
Sittisak Khampa
Faculty of Agricultural Technology, Rajabhat Mahasarakham University, P.O. Box 44000, Thailand
Pala Chaowarat
Faculty of Agricultural Technology, Rajabhat Mahasarakham University, P.O. Box 44000, Thailand
Rungson Singhalert
Faculty of Humanities and Social Sciences, Rajabhat Mahasarakham University, P.O. Box 44000, Thailand
Metha Wanapat
Tropical Feed Resources Research and Development Center (TROFREC), Department of Animal Science, Faculty of Agriculture, Khon Kaen University, P.O. Box 40002, Thailand
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 5 | Page No.: 592-596







ABSTRACT
Four, one-year old of dairy steers were randomly assigned according to a 2x2 Factorial arrangement in a 4x4 Latin square design to study supplementation of malate level at 500 vs 1,000 g with yeast (Saccharomyces cerevisiae) at 1,000 vs 2,000 g in concentrate containing high levels of cassava chip. The treatments were as follows: T1 = supplementation of malate at 500 g with yeast at 1,000 g; T2 = supplementation of malate at 500 g with yeast at 2,000 g; T3 = supplementation of malate at 1,000 g with yeast at 1,000 g; T4 = supplementation of malate at 1,000 g with yeast at 2,000 g in concentrate, respectively. The animals were offered the treatment concentrate at 1% BW and ruzi grass was fed ad libitum. The results have revealed that rumen fermentation and blood metabolites were similar for all treatments. The populations of protozoa and fungal zoospores were significantly different as affected by malate level and yeast. In conclusion, the combined use of concentrate containing high level of cassava chip at 70%DM with malate at 1,000 g and yeast at 2,000 g in concentrate with ruzi grass as a roughage could improved rumen ecology in dairy steers.
PDF Abstract XML References Citation
How to cite this article
Sittisak Khampa, Pala Chaowarat, Rungson Singhalert and Metha Wanapat, 2009. Supplementation of Malate and Yeast in Concentrate Containing High Cassava Chip on Rumen Ecology in Dairy Steers. Pakistan Journal of Nutrition, 8: 592-596.
DOI: 10.3923/pjn.2009.592.596
URL: https://scialert.net/abstract/?doi=pjn.2009.592.596
DOI: 10.3923/pjn.2009.592.596
URL: https://scialert.net/abstract/?doi=pjn.2009.592.596
INTRODUCTION
Cassava (Manihot esculenta, Crantz) production in tropical areas has a potential use in ruminant livestock nutrition and feeding. Cassava root contains high levels of energy and has been used as a source of readily fermentable energy in ruminant rations (Wanapat, 2003; Kiyothong and Wanapat, 2004; Promkot and Wanapat, 2005). One strategy for using high degradable carbohydrates is to use in combination with readily available NPN sources such as urea. Urea is commonly used as N source when highly soluble carbohydrates are fed and maintained (Wohlt et al., 1978). However, efficient utilization of protein and Non-protein Nitrogen (NPN) in ruminants depends upon knowledge of the basic principles underlying ruminal microbial N metabolism (Fernandez et al., 1997). Moreover, ruminal pH has great impact on rumen fermentation efficiency (Wanapat, 2003).
Some strictly anaerobic bacteria use a reductive or reverse citric acid cycle known as the succinate-propionate pathway to synthesize succinate and (or) propionate. Both malate and fumalate are key intermediates in the succinate propionate pathway and S. ruminantium uses this pathway (Gottschalk, 1986). The fact dicarboxylic acids, especially malate and fumalate, stimulate lactate utilization is consistent with the presence of this pathway in this ruminal anaerobe (Callaway and Martin, 1996). Previous studies by Sanson and Stallcup (1984) reported that supplementation of malate in ruminant diets has been shown to increase nitrogen retention in sheep and steers and to improve average daily gain and feed efficiency in bull calves. In addition, supplementing diets with yeast (Saccharomyces cerevisiae) increases milk production of dairy cows and weight gain of growing cattle (Brossard et al., 2006). Production responses attributed to yeast are usually related to stimulation of cellulolytic and lactate-utilizing bacteria in the rumen, increased fiber digestion and increased flow of microbial protein from the rumen which may be beneficial for feedlot cattle fed high-grain diets (Guedes et al., 2007). However, the use of malate and yeast in cassava based-diets has not yet been investigated. Therefore, the objective of this experiment was to investigate the supplementation of malate level and yeast supplementation in concentrates containing high level of cassava chip with ruzi grass as a basal roughage on ruminal fermentation and digestibility of nutrients in dairy steers.
MATERIALS AND METHODS
Animals, diets and experimental design: Four, one-year old of dairy steers weighing at 150±10 kg. Cows were randomly assigned according to a 2x2 Factorial arrangement in a 4x4 Latin square design to study two levels malate at 500 vs 1,000 g with yeast (Saccharomyces cerevisiae) at 1,000 vs 2,000 g in concentrates supplementation on ruminal fermentation efficiency and digestibility of nutrients. The dietary treatments were as follows: T1 = supplementation of malate at 500 g with yeast at 1,000 g; T2 = supplementation of malate at 500 g with yeast at 2,000 g; T3 = supplementation of malate at 1,000 g with yeast at 1,000 g; T4 = supplementation of malate at 1,000 g with yeast at 2,000 g in concentrate, respectively. The composition of dietary treatments and ruzi grass used are shown in Table 1, 2.
Cows were housed in individual pens and individually fed concentrate at 1%BW. All cows were fed ad libitum of ruzi grass with water and a mineral-salt block. Feed intake of concentrate and roughage were measured separately and refusals recorded. The experiment was run in four periods, each experimental period lasted for 21 days, the first 14 days for treatment adaptation and for feed intake measurements whist the last 7 days were for sample collections of rumen fluid and faeces. Body weights were measured daily during the sampling period prior to feeding.
Data collection and sampling procedures: Roughage and concentrate were sampled daily during the collection period and were composted by period prior to analyses. Composites samples were dried at 60oC and ground (1 mm screen using Cyclotech Mill, Tecator, Sweden) and then analyzed for DM, ether extract, ash and CP content (AOAC, 1985), NDF, ADF and ADL (Goering and Van Soest, 1970).
Rumen fluid samples were collected at 0, 2 and 4 h post-feeding. Approximately 200 ml of rumen fluid was taken from the middle part of the rumen by a stomach tube connected with a vacuum pump at each time at the end of each period. Rumen fluid was immediately measured for pH and temperature using (HANNA instruments HI 8424 microcomputer) after withdrawal. Rumen fluid samples were then filtered through four layers of cheesecloth. Samples were divided into two portions. One portion was used for NH3-N analyses where 5 ml of H2SO4 solution (1 M) was added to 50 ml of rumen fluid. The mixture was centrifuged at 16,000 g for 15 min and the supernatant stored at -20oC prior to NH3-N analysis using the micro Kjeldahl methods (AOAC, 1985). Another portion was fixed with 10% formalin solution in normal saline (Galyean, 1989).
The total count of bacteria, protozoa and fungal zoospores were made using the methods of Galyean (1989) based on the use of a haematocytometer (Boeco). A blood sample (about 10 ml) was drawn from the jugular vein at the same time as rumen fluid sampling, separated by centrifugation at 5,000 g for 10 minutes and stored at -20oC until analysis of Blood Urea Nitrogen (BUN) according to the method of Crocker (1967).
Statistical analysis: All data obtained from the experiment were subjected to ANOVA for a 4x4 Latin square design with 2x2 Factorial arrangement of treatments using the General Linear Models (GLM) procedures of the Statistical Analysis System Institute (SAS, 1998). Treatment means were compared by Duncan’s New Multiple Range Test (DMRT) (Steel and Torrie, 1980).
RESULTS AND DISCUSSION
Chemical composition of feeds and feed intake: The chemical compositions of roughage and concentrate diets fed in dairy cows are presented in Table 2. Concentrate diets contained similar concentrations of DM, OM, CP, NDF, ADF and TDN. Diets containing high levels of cassava chip based diets had a slightly higher Non-structural Carbohydrate (NSC) and lower NDF due to increased level of cassava chip in the diets. Furthermore, the chemical composition of ruzi grass is presented in Table 2.
The effects of malate level with yeast (Saccharomyces cerevisiae) on feed-intake of dairy steers are presented in Table 2. Feed intake were non-significantly different among treatments and was higher in dairy steers receiving T4 than T3, T2, T1 (2.7, 2.6, 2.6 and 2.5% BW, respectively). This data indicated that malate level with yeast supplementation had no effect on feed-intake in dairy steers. This result was in agreement with earlier work by (Sommart et al., 2000 and Khampa et al., 2006) which reported that inclusion of cassava chip in diets resulted in satisfactory animal performance and had no negative effects on animal health in finishing beef cattle and lactating dairy cows.
| Table 1: | Ingredients of concentrate used in the experiment (% DM basis) |
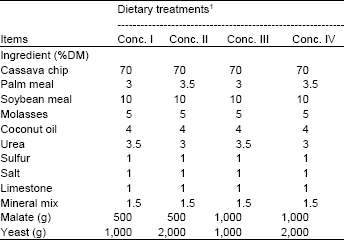 | |
| 1Conc. = concentrate; | |
| Table 2: | Chemical composition of concentrates and ruzi grass used in the experiment |
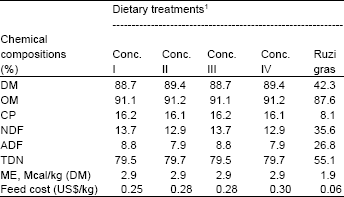 | |
| DM = dry matter, CP = crude protein, OM = organic matter, NDF = neutral detergent fiber, ADF = acid detergent fiber, TDN = total digestible of nutrients, ME = metabolizable energy. Conc. = concentrate | |
| Table 3: | Effects of malate level and yeast (Saccharomyces cerevisiae) on feed-intake and rumen ecology in dairy steers |
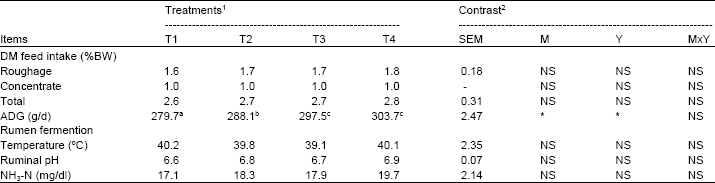 | |
| a,b,cValues on the same row with different superscripts differ (p<0.05). 1T1 = malate at 500 g with yeast at 1,000 g. T2 = malate at 500 g with yeast at 2,000 g. T3 = malate at 1,000 g with yeast at 1,000 g. T4 = malate at 1,000 g with yeast at 2,000 g. 2 Probability of main effects of level malate (M) in concentrates (500 vs 1,000 g), levels of yeast (Y) (1,000 vs 2,000 g), or the MxY interaction. * = p<0.05, NS = p>0.05. | |
| Table 4: | Effects of malate level and yeast on ruminal microorganisms in dairy steers |
 | |
| a,b,cValues on the same row with different superscripts differ (p<0.05). 1T1 = malate at 500 g with yeast at 1,000 g. T2 = malate at 500 g with yeast at 2,000 g. T3 = malate at 1,000 g with yeast at 1,000 g. T4 = malate at 1,000 g with yeast at 2,000 g. 2Probability of main effects of level malate (M) in concentrates (500 vs 1,000 g), levels of yeast (Y) (1,000 vs 2,000 g), or the MxY interaction. * = p<0.05, NS = p>0.05. | |
Characteristics of ruminal fermentation and blood metabolism: Rumen ecology parameters were measured for temperature, pH and NH3-N (Table 3). Rumen pH at 0, 2 and 4 h post-feeding were unchanged by dietary treatments and the values were quite stable at 6.6-6.9, but all treatment means were within the normal range which has been reported as optimal for microbial digestion of fiber and also digestion of protein (6.0-7.0) (Hoover, 1986).
Ruminal NH3-N concentrations was not altered by malate level and yeast supplement in diets containing high cassava-based diets. As NH3-N is regarded as the most important nitrogen source for microbial protein synthesis in the rumen. In addition, the result obtained was closer to optimal ruminal NH3-N between at 15-30 mg% (Wanapat and Pimpa, 1999; Chanjula et al., 2003, 2004) for increasing microbial protein synthesis, feed digestibility and voluntary feed intake in ruminant fed on low-quality roughages.
Rumen microorganisms populations: Table 4 presents rumen microorganism populations. The populations of fungal zoospores, protozoa and total bacteria direct counts were significantly different and populations of bacteria had higher numbers in heifer receiving diets T4 than T3, T2 and T1. In contrast, the present number of protozoa in the rumen was decreased by malate level and yeast supplementation in high cassava-based diets. In the experiment by Newbold et al. (1996) has shown that feeding 100 mg of malate per day in sheep caused an increase in the number of total bacteria and tended to increase the population of cellulolytic bacteria. In agreement with these observations, Lopez et al. (1999) reported that fumalate (another intermediate in the succinate to propionate pathway) increased the number of cellulolytic bacteria almost three-fold during fermentation in the RUSITEC system. In addition Guedes et al. (2007) reported that yeast are usually related to stimulation of cellulolytic and lactate-utilizing bacteria in the rumen, increased fiber digestion and increased flow of microbial protein from the rumen which may be beneficial for feedlot cattle fed high-grain diets. As cassava chip can be readily degraded in the rumen and ruminal pH was decreased, malate could stimulate lactate utilization by S. ruminantium and could improve pH in the rumen. It is possible that supplementation of malate with yeast may play an important role in increasing bacterial populations. Moreover, Martin et al. (1999) reported that increasing dietary concentrations of malate might help to reduce problems associated with ruminal acidosis by stimulating lactate utilization by S. ruminantium.
Conclusions: Based on this experiment, it could be concluded that supplementation of malate level with yeast (Saccharomyces cerevisiae) in concentrate containing high level of cassava chip maintained could improved ruminal fermentation efficiency. Moreover, high level of cassava chip in diet resulted increase populations of bacteria, but decreased protozoal populations. These results suggest that the combined use of concentrates containing high level of cassava chip at 70% DM with malate at 1,000 g and yeast at 2,000 g in concentrate could highest improved rumen ecology in dairy steers.
ACKNOWLEDGMENTS
The authors would like to express their most sincere gratitude and appreciation to the Rajabhat Mahasarakham University, Tropical Feed Resources Research and Development Center (TROFREC) and The National Research Council of Thailand for their financial support of research and the use of research facilities.
REFERENCES
- Brossard, L., F. Chaucheyras-Dur, B. Michalet-Doreau and C. Martin, 2006. Dose effect of live yeasts on rumen microbial communities and fermentations during butyric latent acidosis in sheep new type of interaction. Anim. Sci., 82: 829-836.
CrossRef - Callaway, T.R. and S.A. Martin, 1996. Effects of organic acid and monensin treatment on in vitro mixed ruminal microorganism fermentation of cracked corn. J. Anim. Sci., 74: 1982-1989.
PubMed - Chanjula, P., M. Wanapat, C. Wachirapakorn, S. Uriyapongson and P. Rowlinson, 2003. Ruminal degradability of tropical feeds and their potential use in ruminant diets. Asian-Aust. J. Anim. Sci., 16: 211-216.
Direct Link - Chanjula, P., M. Wanapat, C. Wachirapakorn, S. Uriyapongson and P. Rowlinson, 2004. Effect of synchronizing starch sources and protein (NPN) in the rumen on feed intake, rumen microbial fermentation, nutrient utilization and performance of lactating dairy cows. Asian-Aust. J. Anim. Sci., 17: 1400-1410.
Direct Link - Crocker, C.L., 1967. Rapid determination of urea nitrogen in serum or plasma without deproteinization. Am. J. Med. Technol., 33: 361-365.
PubMedDirect Link - Fernandez, J.M., T. Sahulu, C. Lu, D. Ivey and M.J. Potchoiba, 1997. Production and metabolic aspects of non-protein nitrogen incorporation in lactation rations of dairy goats. Small Ruminant Res., 26: 105-107.
CrossRef - Guedes, C.M., D. Goncalves, M.A.M. Rodrigues and A. Dias-da-Silva, 2007. Effects of a Saccharomyces cerevisiae yeast on ruminal fermentation and fibre degradation of maize silages in cows. Anim. Feed Sci. Technol., 145: 27-40.
CrossRefDirect Link - Hoover, W.H., 1986. Chemical factors involved in ruminal fiber digestion. J. Dairy Sci., 69: 2755-2766.
CrossRefPubMedDirect Link - Khampa, S., M. Wanapat, C. Wachirapakorn, N. Nontaso, M.A. Wattiaux and P. Rowlison, 2006. Effect of levels of sodium DL-malate supplementation on ruminal fermentation efficiency of concentrates containing high levels of cassava chip in dairy steers. Aisan-Aust. J. Anim. Sci., 19: 368-375.
CrossRefDirect Link - Lopez, S., C. Valdes, C.J. Newbold and R.J. Wallace, 1999. Influence of sodium fumarate addition on rumen fermentation in vitro. Br. J. Nutr., 81: 59-64.
PubMed - Martin, S.A., M.N. Streeter, D.J. Nisbet, G.M. Hill and S.E. Williams, 1999. Effect of DL-malate on ruminal metabolism and performance of cattle fed a high concentrate diets. J. Anim. Sci., 77: 1008-1015.
Direct Link - Newbold, C.J., R.J. Wallace and F.M. McIntosh, 1996. Mode of action of the yeast Saccharomyces cerevisiae as a feed additive for ruminants. Br. J. Nutr., 76: 249-261.
CrossRefPubMedDirect Link - Promkot, C. and M. Wanapat, 2005. Effect of level of crude protein and use of cottonseed meal in diets containg cassava chips and rice straw for lactating dairy cows. Asian-Aust. J. Anim. Sci., 18: 502-511.
Direct Link - Sanson, D.W. and O.T. Stallcup, 1984. Growth response and serum constituents of Holstein bulls fed malic acid. Nutr. Rep. Int., 30: 1261-1267.
Direct Link - Sommart, K., M. Wanapat, D.S. Parker and P. Rowlinson, 2000. Cassava chip as an energy source for lactating dairy cows fed rice straw. Asian Aust. J. Anim. Sci., 13: 1094-1101.
Direct Link - Wanapat, M., 2003. Manipulation of cassava cultivation and utilization to improve protein to energy biomass for livestock feeding in the tropics. Asian-Aust. J. Anim. Sci., 16: 463-472.
CrossRefDirect Link - Wanapat, M. and O. Pimpa, 1999. Effect of ruminal NH3-N levels on ruminal fermentation, purine derivatives, digestibility and rice straw intake in swamp buffaloes. Asian-Aust. J. Anim. Sci., 12: 904-907.
CrossRefDirect Link