
Sita Ram Choudhary
Department of Microbiology, College of Basic Sciences and Humanities, CCS Haryana Agricultural University, Hisar, 125 004, India
Satyavir Singh Sindhu
Department of Microbiology, College of Basic Sciences and Humanities, CCS Haryana Agricultural University, Hisar, 125 004, India
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 2 | Page No.: 48-57







ABSTRACT
Clusterbean (Cyamopsis tetragonoloba (L.) Taub) is a kharif legume crop grown under arid zone. Root rot is the major disease in clusterbean caused by Rhizoctonia solani during rainy season and may result upto 21-60% losses at pre-and post-emergence stages. Rhizobacterial isolates were tested in this study for use as biological control agent for suppression of the root rot disease and to minimize the use of fungicides for disease control. Fifty five rhizobacterial isolates obtained from clusterbean rhizosphere soil were screened for antagonistic interactions against Rhizoctonia solani on PDA medium plates. Rhizobacterial isolates HCS2, HCS4, HCS30, HCS36, HCS43 and HFS12 showed significant antagonistic activity and inhibited the growth of fungi on PDA medium plates. Forty percent of these rhizobacterial isolates utilized ACC on minimal medium plates and five rhizobacterial isolates HCS16, HCS35, HCS42, HCS43 and HFS5 showed significant growth on ACC supplemented plates. Inoculation of Bradyrhizobium isolate GSA11 with Pseudomonas isolate HCS36/Bacillus isolate HCS43 and R. solani formed maximum 47 nodules plant-1 and increased shoot dry weight by 306.3 and 281.5%, respectively as compared to uninoculated control at 30 days of plant growth under pot house conditions. At 60 days of plant growth, coinoculation of Rhizobium isolate GSA110 with Bacillus isolate HCS43 and R. solani formed 50 nodules plant-1 and caused 108.9% increase in shoot dry weight in comparison to uninoculated control. Maximum 140.26% increase in shoot dry weight, nodule number (54 nodules plant-1) and nodule weight (346.6 mg plant-1) was observed by coinoculation of Pseudomonas isolate HCS36 with Rhizobium strain GSA110 and fungus R. solani. Coinoculation of Bacillus isolate HCS43 and Pseudomonas isolate HCS36 with Bradyrhizobium/Rhizobium isolates also showed 66.7 and 83.7% disease control. Thus, Bacillus isolate HCS43 and Pseudomonas isolate HCS36 could be further tested for disease control and plant growth stimulation under field conditions.
PDF Abstract XML References Citation
Received: March 24, 2015;
Accepted: May 25, 2015;
Published: June 11, 2015
How to cite this article
Sita Ram Choudhary and Satyavir Singh Sindhu, 2015. Suppression of Rhizoctonia solani Root Rot Disease of Clusterbean (Cyamopsis tetragonoloba) and Plant Growth Promotion by Rhizosphere Bacteria. Plant Pathology Journal, 14: 48-57.
DOI: 10.3923/ppj.2015.48.57
URL: https://scialert.net/abstract/?doi=ppj.2015.48.57
DOI: 10.3923/ppj.2015.48.57
URL: https://scialert.net/abstract/?doi=ppj.2015.48.57
INTRODUCTION
Pathogenic microorganisms cause various plant diseases that usually weaken or destroy plant tissues and reduce crop yields varying from 25-100% (Frisvad and Samson, 1991). Root diseases are estimated to cause 10-15% yield losses annually in the world (Bajoria et al., 2008). Rhizoctonia solani Kuhn (teleomorph: Thanatephorus cucumeris (A.B. Frank). Donk is an ecologically diverse soilborne fungus that causes root rot disease on clusterbean plants. Fungicide bavistin sprays are usually done for controlling the disease. But indiscriminate use of agrochemicals for disease and pest control has resulted into considerable pollution of soil, water and air. Moreover, wide spread use of agrochemicals also have undesirable effects on non-target organisms and possible carcinogenicity effects. Thus, their extensive use is environmentally unsafe and also uneconomical. Therefore, it is imperative to develop some alternate strategies for controlling plant diseases. Biological control using antagonistic microorganisms offers a low cost ecofriendly technology that reduces the number and activity of plant pathogens (Glick et al., 1999; Sindhu et al., 2009; Yang et al., 2014).
Rhizosphere bacteria (rhizobacteria) suppress/control the plant diseases by various mechanisms viz., production of antibiotics (Keel et al., 1992; Saraf et al., 2014), production of hydrolytic enzymes (Sindhu and Dadarwal, 2001), hydrocyanic acid (Sarhan and Shehata, 2014), stimulation of phytoalexins or flavonoid-like compounds in roots (Goel et al., 2001) or by production of siderophores, which chelate metal cations rendering them unavailable for pathogenic forms (Raaijmakers et al., 1995; Sahu and Sindhu, 2011). Certain microbial strains protect the plants against pathogens through induced systemic resistance (Kaiser and Hannan, 1989). Some rhizosphere bacteria possess the enzyme ACC (1-aminocyclopropane-1-carboxylate) deaminase that reduces the level of stress hormone ethylene production. Enzyme ACC deaminase has been reported in many soil microorganisms (Khandelwal and Sindhu, 2012; Glick, 2014). These ACC deaminase-containing bacterial strains were also found more effective biocontrol strains (Glick, 2004; Wang et al., 2000). Thus, use of biocontrol agents isolated from plants and soils holds a great promise to establish them in the rhizosphere to control various plant diseases without disrupting the ecological balance (Weller, 2007).
A major group of rhizobacteria with biological control potential are the Pseudomonas and Bacillus strains (Sundaramoorthy and Balabaskar, 2013; Sarhan and Shehata, 2014). These bacteria are ubiquitous in agricultural soils and possess many traits that make them well suited as biocontrol and growth-promoting agents (Sindhu et al., 2014). Saikia et al. (2004) screened 54 fluorescent Pseudomonas isolates obtained from broad bean rhizosphere for antagonism against Macrophomina phaseolina and R. solani and reported that Pseudomonas aeruginosa strain RsB29 caused suppression of Fusarium wilt and charcoal rot of chickpea, and promoted the plant growth of broad bean. Ramette et al. (2006) found that Pseudomonas populations growing in the rhizosphere soil of tobacco produced the biocontrol compounds viz., 2,4-diacetylphloroglucinol and hydrogen cyanide which were suppressive to root rot disease. Pseudomonas fluorescens strain CHA0 has been found to produce several secondary metabolites, notably HCN, 2,4-diacetylphloroglucinol, pyoluteorin and indole acetic acid (Keel et al., 1992). The combined application of P. fluorescens and B. subtilis exhibited highest reduction of tomato wilt disease and increased the dry weight of tomato plants up to 27% in comparison to the non-bacterized control (Sundaramoorthy and Balabaskar, 2013).
Clusterbean (Cyamopsis tetragonoloba (L.) Taub) is a kharif legume crop grown under arid zone in India (Singh et al., 2001). It is primarily grown for seed, animal feed, fodder, vegetable and green manuring purposes. Clusterbean is a rich source of high quality galatomannan gum and protein rich (40-50%) guar meal as animal feed. Seed gum is used in various industries such as textiles, paper, cosmetics, explosives and food processing. Besides the gum preparation, clusterbean is emerging as a potential source of vegetable protein for human beings. However, the root rot disease of clusterbean during rainy season may result up to 21-60% plant loss at pre-and post-emergence stages (Bajoria et al., 2008). Moreover, the information available on the antagonistic effect of rhizobacteria against R. solani is very scanty. Therefore, attempts were made in this study to develop biological control agents against root rot causing fungi R. solani.
MATERIALS AND METHODS
Isolation of bacteria from rhizosphere soil: Rhizosphere soil samples were collected from different fields of clusterbean grown in CCS Haryana Agricultural University, Hisar farm at 45 and 60 days of plant growth. The serial dilutions of the composite rhizosphere soil samples (up to 10‾4) were plated on King’s B agar medium. Pseudomonas and Bacillus colonies were selected based on morphological and pigment production characteristics after 3 days of incubation at 28±2°C. Purified colonies of bacteria were transferred on Luria Bertani (LB) agar medium slopes. Rhizobium/Bradyrhizobium strains were obtained from Department of Microbiology, CCS H.A.U., Hisar. Phytopathogenic fungus R. solani was obtained from the Department of Plant Pathology, CCS H.A.U., Hisar. Liquid cultures of all the isolates were preserved in 50% glycerol at -20°C.
Screening of rhizobacterial isolates for utilization of ACC: The medium plates were prepared with minimal medium (Dworkin and Foster, 1958) supplemented with 3 mM ACC (Penrose and Glick, 2003) or ammonium sulphate (2 g L-1). A loopful of 48 h old growth of Pseudomonas or Bacillus culture was spotted on the medium plates and incubated at 28±2°C for 2-5 days. The growth of different bacterial isolates on ACC supplemented medium plates was recorded (Khandelwal and Sindhu, 2012). The cultures showing good growth on ACC supplemented medium plates and capable of utilizing ACC as nitrogen source, were scored as ACC+.
Growth inhibition of Rhizoctonia solani by rhizobacterial isolates: The antagonistic interactions of rhizobacterial isolates with phytopathogenic fungus R. solani were studied by the spot test method on Potato Dextrose Agar (PDA) medium plates (Sindhu et al., 1999). Rhizoctonia solani was grown on PDA slants for 4 days and spore suspension was harvested in 3 mL sterilized water. Fungal spore suspension (3.0 mL) was added into sterilized PDA medium, mixed uniformly and plated. Growth suspension (5 μL) of 48 h old rhizobacterial cultures was spotted on spore suspension-containing plates. The inhibition of growth of R. solani by the spotted rhizobacterial isolates was recorded after 4 days of incubation at 28±2°C.
Coinoculation studies of selected rhizobacterial isolates with Bradyrhizobium/Rhizobium for nodulation and plant growth: Three rhizobacterial isolates i.e., HCS5 (control), HCS36 and HCS43 (with antagonistic activity) were used for coinoculation with symbiotically effective Bradyrhizobium/Rhizobium strains in clusterbean variety HG563 (susceptible to root rot disease). The earthern pots of 10 kg capacity were filled with sandy loam soil and river sand mixed in 70:30 ratio. Pseudomonas and Bacillus strains were grown on LB medium for 2 days and Bradyrhizobium/Rhizobium strains were grown on YEMA medium slopes for 7 days. The growth suspension of each Pseudomonas/Bacillus and Bradyrhizobium/Rhizobium cultures was made in 5 mL of sterilized water.
Seeds of clusterbean were inoculated either alone (with 10 mL of growth suspension) or as biomix (obtained from mixing of 5 mL growth of each bacteria in 1:1 ratio, v/v). The viable count in the broth was kept 108-109 cells mL-1 and 10 g seeds were inoculated with 10 mL of bacterial growth suspension and having 107-108 cells mL-1 of growth suspension. There were 20 treatments in this experiment and each treatment had three replications. Growth of 4 days-old R. solani was harvested from PDA plates and fungal growth suspension was prepared in sterilized saline water. Fungal growth suspension (100 mL) was mixed in the 10 kg soil: sand mixture in earthern pots with treatments T7, T10, T11, T14, T15, T18, T19 and T20. The growth suspension of fungus was inoculated on the roots of clusterbean plants in the R. solani treatments only. Uninoculated seeds were sown as control. The plants were grown in the pot house under day light conditions during the month of May-June 2013. Sloger’s nutrient solution was added in the pots as and when required (Sloger, 1969). The plants were uprooted at 30 and 60 days of plant growth and observations were taken for nodule number, nodule fresh weight, plant dry weight and disease index. After washing with tap water, nodules were detached from the roots and dried in the folds of filter paper. The nodules were counted and weighed. Shoot portions of the plants were dried in oven at 90°C for 24 h and weighed.
Disease index and reduction in disease: On the basis of symptoms observed percent disease index, percent final stand and percent disease control were calculated by following equation:
![]()
![]()
Disease control and disease incidence were recorded after 30 and 60 days of sowing. It was calculated on the average of six plants grown per pot.
Statistical analysis: Completely Randomized Design (CRD) was used for experimental data analysis. All determinations were carried out in triplicate and data represented are average values of three replications. Standard Error of Means± (SEM) values were calculated to determine the significant differences between treatment means. The C.D. and C.V. values represent coefficient of deviation and coefficient of variation, respectively.
RESULTS
Fifty five bacterial isolates representing Pseudomonas and Bacillus (based on morphological and pigment production characteristics) were selected from rhizosphere soil at 45 and 60 days of culsterbean growth. Bacterial counts in the rhizosphere soil ranged from 1.7×18.4×105 colony forming units (CFU) g-1 soil at 45 days of plant growth. At 60 days of plant growth, bacterial counts increased and ranged from 19.2-76.8×105 CFU g-1 soil. Similarly, Baig et al. (2002) isolated 105 bacteria from rhizosphere and rhizoplane of groundnut. Out of these, 67% isolates were from the rhizosphere and 33% were from the rhizoplane. Pseudomonas was found as the most predominant (42%) followed by Bacillus (28%) and Enterobacter (21%). Results of oxidase test, catalase test, Gram and spore staining showed that 24 rhizobacterial isolates belonged to Pseudomonas sp. and 31 isolates were found Bacillus (data not shown).
Screening of bacterial isolates for ACC utilization: All the 55 rhizobacterial isolates belonging to Pseudomonas and Bacillus were screened for utilization of ACC on the minimal medium (Dworkin and Foster, 1958) plates supplemented with ammonium sulphate or 3 mM ACC. Twenty two isolates (40.0%) showed growth on ACC supplemented plates (Table 1, Fig. 1). Five rhizobacterial isolates HCS16, HCS35, HCS42, HCS43 and HFS5 showed significant growth on ACC supplemented plates. Three isolates HCS26, HCS33 and HCS34 showed good growth on ammonium sulphate plate and no growth on ACC supplemented plate. Isolates HCS6, HCS13, HCS20, HCS41, HCS44, HFS2, HFS3 and HFS6 showed more growth on ammonium sulphate plate than to ACC supplemented plate. Two isolates HCS36 and HCS40 showed significant growth on ammonium sulphate plate and good growth on ACC supplemented plate whereas, another isolate HCS16 showed significant growth on both ammonium sulphate and ACC supplemented plates. Ten cultures i.e., HCS1, HCS9, HCS11, HCS18, HCS19, HCS28, HCS29, HCS32, HCS37 and HFS8 did not grow on both type of plates and they may require some amino acid for growth.
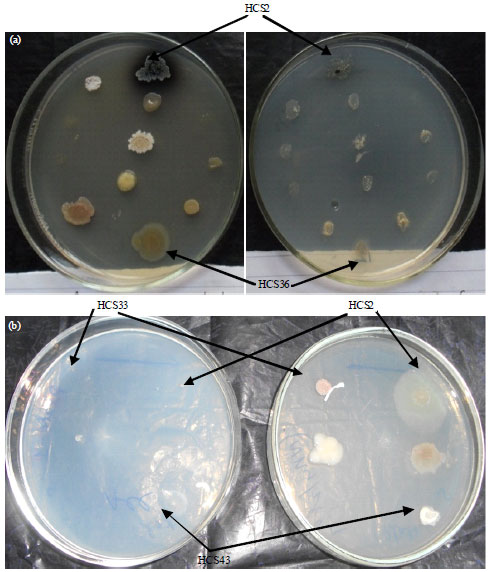 | |
| Fig. 1(a-b): | Growth of rhizobacteria isolates on Dworkin and Foster minimal medium containing either ACC or ammonium sulphate, (a) Medium+Ammonium sulphate plate; Medium+ACC plate and (b) Medium+ACC plate; Medium+Ammonium sulphate plate |
| Table 1: | ACC utilization by rhizobacterial isolates on minimal medium supplemented with ACC or ammonium sulphate |
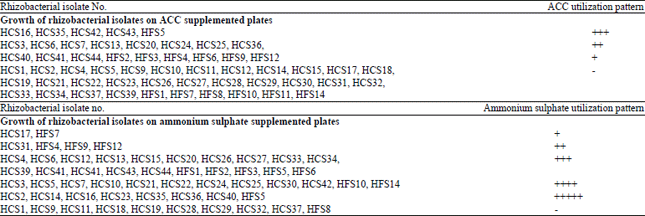 | |
| Growth of bacterial isolates was tested on minimal medium (Dworkin and Foster, 1958) supplemented with ammonium sulphate (2 g L-1) or 3 mM ACC. On the basis of colony size after 2-5 days of incubation at 28±2°C, the growth of the isolates was scored as: -: No growth, +: Little growth, ++: Moderate growth, +++: More growth, ++++: Significant growth, +++++: Maximum growth | |
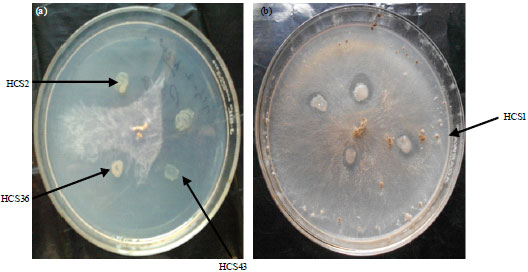 | |
| Fig. 2(a-b): | Rhizobacterial isolates showing antifungal activity against Rhizoctonia solani |
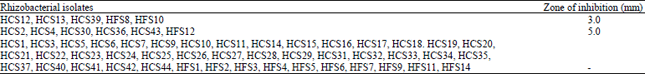
| Table 2: | Antifungal activity of Pseudomonas and Bacillus isolates against Rhizoctonia solani |
 | |
| Antagonistic activity of rhizobacterial isolates was tested on the basis of growth inhibition of fungal pathogens on PDA medium plates by spot test method (Sindhu et al., 1999) | |
Screening of rhizobacterial isolates for antagonistic activity against fungal pathogen under in vitro conditions: Antagonistic activity of all the fifty five rhizobacterial isolates was studied by observing the zone of inhibition of fungal growth on PDA plates. The zone of fungal growth inhibition varied with different isolates tested. Pseudomonas/Bacillus isolates HCS2, HCS4, HCS30, HCS36, HCS43 and HFS12 showed large inhibition zone (5.0 mm) of fungal growth. (Table 2, Fig. 2). Five isolates HCS12, HCS13, HCS39, HFS8 and HFS10 showed 3.0 mm inhibition zone. Isolate HCS2 showed largest inhibition zone. Remaining forty four isolates did not inhibit the growth of fungus R. solani on PDA medium plates. Ten isolates i.e., HCS2, HCS4, HCS12, HCS13, HCS30, HCS36, HCS39, HCS43, HFS10 and HFS12 showed ACC utilization as well as antifungal activity. Cultures HCS4, HCS36 and HCS43 showed more ACC utilization as well as antifungal activity than other isolates.
Coinoculation studies of Rhizobium/Bradyrhizobium and rhizobacterial isolates for disease control and plant growth of clusterbean: Selected rhizobacterial isolates were tested for disease control using wilt susceptible clusterbean variety HG563 under pot house conditions. Clusterbean seeds inoculated with rhizobacterial isolates were grown in pots (with three replications of each treatment) and fungal growth suspension was also inoculated in some treatments. Inoculation of Bradyrhizobium strain GSA11 and Rhizobium isolate GSA110 increased shoot dry weight by 186.33% and 169.63%, respectively as compared to uninoculated control at 30 days of plant growth (Table 3). Coinoculation of Rhizobium strain GSA110 and Bacillus HCS43 formed 42 nodules plant-1 with 209.6 mg nodule weight and shoot dry weight was increased by 207.7% as compared to uninoculated plants. Bradyrhizobium strain GSA11 and Pseudomonas isolate HCS36 along with fungus caused 306.3% increase in shoot dry weight in comparison to control uninoculated plants. Similarly, coinoculation of Bradyrhizobium strain GSA11 and Bacillus isolate HCS43 along with R. solani showed maximum stimulatory effect on nodule formation (47 nodules plant-1) and nodule weight (225.4 mg plant-1) and resulted in 281.5% gains in shoot dry weight. In fungus-inoculated treatments, disease caused by R. solani was effectively controlled by coinoculation of Rhizobium/Bradyrhizobium with rhizobacterial isolates HCS36 and HCS43.
At 60 days of plant growth, nodulation performance of Bradyrhizobium strain GSA11 and Rhizobium strain GSA110 significantly increased and their inoculation resulted in 112.3 and 103.3% gains in shoot dry weight, respectively (Table 4). Coinoculation of Pseudomonas isolate HCS36 and Rhizobium strain GSA110 formed 48 nodules plant-1 and 317.2 mg plant-1 nodule weight was observed. Coinoculation of Rhizobium strain GSA110 with Bacillus isolate HCS43 along with R. solani formed 50 nodules plant-1 and caused 108.9% increase in shoot dry weight in comparison to uninoculated control. Maximum 140.26% increase in shoot dry weight, nodule number (54 nodules plant-1) and nodule weight (346.6 mg plant-1) was observed by coinoculation of Pseudomonas isolate HCS36 with Rhizobium strain GSA110 and fungus R. solani (Fig. 3). Coinoculation of Bacillus isolate HCS43 along with Rhizobium strain GSA110 also reduced the browning of collar root and caused 83.7% disease control (Table 4, Fig. 4) and 108.91% increase in shoot dry weight was observed.
 | |
| Fig. 3(a-c): | Effect of inoculation of Bradyrhizobium GSA11 with rhizobacterial isolate HCS36 for plant growth enhancement under pot house conditions at 60 days of plant growth. (a) Control (uninoculated), (b): Rhizoctonia solani and (c) HCS43+GSA11+R. solani |
| Table 3: | Symbiotic effectiveness and disease control of bacterial isolates on coinoculation of clusterbean at 30 days of plant growth |
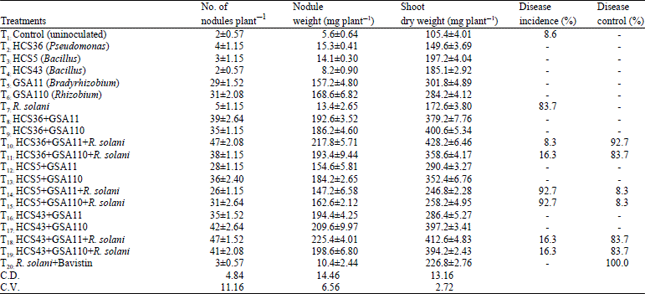 | |
| Values given are average value of three plants. Disease incidence is the % of plants infected and disease control is the % reduction of diseased plants after inoculation with bacteria. The values of nodule fresh and shoot dry weight are calculated as per plant basis. C.D. and C.V. values represent coefficient of deviation and coefficient of variation, respectively. SEM (Standard Error of Means) values are represented as (±) | |
Treatment with bavistin in fungus-inoculated soils caused 92.7% reduction in disease appearance and resulted in 46.7% increase in shoot dry weight as compared to uninoculated control.
DISCUSSION
Previous studies have established that some rhizobacterial strains could serve as a useful biofertilizer and biocontrol agents for various crops (Sindhu et al., 2010). Antagonistic bacteria have also been found to promote the growth of different crops and termed as plant growth-promoting rhizobacteria. Inoculation with rhizobacteria having ACC deaminase activity is an efficient strategy used for lowering the level of stress hormone ethylene to minimize its adverse effect on plant growth. Several bacterial strains that can utilize ACC as a sole source of nitrogen have been isolated from rhizosphere soil samples and subsequently used for inoculation (Glick, 2004). In this study, 40.0% rhizobacterial isolates showed growth on ACC supplemented plates (Table 1, Fig. 1), indicating that these bacteria possess ACC deaminase activity. Five rhizobacterial isolates HCS16, HCS35, HCS42, HCS43 and HFS5 showed significant growth on ACC supplemented plates.
 | |
| Fig. 4(a-c): | Effect of inoculation of Bradyrhizobium and rhizobacterial isolates for disease control under pot house conditions at 60 days of plant growth |
| Table 4: | Symbiotic effectiveness and disease control of bacterial isolates on coinoculation of clusterbean at 60 days of plant growth under pot house conditions |
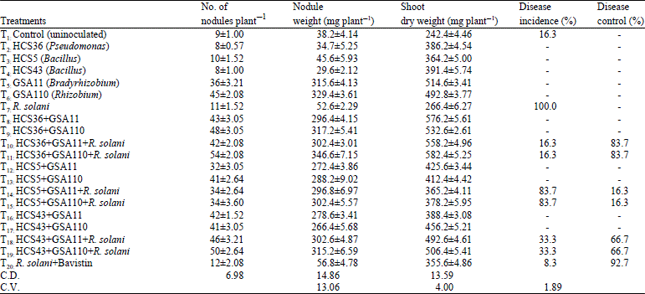 | |
| Values given are average value of three plants. Disease incidence is the % of plants infected and disease control is the % reduction of diseased plants after inoculation with bacteria. The values of nodule fresh and shoot dry weight are calculated as per plant basis. C.D. and C.V. values represent coefficient of deviation and coefficient of variation, respectively. SEM (Standard Error of Means) values are represented as (±) | |
Thanananta et al. (1997) reported that 6 Pseudomonas strains, out of total 55 bacterial isolates, were capable of growing on Dworkin and Foster minimal medium and showed the highest efficiency of ACC utilization. Similarly, the screening of 563 bacteria isolated from the roots of pea, lentil and chickpea showed that only 5% isolates showed ACC deaminase activity and 7% isolates were capable of indole acetic acid production (Hynes et al., 2008). Husen et al. (2009) observed that 11 out of total 13 Pseudomonas isolates possessed ACC deaminase activity and increased root development of soybean. Khandelwal and Sindhu (2012) found that 38.9% Pseudomonas isolates obtained from clusterbean rhizosphere showed good growth on ACC supplemented plates. Chaudhary and Sindhu (2015) observed that 44% Mesorhizobium strains possess the ACC utilization ability. Penrose and Glick (2003) suggested that rhizobial or Pseudomonas strains that are intended for use as inoculants of host legumes should first be selected/ tested for the presence of a functional ACC deaminase to get better crop productivity and disease control.
Screening of rhizobacterial isolates for fungal growth inhibition showed that 20% cultures possess the ability to inhibit pathogenic fungi R. solani under cultural conditions (Table 2). Rhizobacterial isolates HCS2, HCS4, HCS30, HCS36, HCS43 and HFS12 showed 5.0 mm fungal growth inhibition zone (Fig. 2). Five isolates HCS12, HCS13, HCS39, HFS8 and HFS10 showed 3.0 mm inhibition zone. Similar growth inhibition of R. solani has been reported by P. fluorescens (PS-4) strain and the antagonistic potential was attributed to the production of antibiotics and siderophores or due to induced systemic resistance (Kaiser and Hannan, 1989). Siddiqui et al. (2001) showed that Pseudomonas aeruginosa and Bacillus subtilis strains produced growth inhibition zones by inhibiting the radial growth of Macrophomina phaseolina, Fusarium oxysporum and Rhizoctonia solani. Coombs et al. (2004) evaluated 38 stains belonging to Streptomyces and Microspora for their ability to produce antifungal compounds in vitro against G. graminis var. tritici, R. solani and Pythium sp. They observed that 64% cultures exhibited antifungal activity under cultural conditions. Similarly, Pseudomonas aeruginosa strain isolated from Botanical Garden at Udaipur, Rajasthan was used as biocontrol agent against three phytopathogenic fungi Fusarium moniliformae, Alternaria solani and Helminthosporium halodes (Sharma et al., 2007). Karuppiah and Rajaram (2011) reported that eight Bacillus sp., out of 63 different Bacillus isolates, exhibited plant growth promoting activities and six of these Bacillus isolates also inhibited the growth of Penicillium sp., Cercospora sp. and Fusarium oxysporum. Dua and Sindhu (2012) isolated antagonistic bacteria from the rhizosphere soil of wheat and sixteen bacterial isolates were found to inhibit the growth of R. solani. Growth inhibition zone varied from 6-15 mm by different rhizobacterial isolates. Growth inhibition of the pathogenic fungi was also observed by using culture filterates of antagonistic rhizobacterial isolates.
Inoculation of Bradyrhizobium isolate GSA11 and Rhizobium isolate GSA110 on clusterbean seeds increased shoot dry weight by 186.33 and 169.63%, respectively as compared to uninoculated control at 30 days of plant growth (Table 3). Coinoculation of Bradyrhizobium isolate GSA11 with Pseudomonas isolate HCS36/Bacillus isolate HCS43 and R. solani formed maximum 47 nodules plant-1 and increased shoot dry weight by 306.3 and 281.5%, respectively as compared to uninoculated control at 30 days of plant growth under pot house conditions. Disease caused by R. solani was effectively controlled on coinoculation of Rhizobium/Bradyrhizobium with rhizobacterial isolates HCS36 and HCS43 in fungus inoculated soil. At 60 days of plant growth, coinoculation of Pseudomonas isolate HCS36 and Rhizobium strain GSA110 formed 48 nodules per plant and 317.2 mg plant-1 nodule weight was observed (Table 4, Fig. 3). Coinoculation of Bacillus isolate HCS43 along with Rhizobium strain GSA110 reduced the browning of collar root and caused 83.7% disease control (Fig. 4).
In earlier studies, a significant correlation was found between in vitro ACC deaminase activity of the rhizobacteria and growth promoting activity of these bacteria on maize under axenic conditions and on nodulation of mung bean (Vigna radiata L.) under natural pot and field trials (Shaharoona et al., 2006). Ma et al. (2003) found that ACC deaminase lacking mutant of Rhizobium leguminosarum bv. viciae strain 128C53K formed approximately 25% fewer nodules than the wild-type strain on Pisum sativum L. cv. Sparkle and also produced approximately 30% less plant biomass. Afsharmanesh et al. (2010) evaluated the antagonistic potential of Pseudomonas fluorescens strain UTPF5 against R. solani AG-4 in bean and showed that strain UTPF5 could inhibit the growth of R. solani both in vitro and in vivo and suppressed the disease by 33.34 and 14.29% in soil drenching and seed treatment, respectively. Susilowati et al. (2011) reported that Pseudomonas sp. CRB inhibited the growth of the pathogenic fungi i.e., Sclerotium rolfsii, Fusarium oxysporum and Rhizoctonia solani, approximately 11.1-60.0% in vitro. Seed coating with the Pseudomonas sp. CRB accomplished disease suppression in plant about 14.3-100% in sterile soil condition and 5.2-52.6% in non sterile soil condition.
Abeysinghe (2009) reported that a combination of two compatible biological control agents, Bacillus subtilis CA32 and Trichoderma harzianum RU01 showed significant plant protection of Solanum melongena and Capsicum annuum for control of damping-off disease caused by Rhizoctonia solani. Dua and Sindhu (2012) showed that inoculation of R. solani in wheat caused 85-90% root rot disease at 60-90 days of plant growth. Whereas, coinoculation of antagonistic Pseudomonas isolates WPS3 or WPS90 in R. solani-inoculated plants caused 88.9 and 66.7% disease control, respectively at 90 days of plant growth in the pots. Coinoculation of Pseudomonas isolate WPS3 or WPS90 with R. solani enhanced 115 and 98% plant dry weight in comparison to uninoculated plants at 90 days of plant growth. Similar, synergistic effects have been observed on nodulation and plant growth of other legumes by dual inoculation of B. japonicum and P. fluorescens in soybean (Li and Alexander, 1988), R. leguminosarum with an antibiotic-producing P. fluorescens strain F113 in pea (Andrade et al., 1998) and Bradyrhizobium/Mesorhizobium strains with Pseudomonas sp. in green gram and chickpea (Goel et al., 2000; Sindhu et al., 2002). Combined application of Pseudomonas and Bacillus strains has also been found to control effectively the tomato wilt and damping-off disease in alfalfa (Sundaramoorthy and Balabaskar, 2013; Sarhan and Shehata, 2014). Thus, it appears that use of mixtures or combinations of biocontrol agents may be more effective method to improve biological control (Spadaro and Gullino, 2005) leading to enhanced crop productivity. For realistic inoculation effects of this study, Bacillus isolate HCS43 and Pseudomonas isolate HCS36 needs to be evaluated under the field conditions for disease control and plant growth stimulation of clusterbean.
CONCLUSION
Pseudomonas and Bacillus isolates obtained from the rhizosphere soil of clusterbean were found to inhibit the growth of root rot-causing fungi Rhizoctonia solani on PDA medium plates. Seed inoculation of antagonistic Bacillus isolate HCS43 and Pseudomonas isolate HCS36 caused 66.7-83.7% reduction of root rot disease in clusterbean under pot house conditions. Coinoculation of Pseudomonas/Bacillus isolates with Bradyrhizobium/Rhizobium strain GSA11 or GSA110 in clusterbean enhanced the nodule number, nodule fresh weight and plant dry weight as compared to Bradyrhizobium/Rhizobium-inoculated or uninoculated control plants and also suppressed the root rot disease under pot house conditions. These results suggested that rhizosphere bacteria from the soil could minimize/replace the use of fungicides and nitrogenous fertilizers to reduce the pollution of soil and environment.
REFERENCES
- Abeysinghe, S., 2009. Effect of combined use of Bacillus subtilis CA32 and Trichoderma harzianum RU01 on biological control of Rhizoctonia solani on Solanum melongena and Capsicum annuum. Plant Pathol. J., 8: 9-16.
CrossRefDirect Link - Afsharmanesh, H., M. Ahmadzadeh, M. Javan-Nikkhah and K. Behboudi, 2010. Characterization of the antagonistic activity of a new indigenous strain of Pseudomonas fluorescens isolated from onion rhizosphere. J. Plant Pathol., 92: 187-194.
Direct Link - Andrade, G., F.A.A.M. De Leij and J.M. Lynch, 1998. Plant mediated interactions between Pseudomonas fluorescens, Rhizobium leguminosarum and arbuscular mycorrhizae on pea. Lett. Applied Microbiol., 26: 311-316.
CrossRefDirect Link - Bajoria, S., A.K. Varshney, R.P. Pareek, M.K. Mohan and P. Ghosh, 2008. Screening and characterization of antifungal clusterbean (Cyamopsis tetragonoloba) rhizobacteria. Biocontrol Sci. Technol., 18: 139-156.
CrossRefDirect Link - Chaudhary, D. and S.S. Sindhu, 2015. Inducing salinity tolerance in chickpea (Cicer arietinum L.) by inoculation of 1-aminocyclopropane-1-carboxylic acid deaminase containing Mesorhizobium strains. Afr. J. Microbiol. Res., 9: 117-124.
Direct Link - Coombs, J.T., P.P. Michelsen and C.M.M. Franco, 2004. Evaluation of endophytic actinobacteria as antagonists of Gaeumannomyces graminis var. tritici in wheat. Biol. Control, 29: 359-366.
CrossRef - Dua, S. and S.S. Sindhu, 2012. Effectiveness of rhizosphere bacteria for control of root rot disease and improving plant growth of wheat (Triticum aestivum L). J. Microbiol. Res., 2: 26-35.
CrossRefDirect Link - Dworkin, M. and J.W. Foster, 1958. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol., 75: 592-601.
Direct Link - Glick, B.R., 2004. Bacterial ACC deaminase and the alleviation of plant stress. Adv. Applied Microbiol., 56: 291-312.
CrossRefDirect Link - Glick, B.R., 2014. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res., 169: 30-39.
CrossRefDirect Link - Goel, A.K., S.S. Sindhu and K.R. Dadarwal, 2000. Pigment diverse mutants of Pseudomonas sp.: Inhibition of fungal growth and stimulation of growth of Cicer arietinum. Biol. Plant., 43: 563-569.
CrossRefDirect Link - Husen, E., A.T. Wahyudi, A. Suwanto and R. Saraswati, 2009. Soybean seedling root growth promotion by 1-aminocyclopropane-1-carboxylate deaminase-producing pseudomonads. Indonesian J. Agric. Sci., 10: 19-25.
Direct Link - Hynes, R.K., G.C. Leung, D.L. Hirkala and L.M. Nelson, 2008. Isolation, selection and characterization of beneficial rhizobacteria from pea, lentil and chickpea grown in Western Canada. Can. J. Microbiol., 54: 248-258.
CrossRefPubMedDirect Link - Kaiser, W.J. and R.M. Hannan, 1989. Biological control of seed rot and preemergence damping-off of chickpea with fluorescent pseudomonads. Soil Biol. Biochem., 21: 269-273.
CrossRefDirect Link - Karuppiah, P. and S. Rajaram, 2011. Exploring the potential of chromium reducing Bacillus sp. and there plant growth promoting activities. J. Microbiol. Res., 1: 17-23.
CrossRefDirect Link - Keel, C., U. Schnider, M. Maurhofer, C. Voisard and K. Laville et al., 1992. Suppression of root diseases by Pseudomonas fluorescens CHA0: Importance of the bacterial secondary metabolite 2,4-diacetylphloroglucinol. Mol. Plant Microb. Interact., 5: 4-13.
Direct Link - Khandelwal, A. and S.S. Sindhu, 2012. Expression of 1-aminocyclopropane-1-carboxylate deaminase in rhizobia promotes nodulation and plant growth of clusterbean (Cyamopsis tetragonoloba L.). Res. J. Microbiol., 7: 158-170.
Direct Link - Li, D.M. and M. Alexander, 1988. Co-inoculation with antibiotic-producing bacteria to increase colonization and nodulation by rhizobia. Plant Soil, 108: 211-219.
CrossRef - Ma, W., F.C. Guinel and B.R. Glick, 2003. Rhizobium leguminosarum Biovar viciae 1-aminocyclopropane-1-carboxylate deaminase promotes nodulation of pea plants. Applied Environ. Microbiol., 69: 4396-4402.
CrossRef - Penrose, D.M. and B.R. Glick, 2003. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiologia Plantarum, 118: 10-15.
CrossRefDirect Link - Raaijmakers, J.M., M. Leeman, M.M.P. van Oorschot, I.V. Sluis, B. Schippers and P.A.H.M. Bakker, 1995. Dose-response relationships in biological control of Fusarium wilt of radish by Pseudomonas spp. Phytopathology, 85: 1075-1081.
CrossRef - Ramette, A., Y. Moenne-Loccoz and G. Defago, 2006. Genetic diversity and biocontrol potential of fluorescent pseudomonads producing phloroglucinols and hydrogen cyanide from Swiss soils naturally suppressive or conducive to Thielaviopsis basicola-mediated black root rot of tobacco. FEMS Microbiol. Ecol., 55: 369-381.
CrossRefDirect Link - Saikia, R., K. Singh and D. Arora, 2004. Suppression of Fusarium-wilt and charcoal rot of chickpea by Pseudomonas aeruginosa RsB29. Indian J. Microbiol., 44: 181-184.
Direct Link - Sahu, G.K. and S.S. Sindhu, 2011. Disease control and plant growth promotion of green gram by siderophore producing Pseudomonas sp. Res. J. Microbiol., 6: 735-749.
CrossRefDirect Link - Saraf, M., U. Pandya and A. Thakkar, 2014. Role of allelochemicals in plant growth promoting rhizobacteria for biocontrol of phytopathogens. Microbiol. Res., 169: 18-29.
CrossRefDirect Link - Sarhan, E.A.D. and H.S. Shehata, 2014. Potential plant growth-promoting activity of Pseudomonas spp. and Bacillus spp. as biocontrol agents against damping-off in alfalfa. Plant Pathol. J., 13: 8-17.
CrossRefDirect Link - Shaharoona, B., M. Arshad and Z.A. Zahir, 2006. Effect of plant growth promoting rhizobacteria containing ACC-deaminase on maize (Zea mays L.) growth under axenic conditions and on nodulation in mung bean (Vigna radiata L.). Lett. Applied Microbiol., 42: 155-159.
CrossRefDirect Link - Siddiqui, I.A., S. Ehetshamul-Haque and S.S. Shaukat, 2001. Use of rhizobacteria in the control of root rot-root knot disease complex of mungbean. J. Phytopathol., 149: 337-346.
CrossRefDirect Link - Sindhu, S.S. and K.R. Dadarwal, 2001. Chitinolytic and cellulolytic Pseudomonas sp. antagonistic to fungal pathogens enhances nodulation by Mesorhizobium sp. Cicer in chickpea. Microbiol. Res., 156: 353-358.
CrossRefDirect Link - Sindhu, S.S., S.K. Gupta and K.R. Dadarwal, 1999. Antagonistic effect of Pseudomonas spp. on pathogenic fungi and enhancement of growth of green gram (Vigna radiata). Biol. Fertil. Soils, 29: 62-68.
CrossRefDirect Link - Sindhu, S.S., S. Suneja, A.K. Goel, N. Parmar and K.R. Dadarwal, 2002. Plant growth promoting effects of Pseudomonas sp. on coinoculation with Mesorhizobium sp. Cicer strain under sterile and wilt sick soil conditions. Applied Soil Ecol., 19: 57-64.
CrossRefDirect Link - Sloger, C., 1969. Symbiotic effectiveness and N2 fixation in nodulated soybean. Plant Physiol., 44: 1666-1668.
Direct Link - Spadaro, D. and M.L. Gullino, 2005. Improving the efficacy of biocontrol agents against soilborne pathogens. Crop Prot., 24: 601-613.
CrossRef - Sundaramoorthy, S. and P. Balabaskar, 2013. Evaluation of combined efficacy of Pseudomonas fluorescens and Bacillus subtilis in managing tomato wilt caused by Fusarium oxysporum f. sp. lycopersici (Fol). Plant Pathol. J., 12: 154-161.
Direct Link - Susilowati, A., A.T. Wahyudi, Y. Lestari, A. Suwanto and S. Wiyono, 2011. Potential Pseudomonas isolated from soybean rhizosphere as biocontrol against soilborne phytopathogenic fungi. Hayati J. Biosci., 18: 51-56.
Direct Link - Wang, C., E. Knill, B.R. Glick and G. Defago, 2000. Effect of transferring 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase genes into Pseudomonas fluorescens strain CHA0 and its gacA derivative CHA96 on their growth-promoting and disease-suppressive capacities. Can. J. Microbiol., 46: 898-907.
PubMedDirect Link - Weller, D.M., 2007. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology, 97: 250-256.
CrossRef - Yang, Q.Y., K. Jia, W.Y. Geng, R.J. Guo and S.D. Li, 2014. Management of cucumber wilt disease by Bacillus subtilis B006 through suppression of Fusarium oxysporum f. sp. cucumerinum in rhizosphere. Plant Pathol. J., 13: 160-166.
CrossRefDirect Link