect of combined use of Bacillus subtilis CA32 and Trichoderma harzianum RU01 on biological control of Rhizoctonia solani on Solanum melongena and Capsicum annuum. Plant Pathol. J., 8: 9-16.
CrossRefDirect Link - Vassilev, N., M. Vassileva and I. Nikolaeva, 2006. Simultaneous P-solubilizing and biocontrol activity of microorganisms: Potentials and future trends. Applied Microbiol. Biotechnol., 71: 137-144.
CrossRefDirect Link - Dawar, S., S. Hayat, M. Anis and M.J. Zaki, 2008. Effect of seed coating material in the efficacy of microbial antagonists for the control of root rot fungi on okra and sunflower. Pak. J. Bot., 40: 1269-1278.
Direct Link - Khan, M.R., N. Khan and S.M. Khan, 2001. Evaluation of agricultural materials as substrate for mass culture of fungal biocontrol agents of fusarial wilt and root-knot nematode diseases. Tests Agrochem. Cultivars, 22: 50-51.
Direct Link - Choudhary, S.R. and S.S. Sindhu, 2015. Suppression of Rhizoctonia solani root rot disease of clusterbean (Cyamopsis tetragonoloba) and plant growth promotion by rhizosphere bacteria. Plant Pathol. J., 14: 48-57.
CrossRefDirect Link - Altomare, C., W.A. Norvell, T. Bjbrkman and G.E. Harman, 1999. Solubilization of phosphates and micronutrients by the plant-growth-promoting and biocontrol fungus Trichoderma harzianum Rifai 1295-22. Applied Environ. Microbiol., 65: 2926-2933.
Direct Link - Srivastava, S.N., V. Singh and S.K. Awasthi, 2006. Trichoderma induced improvement in growth, yield and quality of sugarcane. SugarTech, 8: 166-169.
CrossRefDirect Link - El-Sherif, A.G., A.R. Refaei, M.E. El-Nagar and H.M.M. Salem, 2007. Integrated Management of Meloidogyne incognita infecting eggplant by certain organic amendments, Bacillus thuringiensis and oxamyl with reference to N P K and Total chlorophyll status. Plant Pathol. J., 6: 147-152.
CrossRefDirect Link