
Roni Pazla
Faculty of Animal Science, Andalas University, Padang, Indonesia
LiveDNA: 62.20146
Novirman Jamarun
Faculty of Animal Science, Andalas University, Padang, Indonesia
Mardiati Zain
Faculty of Animal Science, Andalas University, Padang, Indonesia
Arief
Faculty of Animal Science, Andalas University, Padang, Indonesia
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 10 | Page No.: 462-470







ABSTRACT
Objective: The aim of this research was to measure microbial protein synthesis (MPS) in Phanerochaete chrysosporium and determine its in vitro nutrient digestibility and fermentability of fermented oil palm fronds (FOPF) under supplementation with Ca, P and Mn minerals and grown with tithonia (T) and elephant grass (EG). Methodology: This study used a randomized block design consisting of four treatment groups: T1 = 20% FOPF+16% T+64% EG, T2 = 40% FOPF+12% T+48% EG, T3 = 60% FOPF+8%, T+32% EG and T4 = 80% FOPF+4% T+16% EG. Four replicates were used per treatment. The data were analyzed using one-way analysis of variance (ANOVA) and differences among the means were tested using Duncan’s multiple range tests (DMRT) with 5 and 1% confidence intervals. The variables measured were crude protein (CP) digestibility (CPD), cellulose digestibility (CD), ammonia (NH3) concentration, total volatile fatty acid (VFA) content and fluid ruminal pH values as indicators of fermentability and MPS. Results: The results showed that CPD, CD, NH3 concentration, VFA content and MPS were the highest (p<0.01) in the T1 treatment. However, pH was lowest (p<0.01) in T1. Conclusion: The combination of 20% FOPF+16% T+64% EG had the best effect on MPS, in vitro nutrient digestibility and fermentability.
PDF Abstract XML References Citation
Received: January 31, 2018;
Accepted: July 06, 2018;
Published: September 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Roni Pazla, Novirman Jamarun, Mardiati Zain and Arief, 2018. Microbial Protein Synthesis and in vitro Fermentability of Fermented Oil Palm Fronds by Phanerochaete chrysosporium in Combination with Tithonia (Tithonia diversifolia) and Elephant Grass (Pennisetum purpureum). Pakistan Journal of Nutrition, 17: 462-470.
DOI: 10.3923/pjn.2018.462.470
URL: https://scialert.net/abstract/?doi=pjn.2018.462.470
DOI: 10.3923/pjn.2018.462.470
URL: https://scialert.net/abstract/?doi=pjn.2018.462.470
INTRODUCTION
One of the priorities of the Indonesian government is the achievement of self-sufficiency in milk by 20201. To achieve this goal, livestock seed availability and feed adequacy are necessary. Meet the necessary forage feed provisions to increase milk production, adequate and appropriate land availability for forage growth is urgent. Population growth has changed the land available for forage cultivation into residential areas. This change has prompted researchers to find alternative feed to meet the needs of cattle forage.
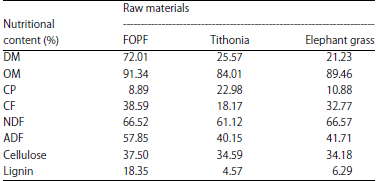
Palm fronds as a waste from oil palm plantations have potential for use as alternative feed forage replacement. Palm frond production from palm oil plantations reaches 69 t ha–1 year–1 or 20 t of dry matter (DM)/ha/year2. This waste is highly abundant throughout the year, but its use as livestock feed is not maximized, especially on farms. Jamarun et al.3 reported that the nutritional content of palm fronds was as follows: DM, (30.43%), organic matter (OM), (94.13%), crude protein (CP) (3.64%), crude fiber (CF) (49.80%), neutral detergent fiber (NDF) (89.98%), acid detergent fiber (ADF) (73.21%), cellulose (41.35%) and lignin (30.63%). The utilization of palm fronds as feed is limited because of the high lignin content4.
Lignin is a natural phenolic polymer that is commonly found in plant cell walls and is a limiting factor in animal feed due to its low digestibility5. One effort that can be made to reduce the lignin content in palm fronds is fermentation using the mold Phanerochaete chrysosporium. The LiP and MnP enzymes produced by P. chrysosporium during bioconversion are among those that first degrade lignin components and then cellulose components6, resulting in a rapid lag phase of approximately 0-3 days7 and the ability to grow optimally at 40°C8, making this species suitable for fermentation generating a lot of heat. Fermentation of palm fronds for 20 days using P. chrysosporium supplemented with Ca, P and Mn minerals can decrease the lignin content by 40.08%3. The digestibility and microbial protein synthesis (MPS) increase as the lignin content in palm fronds decreases9-11. A low lignin content makes it easier for rumen microbes to degrade feed so that more nutrients can be utilized as a source of energy for livestock and rumen microbes.
Tithonia (T) plants also have potential as an alternative feed. T cultivated in West Sumatra can produce as much as 30 t of fresh material or 6 t of DM per year on a land area of approximately 1/5 ha. When grown as a hedge, it can produce 27 kg of dry weight from three harvests in 1 year12. Jamarun et al.13 reported the following nutritional contents of T: DM (25.57%), OM (84.01%), CP (22.98%), CF (18.17%), NDF (61.12%), ADF (40.15%), cellulose (34.59%) and lignin (4.57%). Jama et al.14 reported that the green leaves of T contained high levels of N (3.5-4.0%), P (0.35-0.38%), K (3.5-4.1%), Ca (0.59%) and Mg (0.27%). Research on T as a ruminant feed material is limited because its leaves contain many antinutritional substances that endanger livestock, such as phytic acid, tannins, saponins, oxalates, alkaloids and flavonoids15. The combination of T and elephant grass (EG) can improve the digestibility and fermentability of the rumen fluid. Jamarun et al.16 reported a combination of 20% T and 80% EG in vitro resulting in the DM and OM digestibilities of 58.30 and 57.85%, respectively, pH 6.68, ammonia (NH3) concentration of 16.89 mg/100 mL and volatile fatty acid (VFA) content of 127.50 mM. These values continued to decrease with the addition of T to the ration. The provision of diets with varying forage will increase VFA production and MPS in Etawa goat crossbreeds17. The forage composition of the feed greatly affects the response of livestock growth and production.
The aim of this study was to determine the MPS, in vitro digestibility and fermentability of oil palm fronds (OPF) fermented by P. chrysosporium, supplemented with the minerals (Ca, P and Mn) and combined with T and EG.
MATERIALS AND METHODS
For this study, the OPF were fermented at the Laboratory of Technology for Feed Industries, Andalas University. Analysis of in vitro digestibility and rumen microbial populations was conducted in the Laboratory of Dairy Nutrition of the Bogor Agricultural Institute. Samples of fermented OPF (FOPF), T and EG were collected around the town of Padang. All samples were finely ground and mixed according to the treatment combinations. The parameters measured in this study were CP digestibility (CPD), cellulose digestibility (CD), NH3 concentration, total VFA content, pH and MPS.
Fermented oil palm fronds, tithonia and elephant grass: The OPF materials used as the raw material in this study were taken from the distal two-thirds of the OPF. The OPF substrates were cut, dried and finely milled. The OPF were then fermented using P. chrysosporium, supplemented with 2000 ppm of Ca, 2000 ppm of P and 150 ppm of Mn, according to the procedure described by Jamarun et al.9.
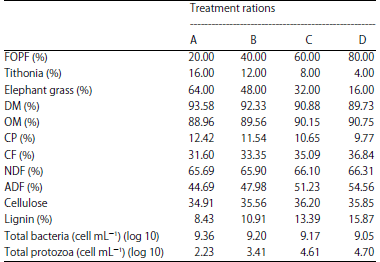
The T plants used in this study had soft plant tissues (stems, leaves and flowers) with a 6-16 cm long trunk measured from the top. T and EG were cut, dried and crushed. The ratio of T and EG in each treatment ratio was 1:4, based on Jamarun et al.13 The nutritional contents of FOPF, T and EG are presented in Table 1. The composition of the feed ingredients and the nutrition of the treatment rations are presented in Table 2.
| Table 1: | Nutritional content of FOPF, tithonia and elephant grass |
 | |
| Table 2: | Composition of feed ingredients and nutrition of treatment rations |
 | |
In vitro procedure: The in vitro procedure in this study followed that of Tilley and Terry18. Rumen fluid was taken from a cow fistula at Bogor, LIPI. The fermenter tube was filled with 0.5 g of sample and 40 mL of McDougall solution was added. The tube was placed in a shaker bath at 39°C, filled with 10 mL of rumen fluid and shaken with CO2 for 30 sec, the pH was checked (6.5-6.9) and the sample was covered with a ventilated rubber cap and fermented for 48 h. After 48 h, the rubber cap of the fermenter tube was removed and 2-3 drops of HgCl2 were added to kill the microbes. The fermenter tube was centrifuged at 5,000 rpm for 15 min. The substrate was separated into a precipitate layer at the bottom and a clear supernatant at the top. The supernatant was removed, the resulting sediment was centrifuged at 5,000 rpm for 15 min and 50 mL of 0.2% pepsin-HCl solution was added. This mixture was then reincubated for 48 h without a rubber cap. The remaining digested residue was filtered using Whatman filter paper No. 41 (identified by weight) with the help of a vacuum pump. The precipitate on the filter paper was placed in a porcelain dish that was placed in a 105°C oven for 24 h. After 24 h, the porcelain cup+filter paper+residue was removed, inserted into a desiccator and weighed to determine the DM content. Furthermore, the ingredients in the cup were placed in a kiln or in an electric furnace for 6 h at 450-600°C and weighed to determine the amount of organic material. Residues originating from fermentation without feed ingredients were used as a control. The proximate analysis of materials and residues followed the AOAC19 and Van Soest et al.20 procedure.
Rumen fluid characteristics: The measured rumen fluid characteristics were as follows: Ruminal pH, NH3-N concentration and VFA content. Fermenter tubes were each filled with 0.5 g of sample and 40 mL of buffer solution and 10 mL of fresh rumen fluid (ratio of 4:1) were added. After the tube received CO2, the tubes were closed with a rubber ventilator. Tubes were inserted into the fermenter shaker water bath at a temperature of 39°C and incubated for 4 h, followed by analyses of pH, NH3 and VFA. The ruminal pH was measured using a pH meter. The NH3-N concentration was measured using the micro diffusion (Conway) method and the total VFA content was measured using the steam distillation method.
Microbial protein synthesis: MPS was measured using a centrifuge and a spectrophotometer. The centrifuge used in this study was a Beckman J2-21 centrifuge, while the spectrophotometer used was an LW scientific spectrophotometer UV-200Rs. To measure MPS, a 20-mL sample of rumen fluid was initially centrifuged at 400 rpm for 45 sec to separate the bacteria. The samples were then centrifuged at 408 rpm for 5 min to reduce the population of protozoa in the rumen fluid and to remove any remaining food particles. Aliquots (rumen fluid that had been centrifuged at 408 rpm with a decreased number of protozoa and no food particles) were taken and 64.5% trichloroacetic acid (TCA) was added to each 10 mL sample (up to 2.5 mL per sample). The samples were centrifuged at 15,000 rpm for 20 min, the supernatant was discarded and the obtained cells/sediment were retained and washed with distilled water. The cell/sediment precipitate was centrifuged again at 15,000 rpm for 20 min. The supernatant was discarded and the cell/sediment was added to a 30-mL solution of NaOH (0.25 N). The precipitate was heated in boiling water for 10 min. The resulting supernatant was collected from each sample and 1 mL was used for analysis of microbial proteins according to Plummer21.
Experimental design and statistical analysis: The study was conducted using a randomized block design (4×4) with 4 replications (levels of FOPF, T and EG as the treatments). The differences between the treatment means were analyzed using Duncan’s multiple range tests (DMRT) with confidence intervals of 5 and 1%.
The treatments were as follows:
| T1 | = | 20% FOPF+16% T+64% EG |
| T2 | = | 40% FOPF+12% T+48% EG |
| T3 | = | 60% FOPF+8% T+32% EG |
| T4 | = | 80% FOPF+4% T+16% EG |
RESULTS AND DISCUSSION
Crude protein digestibility: A higher digestibility rate of a ration suggests that the ration is of good quality for cattle consumption and can meet the cattle’s metabolic needs. CPD in this study ranged from 50.18-70.71% (Table 3). This value is higher than the results from the study of Rahayu et al.22, who measured a CPD of 40.25-46.45% in FOPF fermented with P. chrysosporium without combination with other forages. The combination of EG and T is only capable of producing a CPD of 42.40-66.14%13. This suggests that combined treatment with different types of forage can improve the CPD.
Table 3 shows that the highest CPD (p<0.01) was observed in T1, followed by T2 and T3. T4 had the lowest CPD. The high CPD in T1 was due to the CP content in the T1 ration being higher than in the other treatments (Table 2). High proteins contents provide more N for microbial growth. Nearly 80% of rumen microbes require N for protein synthesis. Good microbial growth also improves feed digestibility. Elihasridas23 stated that the growth and development of rumen microorganisms are highly dependent on the availability of nutrients and precursors, such as amino acids, N and minerals. The dose of T in T1 (16%) is the optimal and maximum dose in terms of the availability of amino acids, proteins and minerals for rumen microbe growth. Fasuyi et al.15 reported that T leaves contain mostly complex amino acids, such as lysine, arginine, aspartate, glutamate, methionine, cystine, isoleucine, tyrosine and phenylethyl. T contains many essential amino acids for the growth of rumen microbes, such as methionine, leucine, isoleucine and valine24, as well as sufficient levels of P and Mg14, which are essential for rumen microbe growth11.
Phytic acid compounds are antinutrients with high concentrations in T plants, reaching 79.2 mg/100 g24. In contrast to monogastric animals, Ruminantia species have rumen microbes that can produce phytase, including the bacteria Actinobacillus sp. and Bacillus pumilus25. This enzyme is capable of breaking the P bond of phytate so that P can be absorbed and used as a mineral source for ruminants.
| Table 3: | Means of crude protein and cellulose digestibility of FOPF by Phanerochaete chrysosporium in combination with tithonia and elephant grass |
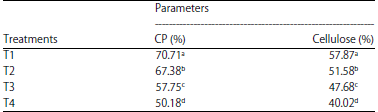 | |
| Means in the same column with different letters (a, b, c and c) are significant (p<0.01) | |
Phytase in the rumen can also form complex bonds with Zn as Zn-phytate complexes, which are likely to be degraded and release Zn slowly to be used for the growth of rumen microbes and livestock26.
The CPD in T is high, at 16%, due to optimal phytic acid antinutrients being degraded by rumen microbial phytase enzymes, resulting in P mineral availability for the growth and development of optimum rumen microbes. P is an essential component of microbial growth, maintaining the integrity of cell membranes, cell walls, components of nucleic acids and parts of high-energy molecules (ATP, ADP, etc.)27-28. Some feed ingredients are deficient in P, so P is added to the feed to promote rumen microbial growth and thereby increase digestibility29-32. Supplementation of P, S and Mg can increase nutrient digestibiity29.
The digestibility of ruminant livestock feed depends on the population and type of rumen microbes, especially bacteria, because the feed is broken down by enzymes produced by rumen microbes. Therefore, increasing the population of rumen bacteria will increase the concentration of these enzymes and thereby improve feed digestibility. Table 2 shows that T1 yielded the highest bacterial population, at 9.36 (log 10) cells/mL. Protein digestibility is related to the rumen fluid populations, especially the proteolytic population. The probable proportion of proteolytic bacteria in T1 was higher than that in the other treatments, so the ability of the bacteria to degrade protein was higher in this treatment.
The T1 had a low crude fiber content (31.60%) (Table 2) compared to that of the other treatments. A high crude fiber content would decrease the digestibility of other nutrients and a high coarse fiber content inhibits rumen microbial degradation of feed3.
Cellulose digestibility: Ruminants require adequate dietary fiber for normal rumen activity and function. Dietary fibers are degraded by microbes acting as energy providers to support basic life, growth, lactation and reproduction33. The role of dietary fiber as a source of energy is closely related to the proportion of constituent fiber components, such as cellulose, hemicellulose and lignin. Cellulose is a complex carbohydrate. Cellulose digestion in the rumen produces energy in the form of VFAs. CD is strongly influenced by the dietary lignin content. Lignin is a natural phenol compound that is usually found in the cell wall of plants and as the limiting factor of digestibility in livestock5. Lignin is a barrier to cellulosic hydrolysis, since lignin protects against attack from cellulose-degrading enzymes34.
Table 3 shows that the CD in this study ranged from 40.02-57.87%. The highest CD was observed in T1, followed by T2 and T3. The lowest CD was observed in T4. The low CD in T4 caused T4 to have the highest lignin content (Table 2) among treatments. Lignin binds cellulose in the form of lignocellulosic bonds, making it difficult for rumen bacteria to degrade feed. This condition will reduce the population of rumen bacteria due to a lack of nutrients. Table 2 shows the lower bacterial population in T4 (9.05 log10 cells/mL) than in T3, T2 and T1. Cellulosic degradation is strongly influenced by the number and activity of cellulolytic bacteria. The number of suspected cellulolytic bacterial populations in T4 was the lowest. Low number and activity of cellulolytic bacteria will decrease CD and vice versa35.
T4 contained 80% FOPF. This high FOPF level contributed a high lignin content to the ration. Although the OPF were fermented using P. chrysosporium, the resulting lignin content was not comparable to the low lignin contents of EG and T (Table 1). Thus, the use of high FOPF levels in automatic rations will decrease CD because of the strong lignocellulosic bonds that must be degraded by rumen bacteria.
The CD in this study was higher than the CD in FOPF fermented by P. chrysosporium without combination with other forage ingredients in studies by Mariani36 and Rahayu et al.22, amounting to 28, 263-32, 558 and 37.37-43.13%, respectively. However, the CD in this study was almost the same as that reported by Jamarun et al.16, who found that the combination of T and EG produced a CD of 41.25-59.65%.
NH3-N: The NH3 concentration reflects the amount of ration protein available in the rumen and its value is strongly influenced by the ability of rumen microbes to degrade the ration protein. NH3 is an important source of N for microorganisms that live in the rumen, where it is used for the synthesis of microbial proteins. According to McDonald et al.37, the optimal range of NH3 concentrations for rumen MPS is from 6-21 mMol. Paengkoum et al.38 stated that the N-NH3 concentration required for rumen microbes to digest feed maximally was 5-20 mg dL–1, equivalent to 3.57-14.28 mM.
| Table 4: | Mean NH3 concentration, VFA content and pH of FOPF by Phanerochaete chrysosporium in combination with tithonia and elephant grass |
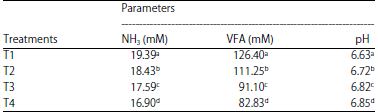 | |
Means in the same column with different letters (a, b, c and d) are significant (p<0.01) | |
Table 4 shows that the treatment had a significant effect (p>0.01) on the NH3 concentration in rumen fluid. The average NH3 concentration in this study was 16.90-19.39 mM. It can be concluded that the availability of N-NH3 in the rumen fluid in these 4 treatments was normal, as this value includes the optimum value for rumen microbial growth.
T1 yielded the highest NH3 concentration (19.39 mM) (p<0.01). This indicates that the protein quality in T1 was higher than that in the other treatments. Thus, the high NH3 concentration in this treatment demonstrated the ability of microbes to degrade feed proteins and was also higher than that in the other treatments. The increased NH3 availability will provide N balance and sufficient energy for rumen microbial growth. The growth rate of rumen microbes can determined from the number of bacteria (Table 2) and CPD (Table 3), which was very high in T1. Mukhtarudin and Liman39 stated that the NH3 concentration could indicate the fermentability of feed associated with CPD and the number and activity of rumen microbes.
The protein content in the ration can affect the NH3 concentration in the rumen. The ration protein content in T1 was the highest value (Table 2). Some of the proteins that enter the rumen will be converted to NH3 by proteolytic enzymes produced by rumen microbes. The production of NH3 depends on the solubility of dietary protein, the protein content in the ration, the duration that food is in the rumen and the pH of the rumen40.
Total VFA content: VFAs in ruminant cattle have a dual function: as a source of carbon for the formation of microbial proteins and as a source of energy for ruminant livestock. Table 4 shows that the average total VFA content was 82.83-126.40 mM. T1 had the highest VFA content (p<0.01) and T4 had the lowest. The high total VFA content in T1 was due to the great conversion of crude fiber and other cell wall components (cellulose, hemicellulose) into simple molecules (VFAs). The higher the fermentability of a feed material is, the greater the content of VFAs derived from protein. The high protein content in T1 (Table 2) also contributed to the high VFA content. VFAs come not only from carbohydrates such as cellulose but also from proteins. Although cellulose levels were the highest in T4, the VFA content was the lowest. This is because the lignin content in T4 (Table 2) was much higher than that in the other treatments. Lignin strongly binds cellulose and hemicellulose. This bond is difficult to penetrate by enzymes produced by rumen bacteria. Therefore, cellulose cannot easily be fermented into VFAs.
The high total VFA content in T1 was also caused by increased fermentation due to the increased number of rumen microbes (Table 2). The results obtained also agreed with the increasing availability of NH3 in the rumen fluid, which would improve microbial growth through production of VFAs. Jamarun et al.10 and Febrina et al.11 reported that increasing the number of rumen microbial cells would increase the production of VFAs, thereby improving rumen microbial growth. The VFA content in the rumen fluid can indicate the feed fermentability level, where the higher the feed fermentability level is, the greater the VFA content41.
In this study, the average total VFA content was sufficient for maximum rumen microbial growth. Waldron et al.42 stated that the total VFA content in rumen fluid was 60-120 mM. The optimum VFA content for rumen microbes is 80-160 mM37.
pH: Table 4 shows that the average pH of the rumen fluid ranged from 6.63-6.86. Based on analysis of variance (ANOVA), the significant effect (p<0.01) was shown to be lowest in T1. This is because the VFA content in T1 was higher than that in the other treatments. High VFA contents will automatically decrease the rumen fluid pH. In accordance with the opinion of Sutardi43, the higher the VFA content is, the more organic acids will be produced, causing the pH of the rumen fluid to be low.
The pH of the rumen fluid in this study was sufficient for the growth of rumen microbes. According to Erdman44, the optimal pH range for CD is 6.4-7. If the pH of the rumen fluid drops below 6, microbial activity will be inhibited. Furthermore, Van Soest45 stated that pH greater than 7.1 can reduce the microbial population drastically so that the resulting energy is low. The conditions for optimal rumen microbial activity are a rumen pH of 6-6.946. The degree of acidity or pH of the rumen fluid is a balance between the buffer capacity and the basic or acidic properties of the fermentation product. The type of feed given to the livestock will affect the pH of the rumen.
Microbial protein synthesis: MPS describes the donation of proteins to rumen microbes. MPS in this study is shown in Fig. 1. In this study, the obtained MPS was 53.56-79.21 mg mL–1, which is similar to that (±68.21-72.93 mg mL–1) in a treatment combining the basal ration with additional rice bran, wet and dried onggok and corn. Some carbohydrate sources include fermentable carbohydrate groups that will be easily digested by rumen microbes47. However, Febrina et al.11 reported that the addition of the P, S and Mg to FOPF resulted in an unstable (p>0.05) MPS of 36.878-46.816 mg mL–1.
The highest MPS was present in T1, followed successively by T2, T3 and T4. The high MPS in T1 was caused by the feed CP content in T1 being is also higher than that in the other treatments (Table 2). The CP is a crucial component for MPS because CP indicates the availability of N for rumen microbes as long as the N concentration does not decrease and the CP is not used as a source of energy48. The increased rumen bacterial growth was also reflected in the increased MPS. Table 2 shows that the population of rumen bacteria was also highest in T1.
Optimal MPS requires the supply of N and other nutrients. N comes from the production of NH3, while other nutrients come from the production of VFAs as a result of carbohydrate fermentation.
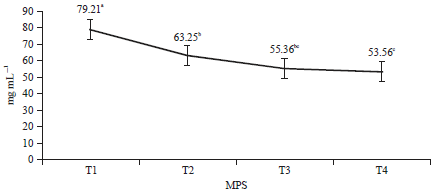 | |
| Fig. 1: | Microbial protein synthesis of OPF fermented by Phanerochaete chrysosporium in combination with tithonia and elephant grass |
| Means in the same line with different letters (a, b, c and d) are significant (p<0.01) | |
Table 1 and 2 show that T1 increased the production of VFAs, which was followed by a high number of microbes. This also indicates that MPS increases because of an increasing number of rumen microbes. The high MPS in T1 indicated carbohydrate degradation, which was faster than in other treatments. The breaking of lignocellulose and lignohemicellulose bonds was optimal in this treatment, increasing the amount of nutrients and making the carbohydrates in the rumen are more easily degraded by rumen microbes. The rumen microbial population will increase when nutrient availability meets the microbial requirements, thereby increasing MPS. The high MPS in T1 is suspected to be from amino acids from T. Microbial protein synthesis requires amino acids derived from feed protein degradation. The P content in T (16%) was also suspected to influence the increase in MPS in T1. The synthesis of microbial proteins will increase when the P content is sufficient49.
Hoover and Stokes50 suggested that the rate of MPS will increase with carbohydrate digestibility. MPS is low in low-quality forages due to slow carbohydrate degradation. This was seen in the T2, T3 and T4, whose rations contained more FOPF than did those of T1, indicating lower CD (Table 3). Low CD also produces lower MPS. OPF are a low-quality forage compared with EG and T because there is sufficient lignin content.
CONCLUSION
The combination of 20% FOPF+16% T and 64% EG (T1) yielded the best MPS, in vitro fermentability and digestibility values.
SIGNIFICANCE STATEMENT
The effect of FOPF combined with T and EG on MPS, in vitro fermentability and digestibility was evaluated in this study. The results indicated that the combination of 20% FOPF+16% T and 64% EG was highly effective in rations derived from plantation waste products, such as OPF. This study will help researchers to realize that the combination of FOPF with T and EG is important for supporting MPS, in vitro fermentability and digestibility. Thus, this research could lead to a new theory on the importance of feed material combinations in rations derived from plantation waste products.
ACKNOWLEDGMENTS
This research was funded by the Andalas University Professor Grant 2017 and is part of the Ph.D. dissertation of Roni Pazla in Animal Nutrition, Faculty of Animal Science, Andalas University.
REFERENCES
- Jamarun, N., M. Zein, Arief and R. Pazla, 2018. Populations of rumen microbes and the in vitro digestibility of fermented oil palm fronds in combination with Tithonia (Tithonia diversifolia) and elephant grass (Pennisetum purpureum). Pak. J. Nutr., 17: 39-45.
CrossRefDirect Link - Zain, M., J. Rahman and Khasrad, 2014. Effect of palm oil by products on in vitro fermentation and nutrient digestibility. Anim. Nutr. Feed Technol., 14: 175-181.
Direct Link - Imsya, E.B., K.G. Laconi, Wiryawan and Y. Widyastuti, 2013. Identification of phenolic compounds and its antioxidant activity from lignin palm oil frond fermented with Phanerochyte chrysosporium. Proceedings of the 4th International Conference on Sustainable Animal Agriculture for Developing Countries, July 23-31, 2013, University Lanzhou, China, pp: 310-312.
- De Koker, T.H., K.K. Nakasone, J. Haarhof, H.H. Burdsall Junior and B.J.H. Janse, 2003. Phylogenetic relationships of the genus Phanerochaete inferred from the internal transcribed spacer region. Mycol. Res., 107: 1032-1040.
CrossRefDirect Link - Shi, J., R.R. Sharma-Shivappa and M.S. Chinn, 2009. Microbial pretreatment of cotton stalks by submerged cultivation of Phanerochaete chrysosporium. Bioresour. Biotechnol., 100: 4388-4395.
CrossRefPubMedDirect Link - Singh, D. and S. Chen, 2008. The white-rot fungus Phanerochaete chrysosporium: Conditions for the production of lignin-degrading enzymes. Applied Microbiol. Biotechnol., 81: 399-417.
CrossRefDirect Link - Jamarun, N., M. Zain, Arief and R. Pazla, 2017. Effects of calcium, phosphorus and manganese supplementation during oil palm frond fermentation by Phanerochaete chrysosporium on laccase activity and in vitro digestibility. Pak. J. Nutr., 16: 119-124.
CrossRefDirect Link - Jamarun, N., M. Zain, Arief and R. Pazla, 2017. Effects of Calcium (Ca), Phosphorus (P) and Manganese (Mn) supplementation during oil palm frond fermentation by Phanerochaete chrysosporium on rumen fluid characteristics and microbial protein synthesis. Pak. J. Nutr., 16: 393-399.
CrossRefDirect Link - Febrina, D., N. Jamarun, M. Zain and Khasrad, 2016. The effects of P, S and Mg supplementation of oil palm fronds fermented by Phanerochaete chrysosporium on rumen fluid characteristics and microbial protein synthesis. Pak. J. Nutr., 15: 299-304.
CrossRefDirect Link - Jamarun, N., Elihasridas, R. Pazla and Fitriyani, 2017. In vitro nutrients digestibility of the combination Titonia (Tithonia difersivolia) and Napier grass (Pennisetum purpureum). Proceedings of the 7th International Seminar on Tropical Animal Production, September 12-14, 2017, Yogyakarta, Indonesia.
- Jama, B., C.A. Palm, R.J. Buresh, A. Niang, C. Gachengo, G. Nziguheba and B. Amadalo, 2000. Tithonia diversifolia as a green manure for soil fertility improvement in Western Kenya: A review. Agrofor. Syst., 49: 201-221.
CrossRefDirect Link - Fasuyi, A.O., F.A.S. Dairo and F.J. Ibitayo, 2010. Ensiling wild sunflower (Tithonia diversifolia) leaves with sugar cane molasses. Livest. Res. Rural Dev., Vol. 22, No. 3.
Direct Link - Jamarun, N., Elihasridas, R. Pazla and Fitriyani, 2017. In vitro nutrients digestibility and rumen fluid characteristics of the combination Titonia (Tithonia difersivolia) and napier grass (Pennisetum purpureum). Proceedings of the 3th Nasional Seminar on Cows and Buffalo, Oktober 4-5, 2017, Padang, Indonesia.
- Suryani, N.N., M. Budiasa, I. Ketut, A. Astawa and I. Putu, 2014. Fermentasi rumen dan sintesis protein mikroba kambing peranakan ettawa yang diberi pakan dengan komposisi hijauan beragam dan level konsentrat berbeda. Majalah Ilmiah Peternakan, Vol. 17, No. 2.
CrossRefDirect Link - Tilley, J.M.A. and R.A. Terry, 1963. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci., 18: 104-111.
CrossRefDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Rahayu, S., N. Jamarun, M. Zain and D. Febrina, 2015. Influence of mineral dosage of Ca and long fermentation of palm oil to lignin contents, digestibility of DM, OM, CP and fiber fraction (NDF, ADF, hemicellulosa and cellulose) using Phanerochaete chrysosporium Kapang. J. Peternakan Indonesia, 17: 151-162.
Direct Link - Elihasridas, 2012. The effect of supplementation of mineral Zinc on in vitro digestibility of amoniated corn cobs ration. J. Peternakan, 9: 9-14.
Direct Link - Oluwasola, T.A. and F.A.S. Dairo, 2016. Proximate composition, amino acid profile and some anti-nutrients of Tithonia diversifolia cut at two different times. Afr. J. Agric. Res., 11: 3659-3663.
CrossRefDirect Link - Hernaman, I., T. Toharmat, W. Manalu and P.I. Pudjiono, 2007. [Study on Zn-fitat processing and its degradation in rumen fluid]. J. Indon. Trop. Anim. Agric., 32: 139-145, (In Indonesian).
Direct Link - Bravo, D., D. Sauvant, C. Bogaert and F. Meschy, 2003. II. Quantitative aspects of phosphorus absorption in ruminants. Reprod. Nutr. Dev., 43: 271-284.
Direct Link - Rodehutscord, M., H. Heuvers and E. Pfeffer, 2000. Effect of organic matter digestibility on obligatory faecal phosphorus loss in lactating goats, determined from balance data. Anim. Sci., 70: 561-568.
Direct Link - Febrina, D., N. Jamarun, M. Zain and Khasrad, 2017. Digestibility of goat rations containing fermented oil palm fronds by Phanerochaete chrysosporium supplemented with phosphorus, sulfur and magnesium. J. Biol. Sci., 17: 298-304.
CrossRefDirect Link - Zain, M., N. Jamarun and A.S. Tjakradidjaja, 2010. Phosphorus supplementation of ammoniated rice straw on rumen fermentability, synthesised microbial protein and degradability in vitro. World Acad. Sci. Eng. Technol., 4: 357-359.
Direct Link - Zain, M., N. Jamarun and Zulkarnaini, 2010. Effect of phosphorus and sulphur supplementation in growing beef cattle diet based on rice straw ammoniated. Asian J. Scient. Res., 3: 184-188.
CrossRefDirect Link - Lu, C.D., J.R. Kawas and O.G. Mahgoub, 2005. Fibre digestion and utilization in goats. Small Rumin. Res., 60: 45-52.
CrossRefDirect Link - Suyitman, L. Warly, A. Rachmat and D.R. Ramadhan, 2015. Effect of minerals S, P and cassava flour leaf supplemented with ammoniation palm leaves on the performance of beef cattle. Pak. J. Nutr., 14: 849-853.
CrossRefDirect Link - Paengkoum, P., J.B. Liang, Z.A. Jelan and M. Basery, 2006. Utilization of steam-treated oil palm fronds in growing saanen goats: II. Supplementation with energy and urea. Asian-Aust. J. Anim. Sci., 19: 1623-1631.
CrossRefDirect Link - Muhktarudin and Liman, 2006. Penentuan tingkat penggunaan mineral organik untuk memperbaiki bioproses rumen pada kambing secara in vitro. J. Ilmu-Ilmu Peternakan Indonesia, 8: 132-140.
Direct Link - Febrina, D., N. Jamarun, M. Zain and Khasrad, 2016. Effects of calcium (Ca) and manganese (Mn) supplementation during oil palm frond fermentation by Phanerochaete chrysosporium on in vitro digestibility and rumen fluid characteristics. Pak. J. Nutr., 15: 352-358.
CrossRefDirect Link - Waldron, M.R., F.N. Schrick, J.D. Quigley, J.L. Klotz, A.M. Saxton and R.N. Heitmann, 2002. Volatile fatty acid metabolism by epithelial cells isolated from different areas of the ewe rumen. J. Anim Sci., 80: 270-278.
PubMedDirect Link - Erdman, R.A., 1988. Dietary buffering requirements of the lactating dairy cow: A review. J. Dairy Sci., 71: 3246-3266.
CrossRefDirect Link - van Soest, P.J., 1994. Nutritional Ecology of the Ruminant. 2nd Edn., Cornell University Press, Ithaca, New York, Pages: 476.
Direct Link - Hindratiningrum, N., M. Bata and S.A. Santosa, 2011. Produk fermentasi rumen dan produksi protein mikroba sapi lokal yang diberi pakan jerami amoniasi dan beberapa bahan pakan sumber energi. J. Agripet, 11: 29-34.
CrossRefDirect Link - Gosselink, J.M.J., C. Poncet, J.P. Dulphy and J.W. Cone, 2003. Estimation of the duodenal flow of microbial nitrogen in ruminants based on the chemical composition of forages: A literature review. Anim. Res., 52: 229-243.
CrossRefDirect Link - Karsli, M.A. and J.R. Russell, 2001. Effects of some dietary factors on ruminal microbial protein synthesis. Turk. J. Vet. Anim. Sci., 25: 681-686.
Direct Link - Hoover, W.H. and S.R. Stokes, 1991. Balancing carbohydrates and proteins for optimum rumen microbial yield. J. Dairy Sci., 74: 3630-3644.
CrossRefPubMedDirect Link