
Fotso
Laboratory of Biochemical and Plant Physiology, Department of Biological Sciences, Higher Teacher Training School, P.O. Box 47, Yaounde, Cameroon
Oumar
Jay P. J. Biotechnology Laboratory, Institute of Agricultural Research for Development (IRAD), P.O. Box 25, Buea, Ekona, Cameroon
Niemenak Nicolas
Laboratory of Biochemical and Plant Physiology, Department of Biological Sciences, Higher Teacher Training School, P.O. Box 47, Yaounde, Cameroon
Donfagsiteli Tchinda Nehemie
Laboratory of Botanic and Traditional Medicine, Institute of Medical Research and Medicinal Plants Studies (IMPM), P.O. Box 6163, Yaounde, Cameroon
Omokolo Ndoumou Denis
Laboratory of Biochemical and Plant Physiology, Department of Biological Sciences, Higher Teacher Training School, P.O. Box 47, Yaounde, Cameroon
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 5 | Page No.: 726-732







ABSTRACT
A productive genotype of Irvingia gabonensis were cultured in vitro for induction embryogenic calli, somatic embryogenesis and regeneration of plantlets. Fragments of young leaves were used as primary explants. Callogenesis was initiated by culture of explants during 30 days on Murashige and Skoog medium half strength (MS/2) supplemented with 1-6 mg L-1 of 2,4-dichlorophenoxyacetic acid (2,4-D). The highest percentage of explants forming calli is 85.1% at 3 mg L-1 of 2,4-D. Somatic embryos were obtained after a sub-culture of embryogenic calli during 60 days on MS/2 supplemented with 1-3 mg L-1 of BAP. The highest percentage of embryogenic calli which differentiates somatic embryos is 63.8 ± 2.3% at 1 mg L-1 of 6-benzylaminopurine (BAP). The highest number of somatic embryos per callus which is 43.6 is obtained with 2 mg L-1 of this phytohormone. When isolated from calli and sub-cultured during 30 days on MS/2 supplemented with 2 mg L-1 of BAP, somatic embryos germinate with a highest percentage of 83%. The sub-culture of germinated somatic embryos on the same Basal Medium (BM) supplemented with 4 mg L-1 of BAP and 2 mg L-1 of Naphthalene Acetic Acid (NAA) during 80 days gives rise to the plantlets with 82.7 ±4.8% of success. With this combination, each plantlet has average length of 5.6 cm, bears 3.3 leaves and 7.2 roots with 1 or 2 pivoting roots. Plantlets acclimatized on a mixture sterilized soil/vermiculite at equal volume survive at 93%. Results of this study constitute a new way for a production of Irvingia gabonensis seedlings with pivoting root and they permit to arrest the difficulties of natural and horticultural reproduction.
PDF Abstract XML References Citation
How to cite this article
Fotso, Oumar, Niemenak Nicolas, Donfagsiteli Tchinda Nehemie and Omokolo Ndoumou Denis, 2008. In vitro Regeneration of Irvingia gabonensis by Somatic
Embryogenesis. Pakistan Journal of Biological Sciences, 11: 726-732.
DOI: 10.3923/pjbs.2008.726.732
URL: https://scialert.net/abstract/?doi=pjbs.2008.726.732
DOI: 10.3923/pjbs.2008.726.732
URL: https://scialert.net/abstract/?doi=pjbs.2008.726.732
INTRODUCTION
Irvingia gabonensis from the family of Irvingiaceae is known as bush mango and as non timber forest products (Vivien and Faure, 1996). It is a fruit tree native to moist lowland tropical forest in central and West Africa. The uses of this tree are varied. The kernel of the nut is a food additive. It is used to thicken and flavour soups. In Gabon, the nut is used in the preparation of Dika bread. Oil can be extracted from the nut. The quality of this oil is comparable to sheaf oil. The mesocarp of fruit is appreciated as a fresh fruit. It can be also used for the preparation of juice, jelly and jam. The sugar concentration of this juice is comparable with pineapples and oranges (Akubor, 1996). This juice contents higher concentration of ascorbic acid (67 mg/100 mL). The wood of the tree is used in carpentry (Ngoye, 1998). Anti diarrhoeic and anti ulcer properties has been reported in Irvingia gabonensis (Raji et al., 2001). Despite the vast array of potentialities, this species is not yet been integrated in the farming system and fruits are still harvested mainly from wild trees. The International Centre for Research in Agro Forestry (ICRAF) identifies Irvingia gabonensis as a priority wild fruit tree species for domestication (Lapido et al., 1996). Irvingia gabonensis is believed to be predominantly out crossing and insect pollinated with seed dispersed by humans during migration and through the digestive system of mammals (Lowe et al., 2000). These indicate that Irvingia gabonensis is highly heterozygote. This results in a great diversity of vegetative and organoleptic characters. Therefore, the problem of multiplication of selected individuals exists. To fight against this problem, vegetative propagation like grafting, cutting and aerial layering are done (Shiembo et al., 1996). But these classic methods are limited by the difficulty of rooting and field growth performance after transplantation as in many woody plants (Hamzah, 1992; Newton et al., 1992). Till date, very little research has been carried on the tissue culture of Irvingia gabonensis compared to the other wild plants such as Bauhina vahlii (Dhar and Upreti, 1999), Tilophora indica (Jayanthi and Mandala, 2001) mango (Litz et al., 1998), Acacia tortilis (Sane et al., 2000) and Plumbago species (Das and Rout, 2002). However, Omokolo et al. (2004) showed that it is possible to propagate this species by in vitro micro cutting. But, plantlets produced by this method have a spread root system, which can not permit them to resist to the hard climatic factors. However, the production of plantlets with pivoting root system through somatic embryogenesis, capable to resist to hard climatic factors has been reported in some species such as Theobroma cacao (Alemano et al., 2001), Hevea brasiliensis (Blanc et al., 2002), Tlophora indica (Jayanthi and Mandala, 2001). But in literature, none research has been reported on somatic embryogenesis of Irvingia gabonensis. However, this method can be used for the rapid and mass production of Irvingia gabonensis plantlets capable to resist to the hard conditions and therefore, contributes to the protection of plant genetic resources.
The aim of this study is to define in vitro the conditions for regeneration and propagation of Irvingia gabonensis through somatic embryogenesis. The effects of 2,4-D, NAA, BAP and different combinations of BAP/NAA on different stages of somatic embryogenesis has been studied.
MATERIALS AND METHODS
Preparation of explants: Young leaves of Irvingia gabonensis were collected on the apical zone of branches of productive genotype tree (8 years old) found in the Obala region in centre province of Cameroon during the period from July to December of the year 2006. These leaves were washed twice with 1% solution of tween 20 for 10 min each. They were then disinfected in 3% solution of sodium hypochlorite for 20 min and rinsed 5 times with sterile distilled water. The leaves were then cut into pieces (1 cm wide), transversally across the leaf lamina. Leaves fragments were placed on the culture media with the adaxial surface in contact with the medium.
Media and conditions of calli induction: Calli were induced on Murashige and Skoog (MS) medium (1962) modified as followed: half-strength macro salts and chelated iron (MS/2) (Morel and Wetmore, 1951), vitamins 3% sucrose, 400 mg L-1 glutamine and 0.8% agar (Difco). This Basal Medium (BM) was supplemented with 0 (control), 1, 2, 3, 4, 5 and 6 mg L-1 of 2,4-D and its pH was adjusted to 5.8 by addition of NaOH 1 N or HCl 0.1 N. Media were sterilized by autoclaving at 115°C for 30 min under a pressure of 1.6 kg cm-2. The control was the basal medium without 2,4-D. Six leaves were cut into about 50 segments and placed in flasks for each medium. The cultures were maintained at 26±1°C with 16 h photoperiod under fluorescent light at 80 μmol m-2 sec-1. The calli induction percentage was calculated in each medium after 30 days of culture. The growth of calli in different media was expressed by measuring the average fresh weight.
Media of somatic embryogenesis: After 30 days of culture, calli were sub-cultured on the basal medium supplemented with BAP in the range of 0 (control), 1, 2 and 3 mg L-1. Sub-cultures were placed under the same environmental conditions as during calli induction. Embryos were induced after 60 days of sub-culture. The percentage of calli which differentiate somatic embryos and the average number of embryos per callus were evaluated.
Statistical analysis: For all experiment, the control was the basal medium without phytohormon. Fifty explants were used for each calli induction medium and 30 calli were used for each somatic embryogenic medium. All experiences were repeated thrice. The data were analysed using the Duncan`s multiple range tests. Significant differences between treatment means were determined at p<0.05.
Histological studies of embryogenic calli and morphology of somatic embryos: Embryogenic calli were fixed in AFA (mixing of 90 mL of 70°C ethanol, 5 mL of formaldehyde and 5 mL of acetic acid). They were then washed on a tap water current during 24 h and were dehydrated by three successive baths in 90°C ethanol during 30 min each. After three treatments of 30 min each in a mixture of 95°C ethanol and toluene in the respective proportions: 0.75-0.25; 0.5-0.5 and 0.25-0.75, calli were then included in liquefied paraffin during 32 h. Sections of 10 μm thickness were then realised using rotary microtome (Shibuya Optical Co., Ltd). Paraffin was removed on these sections by soaking successively in toluene during 1 h, 95°C alcohol during 1 h and 70°C alcohol during 10 min. They were then rinsed three times in distilled water during 30 min each and stained with haematoxylin of Regaud-Safranine O. Observations were done using photonic microscope (Nikon 233729, Japan) equipped with camera.
Development of somatic embryos: Somatic embryos were isolated from embryogenic calli and sub-cultured on the basal medium without phytohormon during 7 days for weaning. After this stage, somatic embryos were submitted to the second sub-culture in BM supplemented with 2 mg L-1 of BAP during 30 days for germination.
Regeneration of plantlets: Germinated embryos were sub-cultured on the basal medium supplemented with 1, 3, 4 or 5 mg L-1 of BAP and 2 mg L-1 of NAA (BAP/NAA ratio) during 80-90 days for regeneration of plantlets. The average length of plantlets and the average number of leaves and roots per plantlet as growth parameters were measured or counted for each BAP/NAA ratio. For each ratio, 40 germinated embryos were used and the experience was repeated thrice. The significant difference means were determined using Duncan`s multiple range tests at p<0.05.
Acclimatization of plantlets: Plantlets were acclimatized in polyethylene bags containing a sterile mixture soil/vermiculite at equal volume (v/v) under a temperature of 26±1°C, 72-76% of relative humidity and 16 h per day of light (80 μmol m-2 sec-1). During acclimatization, the relative humidity was reduced progressively and plantlets were watered firstly with sterile tap water during 30 days and secondary with tap water during 60 days.
RESULTS
Induction of calli: Callogenesis was initiated from young tender leaves fragments cultured during 30 days on BM supplemented with 2.4-D. The percentage of callogenesis increases from 52.7±2.1% at 1 mg L-1 and reaches the maximum value of 85.1±1.6% at 3 mg L-1 and then this percentage decreases and reaches the lowest value of 13.4 at 6 mg L-1 (Table 1). No calli were initiated on control medium. The fresh weight of calli also increases from 0.33 g at 1 mg L-1 and reaches the maximum value of 0.78 g at 3 mg L-1, then decreases and reaches the lowest value of 0.23 g at 6 mg L-1 (Table 1). Calli induced in the presence of 1-5 mg L-1 of 2,4-D are yellowish, friable and embryogenic (Fig. 1a), while those produced in the presence of 6 mg L-1 of 2,4-D are brownish and non embryogenic. The embryogenic calli are characterised by the presence of green piles of embryogenic nodules (en) (Fig. 1a). The transversal section of embryogenic calli shows that each embryogenic nodule is a pile of cells in division (pcd) (Fig. 1b).
Induction of somatic embryos: Somatic embryos were initiated by sub-culture of embryogenic calli on the basal
| Table 1: | Effects of 2,4-D at various concentrations on the induction of calli from leaves fragments of Irvingia gabonensis after 30 days of culture |
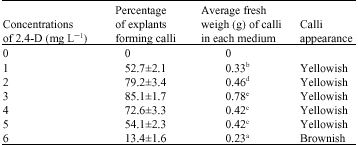 | |
Each value represented the mean±SE of three cultures each with 50 leaves fragments per medium. Means followed by the same letter(s) within columns are not significantly different at p<0.05 according to Duncan`s multiple range test | |
| Table 2: | Effects of BAP at various concentrations on somatic embryos differentiation from calli after 60 days of culture |
 | |
Each value represented the mean±SE of three sub-cultures each with 30 calli per medium. Means followed by the same letter(s) within columns are not significantly different at p<0.05 according to Duncan`s multiple range test | |
Plantlets regeneration and acclimatization: Plantlets were regenerated by sub-culture of germinated somatic embryos on the basal medium supplemented with different ratios of BAP/NAA during 80 days (Table 3). The highest percentage of germinated embryos which develops into
 | |
| Fig 1: | Somatic embryogenesis and regeneration of plantlets of Irvingia gabonensis from leave explants. (a) Embryogenic callus with embryogenic node (ne) obtained from leave fragments cultured on MS/2 supplemented with 3 mg L?1 of 2.4-D during 30 days. (b) Section of embryogenic node showing the piles of cells in division (pcd). (c) Heart-shaped somatic embryos with apical pole (ap) and basal pole (bp) obtained from embryogenic calli sub-cultured on MS/2 supplemented with 1 mg L?1 BAP during 60 days. (d) Section of apical pole of heart-shaped somatic embryos showing the cellular organised zone (coz). (e) Germinated somatic embryos showing the first root (fr) obtained after maintained heart-shaped somatic embryos during 30 days on MS/2 supplemented with 2 mg L?1 BAP. (f) Apex (ax) obtained by the development of the apical pole of germinated somatic embryos on MS/2 supplemented with 4 mg L?1 of BAP and 2 mg L?1 of NAA during 21 days. (g) Plantlets with elongated pivoting roots (epr) obtained on MS/2 supplemented with 4 mg L?1 of BAP and 2 mg L?1 of NAA during 80 days and (h) 90 days olds acclimatized plantlet on sterilized mixture soil vermiculite (v/v) |
| Table 3: | Effect of different ratio of BAP/NAA on the regeneration and growth of plantlets of Irvingia gabonensis from germinated somatic embryos cultured during 80 days |
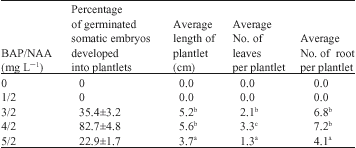 | |
Each value represented the mean±SE of three sub-cultures each with 40 germinated somatic embryos per medium. Means followed by the same letter(s) within columns are not significantly different at p<0.05 according to Duncan`s multiple range test | |
plantlets (82.7±4.8 %) was obtained with 4/2 mg L-1 while the lowest percentage (22.9±1.7%) was obtained with 5/2 mg L-1 ratio (Table 3). The apical pole of germinated embryos develops after 21 days of sub-culture and gives rise to the apex (ax) with the average length of 200±20 mm (Fig. 1f). After 80 days of sub-culture, the apex had developed and gave rise to leafing stem; while on the basal pole the first root elongates and gives rise to pivoting root with average length of 3.5±0.6 cm (Fig. 1g). Then, this elongated pivoting root ramifies and gives rise to root system. Well developed plantlets were obtained with 4/2 mg L-1 BAP/NAA. In fact, with this ratio, each plantlet has the average growth parameter: 5.6 cm of length, 3.3 leaves and 7.2 roots (Table 3).
Under the conditions of acclimatization, regenerated plantlets survive at 93%. They developed during 90 days and gave rise to vigorous dark green plants (Fig. 1h). Each plant at this stage has an average length of 32.4±2.7 cm, presents one or two ramifications and bears an average number of 14.7±2.6 leaves (Fig. 1h).
DISCUSSION
The results of this study show that calli could be initiated successfully from young leaves explants of Irvingia gabonensis cultured on MS/2 supplemented with 2,4-D. During the past decade, 2,4-D has been used to induce calli in some ligneous plants such as Bauhinia vahlii (Dhar and Upreti, 1999), Citrus grandis (Huang et al., 2002), Ricinodendron heudelotii (Fotso et al., 2007). But, the frequency of callogenesis depends not only of the concentration of 2,4-D; also of the type and age of explants. In this study, the maximum percentage of callogenesis (85.1±1.7%) was obtained with 3 mg L-1 of 2,4-D. These results support those obtained by Litz et al. (1998) with Mangifera indica and Huang et al. (2002) with Citrus grandis. Those authors showed that 2,4-D was more effective on callus induction of woody plant explants than naphthalene acetic acid (NAA). Calli induced were embryogenic. In contrast, Hatanaka et al. (1991) and Gill et al. (1995) obtained non embryogenic calli with 2,4-D on Coffea canephora and mandarine explants, respectively. In Irvingia gabonensis, induced calli regenerate somatic embryos after being transferred to differentiation medium supplemented with BAP with the highest percentage of 63.8±2.3 at 1 mg L-1. In most of the reports on in vitro propagation of woody plants by somatic embryogenesis, BAP seems to be one of the best cytokinin used to induce somatic embryos on calli (Omokolo et al., 1997; Jayanthi and Mandala, 2001; Imani et al., 2001; Mohammed et al., 2002). Meanwhile, reports of Zhijian et al. (1998) on Theobroma cacao and Von Arnold et al. (1996) on Picea abies show that the differentiation and development of somatic embryos depend not only of the concentration of BAP in the medium, but also of the others factors such as photoperiods, temperature and relative humidity. In Irvingia gabonensis, obtained results seem to depend to the sum of these factors in addition with 2,4-D which promoted embryogenic calli. In fact, 82.7±4.8% of germinated somatic embryos give rise to plantlets. These results contrast with those obtained by El Maataoui et al. (1990) in Quercus suber and Luo and Koop (1997) in Arabidopsis thaliana. They showed that, only 10-20% of mature somatic embryos obtained in both species give rise to plantlets. In general many reports show that, the success of somatic embryogenesis in higher woody plants depends particularly to the ecotype and genotype of the species studied (Kielly and Bowley, 1992; West and Harada, 1993; Imani et al., 2001). So, the highest percentage of regeneration of plantlets obtained in this study can be also explained by the genotype of the species studied. This regeneration is obtained in this study when BAP is combined whit NAA in the same media. This result permits to approve the existence of complementary and synergic action between auxins and cytokinins as has been reported on somatic embryogenesis in many ligneous species such as Coffea canephora (Hatanaka et al., 1991), Mangifera indica (Litz et al., 1998), Theobroma cacao (Alemano et al., 1997). Acclimatization of plantlets during 90 days in a sterile mixture soil/vermiculite gives 93% of success. This result is similar to those obtained by Omokolo et al. (2004) with plantlets of the same species regenerated by in vitro micro cutting and acclimatized under the same conditions. But, it is higher than the rate of acclimatized plantlets of Dacryodes edulis (51%) obtained by Youmbi and Benbadis (2001) under the same conditions.
In conclusion, with the maxima conditions described at different stage in this study (from callogenesis to acclimatization), it is possible to regenerate through somatic embryogenesis 28±1 survival plantlets of Irvingia gabonensis from a single fragment of leave. So, the present study constitutes a new way for a rapid production of Irvingia gabonensis seedlings with pivoting root system which could be used for the species propagation and domestication. But it is important to use explants from reproductive genotype to pass round the highly heterozygote characteristic of species after regeneration. Further research should be required to determine appropriated stock plant management treatment.
ACKNOWLEDGMENTS
The authors thank Drs. Mbouobda Hermann Desire and Minyaka Emile for technical assistance.
REFERENCES
- Alemanno, L., M. Berthouly and N. Michaux-Ferriere, 1996. Histology of somatic embryogenesis from floral tissues cocoa. Plant Cell. Tissue Organ Cult., 46: 187-194.
CrossRefDirect Link - Blanc, G., L. Lardet, A. Matin, J. Jacob and M.P. Carron, 2002. Differential carbohydrate metabolism conducts morphogenesis in embryogenesis callus of Hevea brasiliensis (Müll Arg.). J. Exp. Bot., 53: 1453-1462.
Direct Link - Das, G. and G.R. Rout, 2002. Direct plant regeneration from leaf explants of Plumbago species. Plant Cell Tissue Organ Cult., 68: 311-314.
CrossRefDirect Link - Dhar, U. and J. Upreti, 1999. In vitro regeneration of a mature leguminous liana (Bauhina vahlii Wight and Arnott). Plant Cell Rep., 18: 664-669.
CrossRefDirect Link - Fotso, T.N., Donfagsiteli, D. Mbouna and N.D. Omokolo, 2007. In vitro Regeneration of Ricinodendron heudelotii. Cahiers d'études et de Recherches Francophones/Agric., 16: 31-36.
Direct Link - Huang, T., S. Peng, G. Dong, L. Zhang and G. Li, 2002. Plant regeneration from leaf-derived callus in Citrus grandis (Pummelo): Effects of auxins in callus induction medium. Plant Cell Tissue Organ Cult., 69: 141-146.
Direct Link - Imani, J., T.L. Thi, G. Langen, B. Arnold-Schmitt and S. Roy et al., 2001. Somatic embryogenesis and DNA organisation of genomes from selected Daucus sp. Plant Cell Rep., 20: 537-541.
Direct Link - Lapido, D.O., J.M. Fondoun and N. Ganga, 1996. Domestication of the Bush Mango (Irvingia sp.): Some Exploitable Intraspecific Variations. In: West and Central Africa: Non-Wood Forest Products No. 9: Domestication and Commercialisation of Non-Timber Forest Products for Agrofestry, Leakey, R.R.B., A.B. Temu, M. Melnyk and P. Vantomme (Eds.). FAO, Rome, Italy, pp: 193-205.
- Lowe, A.J., A.C.M. Gilles, J. Wilson and I.K. Dowson, 2000. Genetics conservation of bush mango from central/west Africa: Implications from random amplified polymorphic DNA analysis. Mol. Ecol., 9: 831-841.
Direct Link - Mohammed, A.M.A., R. Bala and K. Karen, 2002. Somatic embryogenesis in perennial static Limonium bellidifolium, Plumbaginaceae. Plant Cell Tissue Organ Cult., 68: 127-135.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Ndoumou, D.O., G.T. Ndzomo and N. Niemenak, 1997. Phenol content, acidic peroxidase and IAA-oxidase during somatic embryogenesis in Theobroma cacao L. Biol. Plant., 39: 337-347.
CrossRefDirect Link - Shiembo, P.N., A.C. Newton and R.R.B. Leakey, 1996. Vegetative propagation of Irvingia gabonensis, a West African fruit tree. For. Ecol. Manage., 87: 185-192.
Direct Link - Li, Z., A. Traore, S. Maximova and M.J. Guiltinan, 1998. Somatic embryogenesis and plant regeneration from floral explants of Cacao (Theobroma cacao L.) using Thidiazuron. In vitro Cell. Dev. Biol. Plant, 34: 293-299.
CrossRefDirect Link