
D.J. Mukesh Kumar
Centre for Advanced Studies in Botany, University of Madras, Guindy Campus, Chennai, Tamil Nadu, India
R. Rakshitha
Department of Biotechnology, Kalasalingam University, Srivilliputtur, Tamil Nadu, India
M. Annu Vidhya
Department of Biotechnology, Kalasalingam University, Srivilliputtur, Tamil Nadu, India
P. Sharon Jennifer
Department of Biotechnology, Kalasalingam University, Srivilliputtur, Tamil Nadu, India
Sandip Prasad
Department of Biotechnology, Kalasalingam University, Srivilliputtur, Tamil Nadu, India
M. Ravi Kumar
Centre for Advanced Studies in Botany, University of Madras, Guindy Campus, Chennai, Tamil Nadu, India
P.T. Kalaichelvan
Centre for Advanced Studies in Botany, University of Madras, Guindy Campus, Chennai, Tamil Nadu, India
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 4 | Page No.: 529-534







ABSTRACT
The present study aimed at the production, purification and characterization of fibrinolytic nattokinase enzyme from the bacteria isolated from natto food. For the purpose, a fibrinolytic bacterium was isolated and identified as Bacillus subtilis based on 16S rDNA sequence analysis. The strain was employed for the production and optimization of fibrinolytic enzyme. The strain showed better enzyme production during 72nd h of incubation time with 50°C at the pH 9. The lactose and peptone were found to be increasing the enzyme production rate. The enzyme produced was purified and also characterized with the help of SDS-PAGE analysis. The activity and stability profile of the purified enzyme was tested against different temperature and pH. The observations suggesting that the potential of fibrinolytic enzyme produced by Bacillus subtilis RJAS19 for its applications in preventive medicines.
PDF Abstract XML References Citation
Received: March 19, 2013;
Accepted: April 04, 2013;
Published: November 26, 2013
How to cite this article
D.J. Mukesh Kumar, R. Rakshitha, M. Annu Vidhya, P. Sharon Jennifer, Sandip Prasad, M. Ravi Kumar and P.T. Kalaichelvan, 2014. Production, Optimization and Characterization of Fibrinolytic Enzyme by Bacillus subtilis RJAS19. Pakistan Journal of Biological Sciences, 17: 529-534.
DOI: 10.3923/pjbs.2014.529.534
URL: https://scialert.net/abstract/?doi=pjbs.2014.529.534
DOI: 10.3923/pjbs.2014.529.534
URL: https://scialert.net/abstract/?doi=pjbs.2014.529.534
INTRODUCTION
Nattokinase is considered as an active fibrinolytic enzyme synthesized by bacteria belongs to the Bacillus sp. and was identified in natto, a Japanese traditional food (Sumi et al., 1987). Natto is rich in protein and was made by fermenting the soyabeans with Bacillus subtilis. Nattokinase is a serine protease, belongs to the subtilisin family which exhibits increased fibrinolytic activity when compared to plasmin (Fujita et al., 1995; Peng et al., 2003). The increased fibrinolytic activity of nattokinase makes it feasible for the thrombolytic therapy.
The nattokinase possess worldwide attention due to its various therapeutic applications. The breakdown of fibrin by nattokinase can be carried out by promoting the activity of urokinase and tissue plasminogen activator, which in turn promotes the conversion of plasminogen into plasmin and degrades fibrin (Milner and Makise, 2002). Fibrin is a soluble plasma glycoprotein made by fibrinogen in liver. Nattokinase possess various applications including anti-hypertension, blood thinning, thrombolytic and digestive capability (Yokota et al., 1996). They were also used for cardiovascular diseases.
The nattokinase is a food based dietary supplement being marketed worldwide and found to be stable in the gastrointestinal tract, despite of changes in pH and temperature. Nattokinase is produced by bacteria of genus Bacillus especially Bacillus subtilis which is Generally Recognized As Safe (GRAS) organism. During the past decades, considerable progresses have been made to engineer B. subtilis for the production of nattokinase (Milner and Makise, 2002).
With the help of the above points, the present study was aimed at the isolation of a fibrinolytic nattokinase producing bacteria from natto. The production of fibrinolytic enzyme was carried out followed by its optimization. The enzyme produced was then characterized further.
MATERIALS AND METHODS
Chemicals and sample: All the chemicals used in the present study were of analytical grade and bought from Sigma Alrich and SRL.
Isolation of fibrinolytic bacteria: The Bacillus subtilis RJAS19 (Genbank Accession No. KC527834), producing fibrinolytic enzyme was isolated from fermented soya food. The strain was characterized biochemically and identified based on 16S rDNA sequence analysis and was stored at -4°C for further analytical procedures.
Production of fibrinolytic enzyme: For the production of fibrinolytic enzyme, the bacterial inoculum was prepared with the help of LB broth. The overnight grown bacteria culture was inoculated on 100 mL of production medium (Galactose 1 g, NaCl 0.5 g, CaCO3 0.5 g, Tryptone 0.5 g, Soybean 0.3 g, MgSO4.7H2O, KH2PO4 0.08 g, K2HPO4 0.02 g, CuSO4 0.005 g, FeSO4 0.001 g, Distilled water 100 mL) (Hassanein et al., 2011). The production of enzyme was facilitated by incubating the production medium in shaken condition for 48 h at 30°C. The production medium was centrifuged and the supernatant was used as enzyme source.
Assay procedures
| • | Biomass: The biomass was determined by reading the absorbance of production medium at 600 nm (Henriette et al., 1993) |
| • | Total protein content: The amount of protein present in the culture filtrate was measured by Bradford’s method (Bradford, 1976) |
| • | Fibrinolytic enzyme: The fibrinolytic activity was assayed with the help of fibrin as a substrate (Hassanein et al., 2011). The reaction mixture consists of 2 mL of fibrin, 7 mL of sodium phosphate buffer and 1 mL of culture filtrate was incubated at 37°C for 5 min. the reaction was stopped by adding 1 mL of 5% Trichloroacetic acid. The reaction mixture was incubated for 30 mins at 4°C and centrifuged to collect the supernatant. The absorbance was read at 750 nm. One unit of fibrinolytic activity was defined as the amount of fibrin that hydrolyzed by the enzyme during standard incubation conditions |
Parameter optimization studies: For the production of fibrinolytic enzyme the cultural conditions like incubation time, pH and temperature were optimized. The parameters optimization was done by cultivating the bacteria at different incubation time (24-144 h), different pH (5-11) and different temperature (35-65°C). Apart from the environmental factors, medium components such as carbon (Sucrose, Lactose, Glucose, Starch, Fructose, Mannose) and nitrogen (Yeast extract, Beef extract, Peptone, Ammonium chloride, Sodium nitrate, Ammonium carbonate) sources were also optimized.
Extraction and purification of fibrinolytic enzyme: The enzyme was allowed to produce under optimized production conditions. The enzyme from the culture filtrate was extracted with the help of ammonium sulphate precipitation with 40-60% saturation. The extracted protein was dialyzed overnight with sodium phosphate buffer with three buffer transfers. This partially purified fibrinolytic enzyme was then purified with the help of ion exchange chromatography with CM-Sepharose column (Yin et al., 2010).
Characterization of purified fibrinolytic enzyme:
| • | SDS-PAGE analysis: The molecular weight (kDa) of the purified fibrinolytic enzyme was analyzed using SDS-PAGE analysis (LaemmLi, 1970) |
| • | Effect of temperature and pH on enzyme stability: The effect of temperature and pH on the stability of purified fibrinolytic activity was determined with the help of fibrin as the substrate. The pH stability of the purified enzyme was determined by incubating the purified enzyme in buffers with different pH from 8.0 to 12.0 at 50°C for 40 mins (pH 5.0-7.0 Sodium phosphate buffer, pH 7.0-9.0 Tris Hcl buffer, pH 9.0-12.0 Sodium carbonate buffer). Followed by the incubation the relative activity of the enzyme was determined according to Chang et al. (2000). Similarly, the temperature stability of the purified enzyme was determined by incubating the enzyme at different temperatures ranging from 45-95°C at the pH 9.0 |
| • | Effect of temperature and pH on enzyme activity: The pH dependence of the enzyme was determined by using fibrin as substrate in a series of buffers of different pH values ranging from 8.0-2.0 (buffers mentioned above). The fibrinolytic activity of the purified enzyme was measured at different pH after 30 min incubation. The effect of temperature was determined by incubating the samples at different temperatures ranging from 45-95°C for 30 min and the activity of the enzyme was measured by standard conditions |
Application of purified fibrinolytic enzyme: The application of fibrinolytic enzyme was evaluated with the help of the method followed by Omura et al. (2005) by degrading the artificial blood clot. For the same, fresh blood collected from the healthy volunteer was allowed to coagulate in glass test tube. The artificial blood clot was rinsed with phosphate buffer in prior to the treatment with purified enzyme and incubated for 60 min. For control, the blood clot was treated with saline. The observation was made for the degradation of blood clot.
RESULTS AND DISCUSSION
In current scenario, enzymes play as active ingredients in various applications in the fields of food, detergent, medicine, etc. Since enzymes overcomes difficulties of the conventional methods in the breakdown of complex matters (Negi and Banerjee, 2010). Nattokinase is one of the fibrinolytic enzymes which possess applications in medical field. The fibrinolytic activity of the nattokinase made it one of the life saving drug. Since the enzyme having vast applications in medical, the present study was aimed at the isolation, production and purification of fibrinolytic enzyme.
The Bacillus subtilis strain RJAS19 was selected as potent fibrinolytic organism and was used for the present study.
For the production of enzyme, the culture medium plays an important role, as the culture medium should provide all the essential nutrients required by the organism for enzyme production (Al-Juamily and Al-Zaidy, 2012). In order to minimize the production cost, production medium must possess cheap source of carbon, nitrogen, growth promoters and trace elements. The growth conditions such as pH, incubation time and temperature were also maintained at optimal conditions for the enhanced enzyme production. In prior to the inoculation, the cultural and medium conditions were to be optimized.
At very first, the incubation time was optimized by harvesting the production medium at different intervals and assayed for enzyme production. The production of the enzyme by the strain was observed in exponential phase and showed linearity with the cell growth. The maximum production of nattokinase was observed during 72nd h of incubation (Fig. 1). The production of fibrinolytic enzymes usually lies between 48 to 96 h (Krishnaveni and Rani, 2011). The extra cellular enzymes are usually produced during log phase but within the log phase the fermentation time may be vary.
The incubation temperature is the next most critical factor that has to be optimized for enzyme production. The optimization of temperature is very important as it determines the velocity of the enzyme reaction. All enzymes have an optimal temperature at which reaction rates go fastest without denaturing the enzyme (Al-Juamily and Al-Zaidy, 2012). The optimum temperature for the fibrinolytic enzyme production by Bacillus subtilis was found to be 50°C and beyond which the enzyme activity was reduced gradually (Fig. 2). From the observed results, the isolated strain was found to be thermophillic.
When come to the pH, considerably less production of enzyme was observed at acidic pH and increased enzyme production was obtained at weak alkaline pH. The maximum production of fibrinolytic enzyme was noticed at pH of 9.0 (Fig. 3). The results indicated that the isolated Bacillus subtilis was an alkaline organism. Changes in pH can change the shape of the active site and during high or low pH concentrations result in loss of enzyme activity due to denaturation.
The medium components play a significant role in enhancing the enzyme production (Gupta et al., 2002). Various carbon sources were used for the production of fibrinolytic enzyme by the isolated Bacillus subtilis. Lactose was found to be the best carbon source for fibrinolytic enzyme production (Fig. 4).
Bacillus subtilis RJAS19 was allowed to grow in the presence of different nitrogen sources. Interestingly increased fibrinolytic activity was achieved, when the cells were grown in a medium containing peptone (Fig. 5).
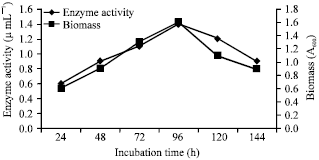 | |
| Fig. 1: | Enzyme activity and growth profile with respect to different incubation time |
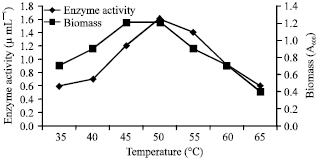 | |
| Fig. 2: | Enzyme activity and growth profile with respect to different temperature |
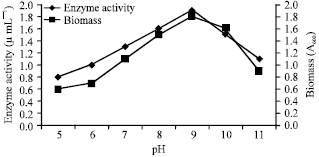 | |
| Fig. 3: | Enzyme activity and growth profile with respect to different pH |
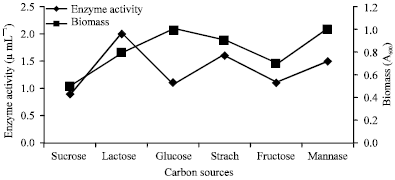 | |
| Fig. 4: | Enzyme activity and growth profile with respect to different carbon sources |
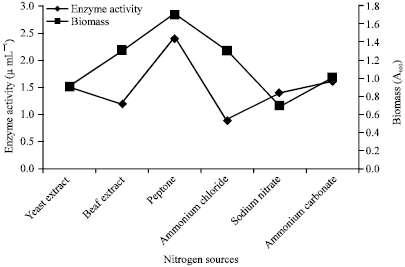 | |
| Fig. 5: | Enzyme activity and growth profile with respect to different nitrogen sources |
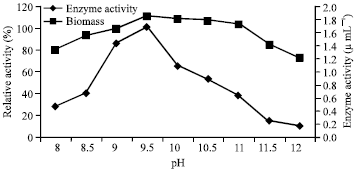 | |
| Fig. 6: | Effect of pH on enzyme activity and stability |
In contrast, a medium containing casein and soyabean meal enhanced the production of similar proteolytic enzyme by bacteria (Kiran et al., 2008; Saurabh et al., 2007).
The enzyme produced under optimal conditions was precipitated by ammonium sulphate precipitation. The further purification was done using dialysis. The dialysed enzyme samples were subjected to SDS-PAGE analysis.
The molecular weight of the enzyme produced was analyzed by SDS-PAGE. The purified fibrinolytic enzyme was checked for its homogeneity and molecular weight was determined by SDS-PAGE. The purified fibrinolytic enzyme migrated as a single band corresponding to molecular mass of 35 kDa. The findings are in accordance with Chang et al. (2000) and Borah et al. (2012).
While carrying out stability test, the purified fibrinolytic enzyme was found to be stable at pH 9.5 to 11.0 (Fig. 6). The pH stability of the fibrinolytic enzyme was found to be similar to that of similar enzyme produced by B. amyloliquefaciens DC-4 (Peng et al., 2003).
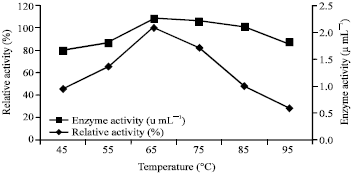 | |
| Fig. 7: | Effect of temperature on enzyme activity and stability |
The thermostability of the enzyme was found to be high at 65 to 85°C (Fig. 7). The similar thermal stability profile was also observed in the strains such as Bacillus subtilis YJ1 (Yin et al., 2010), B. amyloliquefaciens DC-4 (Peng et al., 2003) and Bacillus sp. KCK-7 (Kim et al., 1996).
In further to the stability test, the enzyme activity profile was tested against the broad range of temperature and pH. From the observations, the enzyme activity was found to be maximum at 65°C and at the pH of 9.5. The similar enzyme produced by Bacillus subtilis YJ1 (Yin et al., 2010) and Bacillus subtilis (Dubey et al., 2011) were also possess similar optimal conditions. The ability of fibrinolytic enzyme to degrade the blood clot was experimented as the method followed by Omura et al. (2005). While comparing to the control, the enzyme degraded the clot completely which indicated the degrading ability of the enzyme. The theory was previously supported by Sumi et al. (1987).
CONCLUSION
From the results obtained in the present investigation, the isolated bacterial strain Bacillus subtilis RJAS19 was found to be having ability to synthesize economically important fibrinolytic enzyme. Furthermore, the activity and stability profile of the fibrinolytic enzyme from Bacillus subtilis RJAS19 suggesting its potential for the further applications in preventive medicines.
REFERENCES
- Al-Juamily, E.F. and B.H. Al-Zaidy, 2012. Optimization conditions of production fibrinolytic enzyme from Bacillus lichniformis B4 local isolate. Br. J. Pharmacol. Toxicol., 3: 289-295.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chang, C.T., M.H. Fan, F.C. Kuo and H.Y. Sung, 2000. Potent fibrinolytic enzyme from a mutant of Bacillus subtilis IMR-NK1. J. Agric. Food Chem., 48: 3210-3216.
CrossRef - Borah, D., R. Yadav, A. Sangra, L. Shahin and A.K. Chaubey, 2012. Production, purification and characterization of nattokinase from Bacillus subtilis, isolated from tea garden soil samples of Dibrugarh, Assam. Asian J. Pharm. Clin. Res., 5: 124-125.
Direct Link - Fujita, M., Y. Ito, K. Hong and S. Nishimuro, 1995. Characterization of nattokinase-degraded products from human fibrinogen or cross-linked fibrin. Fibrinolysis, 9: 157-164.
CrossRef - Gupta, R., Q. Beg and P. Lorenz, 2002. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol., 59: 15-32.
CrossRefPubMedDirect Link - Henriette, C., S. Zinebi, M.F. Aumaitre, E. Petitdemange and H. Petitdemange, 1993. Protease and lipase production by a strain of Serratia marcescens. J. Ind. Microbiol., 12: 129-135.
CrossRefDirect Link - Kim, W., K. Choi, Y. Kim, H. Park and J. Choi et al., 1996. Purification and characterization of a fibrinolytic enzyme produced from Bacillus sp. strain CK 11-4 screened from Chungkook-Jang. Applied Environ. Microbiol., 62: 2482-2488.
Direct Link - Kiran, G.S., S. Shanmughapriya, S. Jayalakshmi, J. Selvin and R. Gandhimathi et al., 2008. Optimization of extracellular psychrophilic alkaline lipase produced by marine Pseudomonas sp. (MSI057). Bioprocess Biosyst. Eng., 31: 483-492.
CrossRefPubMedDirect Link - Krishnaveni, A. and T.S. Rani, 2011. Phyto-pharmacological review of Argyreia nervosa. Int. Res. J. Pharm., 2: 28-31.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Yin, L.J., H.H. Lin and S.T. Jiang, 2010. Bioproperties of potent nattokinase from Bacillus subtilis YJ1. J. Agric. Food Chem., 58: 5737-5742.
CrossRef - Milner, M. and K. Makise, 2002. Natto and its active ingredient nattokinase: A potent and safe thrombolytic agent. Altern. Complement. Ther., 8: 157-164.
CrossRef - Peng, Y., Q. Huang, R.H. Zhang and Y.Z. Zhang, 2003. Purification and characterization of a fibrinolytic enzyme produced by Bacillus amyloliquefaciens DC-4 screened from dou-chi, a traditional Chinese soybean food. Comp. Biochem. Physiol., 134: 45-52.
CrossRefDirect Link - Negi, S. and R. Banerjee, 2010. Optimization of culture parameters to enhance production of amylase and protease from Aspergillus awamori in a single fermentation. Afr. J. Biochem. Res., 4: 73-80.
Direct Link - Saurabh, S., I. Jasmine, G. Pritesh and S.R. Kumar, 2007. Enhanced productivity of serine alkaline protease by Bacillus sp. using soybean as substrate. Mala. J. Microbiol., 3: 1-6.
Direct Link - Sumi, H., H. Hamada, H. Tsushima, H. Mihara and H. Muraki, 1987. A novel fibrinolytic enzyme (nattokinase) in the vegetable cheese Natto: A typical and popular soybean food in the Japanese diet. Experientia, 43: 1110-1111.
CrossRef - Yokota, T., T. Hattori, H. Ohishi, K. Hasegawa and K Watanabe, 1996. The effect of antioxidant-containing fraction from fermented soybean food on atherosclerosis development in cholesterol-fed rabbits. LWT-Food Sci. Technol., 29: 751-755.
CrossRef - Hassanein, W.A., E. Kotb, N.M. Awny and Y.A. El-Zawahry, 2011. Fibrinolysis and anticoagulant potential of a metallo protease produced by Bacillus subtilis K42. J. Biosci., 36: 773-779.
CrossRefDirect Link - Omura, K., M. Hitosugi, X. Zhu, M. Ikeda, H. Maeda and S. Tokududome, 2005. A newly derived protein from Bacillus subtilis natto with both antithrombotic and fibrinolytic effects. J. Pharmacol. Sci., 99: 247-251.
PubMedDirect Link