
S. Hartini
Faculty of Animal Husbandry, Papua University, Jalan Gunung Salju Amban, Manokwari 98314, Papua Barat, Indonesia
P. Purwaningsih
Faculty of Animal Husbandry, Papua University, Jalan Gunung Salju Amban, Manokwari 98314, Papua Barat, Indonesia
Pakistan Journal of Nutrition
Year: 2017 | Volume: 16 | Issue: 4 | Page No.: 227-235







ABSTRACT
Background and Objective: It is possible that a commercial broiler ration that contains many fleas/insects and excess fine feed may have poor nutrient quality. This study aimed to investigate the effects of Wheat Pollard (WP), Rice Hull (RH), phytase and cellulase supplementation in a commercial broiler diet on the growth performance, carcass weight and gut characteristics of broiler chickens from 0-35 days of age. Methodology: A total of 175 male broiler chicks of 1 day old (Lohmann) were fed using 7 dietary treatments (5 replicates treatment–1). The diets used were as follows: (1) A commercial broiler diet as a control (C) diet, (2) C+WP (CWP)+phytase, (3) CWP+cellulase, (4) CWP+phytase+cellulase, (5) C+RH (CRH)+phytase, (6) CRH+cellulase and (7) CRH+phytase+cellulase. Wheat pollard and RHs were added at 40 g kg–1 of diet. Phytase was added at 1250 FTU kg–1 and cellulase was added at 250 U kg–1. Results: The control diet contained a low level of Ca (0.5%) and total P (0.4%). Treatment diets did not affect the growth performance of broiler chickens (p>0.05). Birds fed diets 5, 6 and 7 exhibited higher carcass weights (p<0.05), the lightest duodenum and jejunum weights (p<0.01), the lowest jejunum pH values (p<0.05) and the lowest cecal pH values (p<0.01), as well as higher cecal acetic and butyric acid concentrations (p<0.01). The duodenal digesta was the highest in birds fed the control diet (p<0.05). Conclusion: Adding 40 g kg–1 of RHs to broiler diets containing low levels of Ca (0.5%) and total P (0.3-0.5%) enhanced carcass weights. The increase in carcass weights was due to a reduction in gut weights and the addition of RHs, partly because of their physical structure, contributed more to this than phytase supplementation. Cellulase supplementation did not enhance phytase efficacy.
PDF Abstract XML References Citation
Received: November 21, 2016;
Accepted: February 13, 2017;
Published: March 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
S. Hartini and P. Purwaningsih, 2017. Effects of Adding Insoluble Non-starch Polysaccharides and Exogenous Enzymes to a Commercial Broiler Diet on the Growth Performance and Carcass Weight of Broiler Chickens. Pakistan Journal of Nutrition, 16: 227-235.
DOI: 10.3923/pjn.2017.227.235
URL: https://scialert.net/abstract/?doi=pjn.2017.227.235
DOI: 10.3923/pjn.2017.227.235
URL: https://scialert.net/abstract/?doi=pjn.2017.227.235
INTRODUCTION
In developing countries such as Indonesia, broiler farmers who live in small towns or villages often face the fact that the broiler rations they purchase are "old". Broiler rations in this condition usually contain many fleas/insects and their structure is no longer intact1. Moreover, a broiler finisher ration in pellet form becomes a powder. Because of this, it is suspected that some types of rations have poor nutrient quality. The majority of feed ingredients used in poultry diets are derived from plant cereals such as soybean, corn, wheat, or barley2. Anti-nutritional factors and indigestible components in plants can be a factor that can affect the quality of a formulated diet. The phytate in plant material is a nutritional obstacle for monogastric animals3. Some researchers have indicated that the supplementation of broiler feeds with phytase can alleviate the effects of anti-nutritional factors and improve bird performance4,5. The use of exogenous phytase supplementation has been reported to improve the use of phytate phosphorus (P)6,7, which represents between 50 and 80% of the total P content in cereals8.
In addition to enzymes, the benefit of insoluble non-starch polysaccharides (iNSPs) in broiler diets has also been reported in many studies. Many of the benefits have been associated with improving gut health9-11 and gizzard function12 and increasing starch digestibility13 and growth performance in broilers14,15 and layers16.
Lately, there has been an interest in combining phytase with cell-wall-degrading enzymes in broiler diets based on the assumption that the phytate hidden in the plant cell wall cannot be degraded by phytase. The use of phytase in combination with exogenous carbohydrase has been investigated in many studies17-19. It has been reported that the combination of phytase and carbohydrase is effective in improving broiler growth performance17,18.
The inclusion of iNSPs and/or enzyme supplements in broiler diets are common in poultry research but minimal data are available on the addition of iNSPs and enzymes to commercial broiler diets. In a previous study, Hartini and Massora11 tried to add graded levels (20, 40 and 60 g kg–1) of iNSPs Wheat Pollard (WP) and Rice Hull (RH)) to commercial broiler diets. They found that the addition of 40 g kg–1 of iNSPs decreased cecal microbial counts in young broilers. Moreover, the addition of 40 g kg–1 of RHs and 20 g kg–1 of WP reduced the total number of Escherichia coli and Enterobacter, respectively, in the small intestine as a source of iNSPs, WP contains 33.6% Dry Matter (DM) as iNSPs and 1.7% soluble NSPs20. The RHs are a by product of rice processing and contain mostly cellulose21.
The current study was designed to evaluate the effects of using exogenous enzymes (phytase and cellulase) and the addition of iNSPs (WP and RH) to a commercial broiler diet on the growth performance, carcass weight and gastrointestinal tract (GIT) characteristics of broilers from 0-35 days of age. The hypothesis for this study was that the addition of exogenous enzymes and iNSPs to commercial broiler diets could improve the nutritive values of the diets and consequently increase broiler performance. However, the degradation of nutrients in the commercial diets used was also considered as a factor that may influence the results of adding iNSPs and enzymes.
MATERIALS AND METHODS
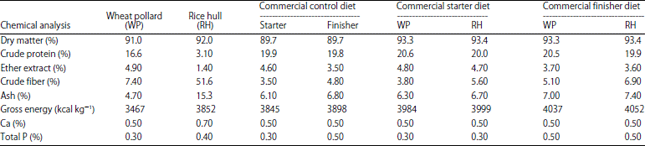
Experimental diets and fiber sources: The commercial broiler diet used in this study was one of the commercial broiler diets available on the Indonesian market and has characteristics that have been described previously. The WP and RHs were used as sources of iNSPs. The RHs were ground using a grinder fit with a 2 mm screen. The WP and RHs were added to the commercial starter and finisher diets at 40 g kg–1, an amount similar to that in the study by Hartini and Massora11. The commercial broiler diets, WP and RHs were analyzed for DM, crude protein (CP), ether extract (EE), crude fiber (CF), gross energy (GE), calcium (Ca) and phosphorus (P). Dry matter, CP, EE and CF were analyzed using standard AOAC methods, GE was analyzed via bomb calorimetry, Ca was analyzed using the Atomic Absorption Spectrophotometry (AAS) method and P was analyzed using a colorimetric method. All of the chemical analyses were performed in a laboratory at the Nutrition and Food Study Center, Gadjah Mada University, Yogyakarta, Indonesia. The analyzed and calculated nutrient values of the experimental diets are shown in Table 1.
The phytase used was derived from E. coli (Quantum Blue, ABVista Feed Ingredients, Marlborough, UK). The standard recommended level of phytase was 100 g t–1 to obtain an activity of 500 FTU kg–1. The cellulase used was an SQzyme CSP product at 20,000 U g–1 from Suntaq International Limited, Shenzhen, China.
Diets used in the experiment were as follows:
| • | Commercial broiler diets (corn-soybean based) as the control diet (C) |
| • | C+40 g kg–1 of WP (CWP)+1250 FTU kg–1 of phytase |
| • | CWP+250 U kg–1 of cellulase |
| • | CWP+enzyme complex (1250 FTU kg–1 of phytase and 250 U kg–1 of cellulase) |
| • | C+40 g kg–1 of RHs (CRH)+1250 FTU kg–1 of phytase |
| • | CRH+250 U kg–1 of cellulase |
| • | CRH+enzyme complex (1250 FTU kg–1 of phytase and 250 U kg–1 of cellulase) |
| Table 1: | Chemical analyses of fiber sources and commercial diets and calculation analyses of basic experimental diets before addition of exogenous enzymes |
 | |
Bird management: One hundred seventy five 1 day old male broiler chicks (Lohmann) were randomly allocated to 35 floor pens, with 5 birds pen–1 and 5 pens treatment–1. Flock uniformity was determined by calculating the coefficient of variance (CV) for body weight (BW). The birds were fed a starter diet with the addition of 40 g kg–1 of iNSPs (WP or RHs) and exogenous enzymes (1250 FTU kg–1 of phytase and/or 250 U kg–1 of cellulase) until 21 days of age; then, they were fed a finisher diet with the same supplements from 22 days until the end of experiment at 35 days. Both feed and water were offered ad libitum during the experiment. Pens were illuminated 24 h day–1.
Variables measured: Variables measured in the experiment were BW gain (BWG) (gram/bird/day), feed intake (FI) (g/b/d), Feed Conversion Efficiency (FCE) (g/g), relative weight of digestive organs (gizzard, duodenum, jejunum, ileum and caeca) expressed as g/100 g b.wt., relative length of digestive organs (duodenum, jejunum and ileum) expressed as cm/100 g b.wt. pH in digestive organs (duodenum, jejunum, ileum and caeca), carcass weight (% of live body weight) and cecal volatile fatty acids (VFAs) (acetate, propionate and butyrate). Body weight gain and FI were determined at the beginning and end of the experiment. Body weight gain and FI were used to calculate FCE feed (FCE = BWG/FI) during the entire experimental period. Mortality was recorded daily.
Sample collection and analyses: At the end of the experiment (35 days), two birds from each of the 5 replicates treatment–1 were selected based on the similarity of their weight to the average bird weight per cage. The birds were fasted for approximately 8 h before being slaughtered. After determining their live weight, the birds were slaughtered by cutting the jugular vein and they were used to measure the variables below.
Carcass: The broiler carcass was eviscerated, the neck with the skin was cut off at the tip of the shoulders and the shanks were removed at the hocks. Carcass weight was calculated as a percentage of live body weight (g/100 g b.wt.).
Cecal volatile fatty acids: The VFA levels in cecal digesta were determined via gas chromatography using the method described by Hartini22. Three grams of sample were diluted with 3 mL of 0.1 M H2SO4 and thoroughly mixed. The sample was then centrifuged (15,000×g for 15 min) and 0.1 mL of a reference VFA (caproic acid) was added to a 1 mL aliquot of the supernatant. The VFAs were distilled using thundberg tubes. The concentration of VFAs was quantified via Gas Liquid Chromatography (GLC).
Gastrointestinal tract length, weight, digesta and pH: The GIT was removed and cut into the following segments: Gizzard, duodenum (from gizzard to pancreo-biliary ducts), jejunum (measured from the end of duodenal loop to Meckel’s diverticulum), ileum (from Meckel’s diverticulum to the ileocecocolic junction) and ceca. The length of the duodenum, jejunum and ileum were measured to the nearest mm. Next, the duodenum, jejunum, ileum, gizzard and ceca were weighed prior to and after the removal of their contents. The pancreas was also removed and weighed. Digesta contents of the duodenum, jejunum, ileum and ceca were measured and then put in 50 mL glass beakers. Distilled water was added at a 1:1 ratio, except for cecal digesta, which was diluted with distilled water at a ratio of 1:5. The mixture was stirred until homogenized and a pH meter was inserted. The pH was read when the number was stable.
Statistical analyses: The data obtained were analyzed statistically using a one-way analysis of variance23. If F tests were significant, Duncan’s multiple range tests were used to test for differences among group means. Statistical significance was accepted at p<0.05. For differences among means, 0.05<p<0.10 were accepted as representing tendencies and differences.
RESULTS
Fiber sources and experimental diets: Table 1 shows the nutrient values of the fiber sources and the experimental diets used in this study. Rice hulls contained almost 7 times more crude fiber than WP. The addition of RHs in the starter and finisher diets resulted in higher crude fiber contents than the addition of WP. The starter and finisher diets contained low concentrations of Ca and total P and these concentrations did not differ among diets.
Growth performance and carcass weight and gastrointestinal tract pH and weight: The effects of the diets on performance, carcass weight, GIT pH and GIT weight are summarized in Table 2. The diets did not have significant effects on BWG, FI and FCE (p>0.05); however, diet did affect carcass weights (p<0.05). The addition of RHs with a single enzyme or a combination of enzymes resulted insignificantly higher carcass weights than were observed in the other treatment groups. Supplementation of WP with phytase and cellulase resulted in higher carcass weights than the addition of WP with either phytase or cellulase alone (Table 2). There was no mortality during the experiment.
Table 2 shows that CRH diets supplemented with phytase, cellulase, or with phytase and cellulase significantly lowered the jejunal pH (p<0.05) and cecal pH (p<0.01), which did not occur in groups fed the other treatment diets. Among the CRH diets, supplementation of an enzyme combination (phytase and cellulase) resulted in the lowest jejunal pH. The duodenal pH of birds fed CRH diets with the addition of an enzyme combination (phytase and cellulase) also tended to be lower than that of birds fed the other diets (p = 0.095). Ileal pH was not affected by the diets (p>0.05) (Table 2).
The weights of the gizzard, duodenum, jejunum, ileum, ceca and pancreas are expressed per 100 g b.wt. (g/100 g b.wt.). Birds fed the control diet had significantly higher (p<0.01) duodenum and jejunum weights compared with those fed the other diets (Table 2). Supplementing the CWP and CRH diets with enzymes reduced duodenum weights (p<0.01). Birds fed CRH diets with the addition of phytase or with the addition of phytase and cellulase had the lightest duodenal and jejunal weights (p<0.01) (Table 2).
Gastrointestinal tract length, digesta content and cecal VFAs: The effect of iNSP and enzyme additions on GIT length, digesta content and cecal VFA levels are shown in Table 3. The length of the duodenum, jejunum and ileum are expressed per 100 g b.wt. (cm/100 g b.wt.). Diets had no effect on the length of the GIT (p>0.05).
Diets affected duodenal contents (p<0.05) but the effect on jejunal and cecal contents only tended to be different (p = 0.08 and p = 0.07, respectively) (Table 3). Birds fed the control diets had higher duodenal contents than those fed the other diets (p<0.05).
| Table 2: | Responses of performance, carcass weight (%) and gastrointestinal tract (GIT) weight (g/100 g b.wt.) to diet type |
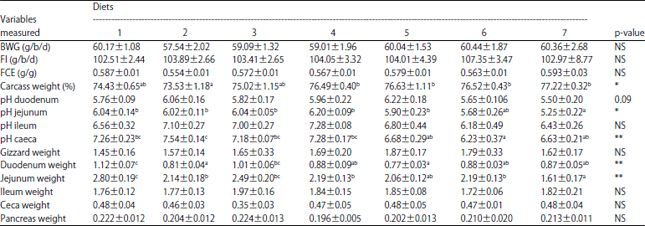 | |
*p<0.05, **p<0.01, NS: Not significant, diets 1: Commercial diet as a control (C) diet, 2: C with 40 g kg–1 of WP (CWP) and 1250 FTU kg–1 of phytase, 3: CWP with 250 U kg–1 of cellulase, 4: CWP with 1250 FTU kg–1 of phytase and 250 U kg–1 of cellulase, 5: C with 40 g kg–1 of RHs (CRH) and 1250 FTU kg–1 of phytase, 6: CRH with 250 U kg–1 of cellulase, 7: CRH with 1250 FTU kg–1 of phytase and 250 U kg–1 of cellulase, abcMean values within a row bearing different superscripts differ significantly | |
| Table 3: | Effect of diets on GIT length (duodenum, jejunum and ileum) (cm/100 g b.wt.) and GIT digesta (gizzard, duodenum, jejunum, ileum and ceca) (g/100 g b.wt.) and cecal volatile fatty acids (acetic, propionic and butyric) (μM mL–1) |
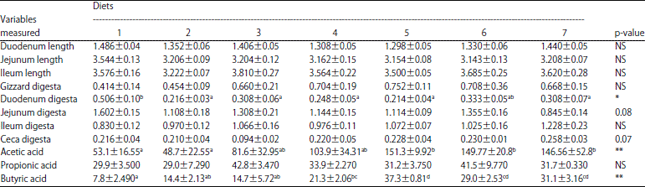 | |
*p<0.05, **p<0.01, NS: Not significant, diets 1: Commercial diet as a control (C) diet, 2: C with 40 g kg–1 of WP (CWP) and 1250 FTU kg–1 of phytase, 3: CWP with 250 U kg–1 of cellulase, 4: CWP with 1250 FTU kg–1 of phytase and 250 U kg–1 of cellulase, 5: C with 40 g kg–1 of RHs (CRH) and 1250 FTU kg–1 of phytase, 6: CRH with 250 U kg–1 of cellulase, 7: CRH with 1250 FTU kg–1 of phytase and 250 U kg–1 of cellulase, abcMean values within a row bearing different superscripts differ significantly | |
Whereas jejunal contents tended to be higher in birds fed the control diet (p = 0.08), cecal contents tended to be higher in those fed the CRH diet supplemented with the enzyme complex (phytase and cellulase) (p = 0.07) (Table 3).
Cecal VFAs, except propionate were highly affected by the diets (p<0.01) (Table 3). The addition of RHs and enzymes increased the concentrations of cecal acetic and butyric acids more than was observed for the other diets (p<0.01). Among the CRH diets, cellulase supplementation resulted in a higher concentration of butyric acid than was observed with phytase or combined phytase and cellulase supplementation. Birds fed the control diets had the lowest concentrations of cecal acetic and butyric acids (Table 3).
DISCUSSION
The reduction in nutrient quality in the control diets in this study was predicted. However, the low concentrations of Ca and P in these diets was not expected. The growth of broilers requires sufficient amounts of Ca and P for the formation and maintenance of their skeleton. Phytase supplementation significantly increases Ca digestibility regardless of the Ca and P levels of the diets24 and significantly increased body weight. The inclusion of iNSPs in broiler diets has been reported to improve growth performance of broiler chickens14,15.
In the current study, BWG, FI and FCE were not affected by the addition of 4% WP or 4% RHs and exogenous enzymes. Insoluble non-starch polysaccharide inclusion (3% oat hulls or 3% soy hulls) in corn based-diets has been shown to improve BWG and feed conversion efficiency in young broiler chicks14. However, the inclusion of 4% oat hulls25 or 10% cellulose26 in wheat-based diets was found to only increase FI but not BWG or FCR, in broilers at 21 days of age. The different results found indicate that the effect of the iNSP source not only depends on the physicochemical properties of the iNSP source per se but also on the interaction between the individual iNSP sources and other cell wall components in the diet27. Supplementation with phytase or cellulase or with both phytase and cellulase in this study did not show any significant effect. In contrast, a previous study by Meng et al.28 demonstrated that multiple enzyme preparations statistically improved iNSP digestibility. The effect of phytase has been shown to be significant in increasing broiler weight gain when phytase is added to a P-deficient diet7,19. Biehl and Baker29 observed that supplementation with a high dose of microbial phytase increased FCE when the diet was deficient in amino acids. Furthermore, Avila et al.18 found that the use of the multiple-enzyme complex with typical commercial sorghum-SBM broiler diets reformulated at -85 kcal kg–1 of AMEn, -1.5% CP, -1.5% amino acids, -0.153% of available P and -0.12% of Ca resulted in broiler performance equivalent to that of those fed the control diet.
The diets in the current study contained 0.5% Ca and 0.3% P during the starter period and 0.5% Ca and 0.5% P during the finisher period. In Indonesia, the standard nutrient requirements of meat-type broiler chickens during the starter period (0-3 weeks of age) are 19-23% CP, 0.9-1% Ca and 0.6-1% total P, whereas during the finisher period (>3-6 weeks of age), the requirements are 18-20% CP, 0.9-1.2% Ca and 0.6-1% total P30. The concentration of Ca and P in the present study were below the standard requirements. In broiler starter diets, the concentrations of Ca and P were reduced by approximately 44% each, whereas in the broiler finisher diets, the concentrations of Ca and P were reduced by approximately 44 and 17%, respectively. The exact cause of Ca and P reduction is unknown.
The addition of iNSPs (WP or RHs) and exogenous enzymes (phytase and/or cellulase) did not enhance growth performance. In broiler diets, Ca and P are essential minerals for growth and a deficiency or excess of one may interfere with the proper utilization of the other. Therefore, in the formulation of feeds, Ca and P should be included in a definite relationship for bone formation in birds. The SNI30 recommended a Ca-P ratio of approximately 1.5:1 for broilers based on total P. Calcium and P and particularly the Ca:P ratio, influence phytase activity31. In their review, Bedford et al.32 included Ca and P as major determinants of phytase activity. Tamim et al.33 demonstrated that dietary Ca can precipitate with phytate P, forming insoluble phytate complexes. The addition of phytase at either Ca level (0.18 or 0.68% Ca) resulted in an increase in the apparent absorption of Ca and P in the GIT of broiler chickens. Delezie et al.31 concluded that Ca and P concentrations can be reduced by 20% if done in a balanced way and the supplementation of phytase in this condition will further improve broiler growth. Adding graded levels of Ca and phytase in a diet deficient in Ca but adequate in total P was found to improve weight gain in young broiler chicks34, suggesting that the addition of Ca causes a significant improvement in weight gain and the addition of phytase causes more Ca to be available, which significantly increases body weight. In the current study, although a high dosage of phytase (1250 FTU kg–1) was used, the reduced concentrations of both Ca and P, by approximately 44%, from the normal requirements during the starter period were more likely to constrain optimal phytase activity. Even at normal Ca concentrations, the phytate P liberated by phytase may not be sufficient to meet broiler’s P requirements if the diet has a low P concentration31. Moreover, growth rates of broilers are relatively high during the starter period (1-21 days), indicating that the demand for Ca and P during this period is also higher than during the finisher period. The reduction in Ca and P concentrations during the starter period was assumed to greatly affect growth performance. The addition of cellulase in diets in the current study did not result in any differences in BWG, which indicates that cellulase supplementation failed to increase phytase efficacy. The GE content of the diets in the present study ranged from 3845-3999 kcal kg–1 for the starter diets and 3898-4052 kcal kg–1 for the finisher diets. The fact that FI was found to be similar among the dietary treatment groups implies that the contributions of iNSPs and enzymes in the feed did not affect energy availability, which is supported by the fact that birds have been shown to compensate by increasing FI when diets are low in energy35,36.
Carcass weights were found to be higher in broiler chickens fed the CRH diets with the addition of phytase or phytase and cellulase than in those fed the other diets. The broiler carcass weight is a bird's body weight without blood, feathers, head, legs and visceral organs. The lighter weights of the duodenum and jejunum and the duodenal content of birds fed the CRH diets plus phytase, cellulase or phytase and cellulase might be responsible for the increase in carcass weights. Rolls et al.37 reported that modifications to gut morphology are associated with an increase in gut microflora. Cereal by-products contain little water-soluble NSPs but contain high concentrations of iNSPs38. Insoluble β-glucans pass into the large intestine, where they are fermented by bacteria39. Significant levels of disaccharides, low-molecular weight polysaccharides and oligosaccharides derived from either water-soluble or insoluble NSPs resulting from the use of exogenous enzymes may be fermented in the ceca40. Large numbers of microorganisms in the caeca tend to migrate to the gut where nutrient absorption occurs, causing the lining of the gut to become thicker41. Supplementation with enzymes may reduce the microbial influence on the digestive tract of birds, possibly by reducing fermentative substrate levels42,43 and consequently reducing gut weight. We only agree in part with this idea. The differing results for the effects of the addition of WP and enzymes on carcass weight imply that there was another factor besides enzymes that influenced the increase in carcass weight. A previous study by Hartini and Massora11 found that the addition of 40 g kg–1 of iNSPs (RHs or WP) in commercial broiler diets without enzyme supplementation significantly reduced the total number of microbes in the small intestine. Therefore, it seems that iNSPs have an effect by decreasing microbial counts and the effects of the enzyme was the only addition to it.
Jozefiak et al.44 demonstrated that supplementation with oligosaccharides increases fermentation in ceca and consequently increases the production of VFAs (acetate, propionate and butyrate). In the present study, the addition of RHs, regardless of the enzymes added, resulted in a greater production of acetic and butyric acids than the addition of WP. Different fiber sources affect VFA production in different segments of the GIT45. He et al.46 also demonstrated that different fiber sources resulted in different VFA profiles, especially in the gizzard and ceca of growing Greylag geese. Butyric acid has been reported to reduce the pH of the small intestine and has anti-bacterial properties47. Indeed, in the current study, birds fed the control diets with the addition of RHs, regardless of the enzymes added, showed a reduced pH in their jejunum and ceca. This may also explain the lower amounts of duodenal and jejunal digesta in birds fed the control diets with the addition of RHs and enzymes.
Moreover, Jamroz et al.48 reported that feeding geese a large proportion (50%) of rye resulted in an increase in the thickness of their intestinal wall, while feeding geese rapeseed meal (20%) decreased the thickness of their intestinal wall. The addition of RHs in the present study was only 4%. Therefore, it is possible that the level of RHs in the diet and the physical structure of RHs might affect reductions in intestinal wall thickness and consequently reduce gut weights. Montagne et al.49 reported that the effect of dietary fiber on epithelial morphology and cell turnover is variable and depends on the physicochemical characteristics of the fiber, the level of incorporation into the diet and the duration of ingestion.
Hetland and Svihus25 suggested that a high-fiber diet tends to stay in the gizzard longer. The effect of diet on gizzard weight in the current study was not significant. The addition of 4% iNSPs may not be sufficient to stimulate gizzard function as the activity of the gizzard depends on its contents13. However, in birds fed commercial broiler diets with the addition of RHs and enzymes, the lower amounts of duodenal and jejunal digesta indicate that the presence of RHs actually slowed the rate of passage of digesta relative to rates in birds fed the other diets.
CONCLUSION
Based on these results, it can be concluded that adding 40 g kg–1 of iNSPs and enzymes (phytase or both phytase and cellulase) to commercial broiler diets containing low concentrations of Ca (0.5%) and P (0.3-0.5%) does not affect the growth performance of broiler chickens but does affect carcass weight. The increase in carcass weight was due to the reduction in gut weight and the addition of RHs, partly due to their physical structure, contributed more than to this than phytase supplementation. Cellulase supplementation did not enhance phytase efficacy.
SIGNIFICANCE STATEMENTS
This study demonstrates the possible effect of RH supplementation in decreasing the thickness of the intestinal wall, which can be beneficial for increasing the carcass weights of broiler chickens. This study will help researchers to determine the importance of RHs in influencing epithelial morphology and cell turnover, which many researchers are not aware of. Thus, new findings regarding the importance of using RHs in broiler diets may be discovered based on the results presented here.
ACKNOWLEDGMENTS
The experimental study presented here was made possible through the financial assistance provided by the Directorate General of Higher Education through DIPA DP2M No. 023.04.1.673453/2015, November 14, 2014 and contract No. 150/SP2H/PL/DIT. LITABMAS/11/2015, April 19, 2015, under the BOPTN research funding, Hibah Bersaing study.
REFERENCES
- Retnani, Y., E.D. Putra and L. Herawati, 2011. The effect of different water spraying level and storage period on endurance of pellet broiler finisher. J. Agripet, 11: 10-14.
CrossRefDirect Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Rev. Edn., National Academy Press, Washington, DC., USA., ISBN-13: 978-0309048927, Pages: 176.
Direct Link - Cabahug, S., V. Ravindran, P.H. Selle and W.L. Bryden, 1999. Response of broiler chickens to microbial phytase supplementation as influenced by dietary phytic acid and non-phytate phosphorus contents. I. Effects on bird performance and toe ash. Br. Poult. Sci., 40: 660-666.
CrossRefPubMedDirect Link - Shirley, R.B. and H.M. Edwards Jr., 2003. Graded levels of phytase past industry standards improves broiler performance. Poult. Sci., 82: 671-680.
PubMedDirect Link - Selle, P.H., V. Ravindran, G. Ravindran and W.L. Bryden, 2007. Effects of dietary lysine and microbial phytase on growth performance and nutrient utilisation of broiler chickens. Asian-Aust. J. Anim. Sci., 20: 1100-1107.
CrossRefDirect Link - Baidoo, S.K., Q.M. Yang, and R.D. Walker, 2003. Effects of phytase on apparent digestibility of organic phosphorus and nutrients in maize-soya bean meal based diets for sows. Anim. Feed Sci. Technol., 104: 133-141.
CrossRefDirect Link - Cowieson, A.J., T. Acamovic and M.R. Bedford, 2006. Supplementation of corn-soy-based diets with an Escherichia coli-derived phytase: Effects on broiler chick performance and the digestibility of amino acids and metabolizability of minerals and energy. Poult. Sci., 85: 1389-1397.
CrossRefPubMedDirect Link - Israel, D.W., P. Kwanyuen and J.W. Burton, 2006. Genetic variability for phytic acid phosphorus and inorganic phosphorus in seeds of soybeans in maturity groups V, VI and VII. Crop Sci., 46: 67-71.
Direct Link - Bao, Y.M. and M. Choct, 2010. Dietary NSP nutrition and intestinal immune system for broiler chickens. World's Poult. Sci. J., 66: 511-517.
CrossRefDirect Link - Mateos, G.G., E. Jimenez-Moreno, M.P. Serrano and R.P. Lazaro, 2012. Poultry response to high levels of dietary fiber sources varying in physical and chemical characteristics. J. Applied Poult. Res., 21: 156-174.
CrossRefDirect Link - Svihus, B., 2011. The gizzard: Function, influence of diet structure and effects on nutrient availability. World's Poult. Sci. J., 67: 207-224.
CrossRefDirect Link - Hetland, H., M. Choct and B. Svihus, 2004. Role of insoluble non-starch polysaccharides in poultry nutrition. World's Poult. Sci. J., 60: 415-422.
CrossRefDirect Link - Gonzalez-Alvarado, J.M., E. Jimenez-Moreno, R. Lazaro and G.G. Mateos, 2007. Effect of type of cereal, heat processing of the cereal and inclusion of fiber in the diet on productive performance and digestive traits of broilers. Poult. Sci., 86: 1705-1715.
CrossRefDirect Link - Jimenez-Moreno, E., A. de Coca-Sinova, J.M. Gonzalez-Alvarado and G.G. Mateos, 2016. Inclusion of insoluble fiber sources in mash or pellet diets for young broilers. 1. Effects on growth performance and water intake. Poult. Sci., 95: 41-52.
CrossRefPubMedDirect Link - Hartini, S., M. Choct, G. Hinch, A. Kocher and J.V. Nolan, 2002. Effects of light intensity during rearing and beak trimming and dietary fiber sources on mortality, egg production and performance of ISA brown laying hens. J. Applied Poult. Res., 11: 104-110.
CrossRefDirect Link - Cowieson, A.J. and O. Adeola, 2005. Carbohydrases, protease and phytase have an additive beneficial effect in nutritionally marginal diets for broiler chicks. Poult. Sci., 84: 1860-1867.
CrossRefPubMedDirect Link - Avila, E., J. Arce, C. Soto, F. Rosas, M. Ceccantini and D.R. McIntyre, 2012. Evaluation of an enzyme complex containing nonstarch polysaccharide enzymes and phytase on the performance of broilers fed a sorghum and soybean meal diet. J. Applied Poult. Res., 21: 279-286.
CrossRefDirect Link - Karimi, A., C. Coto, F. Mussini, S. Goodgame and C. Lu et al., 2013. Interactions between phytase and xylanase enzymes in male broiler chicks fed phosphorus-deficient diets from 1 to 18 days of age. Poult. Sci., 92: 1818-1823.
CrossRefPubMedDirect Link - Choct, M., 1997. Feed non-starch polysaccharides: Chemical structures and nutritional significance. Feed Milling Int., 7: 13-26.
Direct Link - Valchev, I., V. Lasheva, T. Tzolov and N. Josifov, 2009. Silica products from rice hulls. J. Univ. Chem. Technol. Metallurgy, 44: 257-261.
Direct Link - Paiva, D., C. Walk. and A. McElroy, 2004. Dietary calcium, phosphorus and phytase effects on bird performance, intestinal morphology, mineral digestibility and bone ash during a natural necrotic enteritis episode. Poult. Sci., 93: 2752-2762.
CrossRefPubMedDirect Link - Hetland, H. and B. Svihus, 2001. Effect of oat hulls on performance, gut capacity and feed passage time in broiler chickens. Br. Poult. Sci., 42: 354-361.
CrossRefDirect Link - Svihus, B. and H. Hetland, 2001. Ileal starch digestibility in growing broiler chickens fed on a wheat-based diet is improved by mash feeding, dilution with cellulose or whole wheat inclusion. Br. Poult. Sci., 42: 633-637.
CrossRefDirect Link - Hartini, S., M. Choct, G. Hinch and J.V. Nolan, 2003. The relationship between physico-chemical properties of fibre and their effects on the gut weight of chickens. Proc. Aust. Poult. Sci. Sym., 15: 135-138.
Direct Link - Meng, X., B.A. Slominski, C.M. Nyachoti, L.D. Campbell and W. Guenter, 2005. Degradation of cell wall polysaccharides by combinations of carbohydrase enzymes and their effect on nutrient utilization and broiler chicken performance. Poult. Sci., 84: 37-47.
CrossRefPubMedDirect Link - Biehl, R.R. and D.H. Baker, 1997. Microbial phytase improves amino acid utilization in young chicks fed diets based on soybean meal but not diets based on peanut meal. Poult. Sci., 76: 355-360.
CrossRefPubMedDirect Link - Delezie, E., K. Bierman, L. Nollet and I. Maertens, 2015. Impacts of calcium and phosphorus concentration, their ratio and phytase supplementation level on growth performance, foot pad lesions and hock burn of broiler chickens. J. Applied Poult. Res., 24: 115-126.
CrossRefDirect Link - Bedford, M.R., C.L. Walk and H.V.M. O'Neill, 2016. Assessing measurements in feed enzyme research: Phytase evaluations in broilers. J. Applied Poult. Res., 25: 305-314.
CrossRefDirect Link - Tamim, N.M., R. Angel and M. Christman, 2004. Influence of dietary calcium and phytase on phytate phosphorus hydrolysis in broiler chickens. Poult. Sci., 83: 1358-1367.
PubMedDirect Link - Savory, C.J., 1984. Regulation of food intake by Brown Leghorn cockerels in response to dietary dilution with kaolin. Br. Poult. Sci., 25: 253-258.
CrossRefPubMedDirect Link - Newcombe, M. and J.D. Summers, 1985. Effect of increasing cellulose in diets fed as crumbles or mash on the food intake and weight gains of broiler and leghorn chicks. Br. Poult. Sci., 26: 35-42.
CrossRefDirect Link - Rolls, B.A., A. Turvey and M.E. Coates, 1978. The influence of the gut microflora and of dietary fibre on epithelial cell migration in the chick intestine. Br. J. Nutr., 39: 91-98.
PubMedDirect Link - Bao, Y.M., L.F. Romero and A.J. Cowieson, 2013. Functional patterns of exogenous enzymes in different feed ingredients. World's Poult. Sci. J., 69: 759-774.
CrossRefDirect Link - Lynch, M.B., T. Sweeney, J.J. Callan and J.V. O'Doherty, 2007. Effects of increasing the intake of dietary β-glucans by exchanging wheat for barley on nutrient digestibility, nitrogen excretion, intestinal microflora, volatile fatty acid concentration and manure ammonia emissions in finishing pigs. Animal, 1: 812-819.
CrossRefPubMedDirect Link - Campbell, G.L. and M.R. Bedford, 1992. Enzyme applications for monogastric feeds: A review. Can. J. Anim. Sci., 72: 449-466.
CrossRefDirect Link - Choct, M., R.J. Hughes, J. Wang, M.R. Bedford, A.J. Morgan and G. Annison, 1996. Increased small intestinal fermentation is partly responsible for the anti-nutritive activity of non-starch polysaccharides in chickens. Br. Poult. Sci., 37: 609-621.
CrossRefPubMedDirect Link - Brenes, A., M. Smith, W. Guener and R. Marquardt, 1993. Effect of enzyme supplementation on the performance and digestive tract size of broiler chickens fed wheat- and barley-based diets. Poult. Sci., 72: 1731-1739.
CrossRefPubMedDirect Link - Jozefiak, D., A. Rutkowski and S.A. Martin, 2004. Carbohydrate fermentation in the avian ceca: A review. Anim. Feed Sci. Technol., 113: 1-15.
CrossRefDirect Link - Raninen, K., J. Lappi, H. Mykkanen and K. Poutanen, 2011. Dietary fiber type reflects physiological functionality: comparison of grain fiber, inulin and polydextrose. Nutr. Rev., 69: 9-21.
CrossRefDirect Link - He, L.W., Q.X. Meng, D.Y. Li, Y.W. Zhang and L.P. Ren, 2015. Influence of feeding alternative fiber sources on the gastrointestinal fermentation, digestive enzyme activities and mucosa morphology of growing Greylag geese. Poult. Sci., 94: 2464-2471.
CrossRefPubMedDirect Link - Meynell, G.G., 1963. Antibacterial mechanisms of the mouse gut: II. The role of Eh and volatile fatty acids in the normal gut. Br. J. Exp. Pathol., 44: 209-219.
Direct Link - Jamroz, D., A. Wiliczkiewicz and J. Skorupinska, 1992. The effect of diets containing different levels of structural substances on morphological changes in the intestinal walls and the digestibility of the crude fibre fractions in geese (Part III). J. Anim. Feed Sci., 1: 37-50.
Direct Link - Montagne, L., J.R. Pluske and D.J. Hampson, 2003. A review of interactions between dietary fibre and the intestinal mucosa and their consequences on digestive health in young non-ruminant animals. Anim. Feed. Sci. Technol., 108: 95-117.
CrossRefDirect Link