
A.A. Zayed
Department of Parasitology and Animal Diseases, National Research Center, P.O. Box 12622, Dokki, Giza, Egypt
S. Abdel-Shafy
Department of Parasitology and Animal Diseases, National Research Center, P.O. Box 12622, Dokki, Giza, Egypt
R.M. El-Khateeb
Department of Parasitology and Animal Diseases, National Research Center, P.O. Box 12622, Dokki, Giza, Egypt
Research Journal of Parasitology
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 1-11







ABSTRACT
Scanning electron microscope was used to describe the fine structure of posterior spiracles of third instars of Cephalopina titillator, Oestrus ovis and Rhinoestrus purpureus. The posterior spiracles was found to locate a hallow depressed cuticle of the posterior end of the larval body with an ecdysal scar. The cuticle surrounding the posterior spiracles was provided with spinules and sensory papillae of a taxonomic value. The spinules were numerous and irregularly distributed in dorsal and ventral rows in C. titillator and O. ovis. However, these spinules were few or absent in R. purpureus. The sensory papillae were of different shape; it long and cone-shape in R. purpureus, flat and button-shaped in C. titillator and reduced in O. ovis. The posterior spiracles consisted of two spiracular plates. Each plate appeared strongly sclerotized bearing numerous of respiratory units. The situation of spiracular plate together with ecdysal scar was also of a characteristic taxonomic value. The ecdysal scar surrounded by the spiracular plate either completely in O. ovis or partially in R. purpureus or never surrounded by it in C. titillator but situated medially at the edge of the spiracular plate. The respiratory unit consisted of rima and respiratory slit which appeared a narrow as zigzag-like in O. ovis. This respiratory slit appeared linear not zigzag with a few trabecular tissues in C. titillator. However, this slit was covered with a cap-like structure in R. purpureus. It was concluded that the posterior spiracles of nasal bots have a characteristic line structures of taxonomic value, which may be used for differentiation of these larval species.
PDF Abstract XML References Citation
How to cite this article
A.A. Zayed, S. Abdel-Shafy and R.M. El-Khateeb, 2008. Surface Ultrastructure of Posterior Abdominal Spiracles of Third Instars of Nasal Bots of Cephalopina titillator, Oestrus ovis and Rhinoestrus purpureus (Diptera:Oestridae) Infesting Camels, Sheep and Donkeys in Egypt. Research Journal of Parasitology, 3: 1-11.
URL: https://scialert.net/abstract/?doi=jp.2008.1.11
URL: https://scialert.net/abstract/?doi=jp.2008.1.11
INTRODUCTION
Nasopharyngeal myiasis is caused by some obligatory parasitic larvae of dipterous belonging to order Oestridae (Zumpt, 1965). These larvae include sheep nasal botfly, Oestrus ovis, camel nasal botfly Cephalopina titillator and equine nasal botfly, Rhinoestrus purpureus (Hall and Wall, 1995). Such dipterous parasites are widely distributed in Egypt (Abd El-Aziz, 1988; Zayed et al., 1993, 1994) and many countries of the world (Hall and Wall, 1995). During their life cycle, O. ovis larvae develop in the head sinuses and nasal passages of sheep and goats in all sheep farming areas of the world. Moreover, C. titillator larvae were found to locate labyrinth of ethmoid bone and turbinates of nasal cavity (Zayed, 1998). However, R. purpureus larvae locate labyrinth of ethmoid bone and sphenopalatine communication (Zayed and Hilali, 1993). Since, there are differences between the head size of sheep, camel and equines, the size and shape of the corresponding ethmoid bones and turbinates exhibit also some varieties. These anatomical criteria together with the differences in normal respiratory rates at rest between these animal hosts may have influence on the oxygen tension inside the nasal cavity of the animal species to another. Therefore, the respiratory system especially those posterior spiracles of these larvae may have some modification differences correlated to the animal host and the oxygen tension in their nasal cavity.
The available literatures revealed that few contributions dealing with ultrastructural of posterior spiracles of oestrids larvae were found. A single superficial description for O. ovis larvae by scanning electron microscope was done by Giannetto et al. (1999). They described 1st, 2nd and 3rd instar larvae of O. ovis and mentioned that the posterior spiracles of O. ovis appeared strongly sclerotized and surround the outlet the respiratory canal completely. The respiratory holes of 3rd instars are small and prominent than those of 2nd instar larvae. However, Fahmy (1991) illustrated some details for respiratory spiracles of 3rd larval instars of C. titillator by scanning electron microscope. He found that the spiracles exhibited several irregularly scattered zigzag-like tracheal gills and concluded that this structure is specific shape for these larval stages. Unfortunately, no previous description for posterior spiracles of R. purpureus larvae was found except the preliminary scanning electron microscopy of Guitton et al. (1996) for R. usbekistanicus larvae recovered from donkeys in Senegal.
The main target of this study, therefore, is to describe at comparison the fine structure of posterior spiracles of 3rd larval instars of oestrids including O. ovis of sheep, C. titillator of camels and R. purpureus of equines using scanning electron microscope. Adaptations of these fine structures for its parasitism are also undertaken.
MATERIALS AND METHODS
Collection of Larvae
Third instar larvae of Oestrus ovis, Cephalopina titillator and Rhinoestrus purpureus were directly collected from nasal cavities of infested sheep, camels and donkeys, respectively. The larvae were collected during spring season of 2004, a season of peak infestation with 3rd instars of these larvae (Abd El-Aziz 1988; Zayed et al., 1994, 1993). Sheep and camels were slaughtered at the main abattoir of Cairo (El-Basateen). However, the donkeys were slaughtered at slaughterhouse of National Circus, Giza, Egypt.
Scanning Electron Microscopy (SEM)
Fresh collected larvae of O. ovis, C. titillator and R. purpureus were first identified immediately after Zumpt (1965) and washed several times with distilled water to remove the contact debris. Larvae of each instar were individually injected with 2.5% glutaraldhyde and immersed in the same fixative (Colwell, 1989; Innocenti et al. 1997). Specimens from the posterior part including the posterior spiracles were prepared from larvae of each instar species. The specimens were then dehydrated throw ascending ethanol series, dried in CO2 critical point drier (Blazzer Union F1-9496 Blazer/Furstentun Liechtenstein, Germany) glued over specimens stubs and coated with 20 nm gold in a sputter coater (S150A Sputter Coater Edward, UK). Finally the specimens were examined and photographed with scanning electron microscope (JXA 840, Electron Probe Microanalyzer, Jeol, Japan).
Morphometric Measurements
Permanent specimeltrch for spiraculr plates of 3rd instars of O. ovis, C. titillator and R. purpureus were prepared. The spiraculr plates were first separated from the larval body, cleaned in 10% sodium hydroxide, dehydrated in ascending series of ethanol and mounted in Canada balsam (Zayed and Abd El-Baky, 1999). Measurements of spiraculr plates and respiratory units for 10 specimens for each bots were estimated in micrometers using optical microscopy (PZO,Warszawa, Germany).
RESULTS
SEM demonstrated that the abdominal respiratory spiracles of third instar larvae of Cephalopina titillator, Oestrus ovis and Rhinoestrus purpureus were deeply located in a hallow depressed cuticle at the last posterior end of the larval body. This depressed cuticle formed a dorsal lip (dl) and ventral lip(vl) that jointed to each other medially to form a complete cuticle ring enclosing the posterior spiracles inside (Fig. 1-3). An anal orifice (an) was provided with freshly processes posteriorly. These processes appeared to be dumpy and larger in O. ovis and R. purpureus (Fig. 2-3) than C. titillator larvae (Fig. 1).
The cuticle surrounding the posterior spiracles was provided with both spinules and sensory papillae. The spinules were numerous and regularly arranged in dorsal and ventral rows in C. titillator and O. ovis (Fig. 1-2). These spinules were few or absent in R. purpureus larvae (Fig. 3). The papillae were estimated to be 10 papillae; 4 of them were regularly situated dorsally (dp) above the spinules. The other six papillae were located ventrally, 4 of them (vp) below the posterior spiracles and two anal papillae (ap) below the anal orifice (an) (Fig. 1-3). The cuticle bearing papillae was clearly delineated from the a djoining cuticle forming a raised circular area (ra) in which the papillae originated centrally (Fig. 4-6). SEM indicated that there was a clear morphological difference between papillae of these larvae. It appeared to be a long and conical-shaped in R. purpureus (Fig. 6), flat and button-shaped in C. titillator (Fig. 4) and reduced or rudimentary in O. ovis (Fig. 5).
The posterior spiracles composed of two spiracular plates. Each plate appeared strongly sclerotized bearing numerous respiratory units (ru) and ecdysal scar (es). The ecdysal scar was a cuticular structure with a small hole. It represented the remnant of the previous spiracular opening of 2nd instar larvae after the process of ecdysis. It located medially to the spiracular plate (spl). This paired spiracular plates together with ecdysal scar had a characteristic configuration to distinguish the 3rd larval stages of these nasal bots (Fig. 7-9). The spiracular plates surrounded the ecdysal scar completely in O. ovis (Fig. 8) or partially in R. purpureus (Fig. 9), but never surrounded the ecdysal scar in C. titillator (Fig. 7). The ecdysal scar located at the median edge of each spiracular plate.
The spiracular plates of C. titillator were kidney shaped measuring 784-823 (811.2±4.8)x 470-480 (472±1.3) μm. While, the spiracular plates of O. ovis and R. purpureus were being broader rather than wider measuring 1176-1372 (1313±26.1) x 1048-1117 (1095±9.4) μm and 625-638 (631±1.9) x 713-738 (725.6±3.7) μm, respectively (Table 1).
SEM illustrated that each respiratory plate beard numerous respiratory units (ru). These respiratory units were prominent and scattered irregularly in the spiracular plates (Fig. 10-12). Each respiratory unit was oval to kidney-shaped. Thier long measured 28-37 (32.5±2.9) μm with nearly equal width measured 14-18 μm. Each respiratory unit had a longitudinal respiratory slit and surrounded by rima (rm). The fine structures of respiratory slit were of taxonomic value. It appeared as a narrow zigzag-like with no serration of rima or trabecular tissue in O. ovis (Fig. 14). However. it appeared linear not zigzag having a few irregular trabecular tissues with no serration of rima in C. titillator (Fig. 13). While the respiratory slit in R. purpureus was linear as C. titillator but exhibited a cap-like structure covering the slit (Fig. 15).
The number of respiratory units in each spiracular plate showed a significant difference between the larval instars to another. The spiracular plate of O. ovis showed to have the largest number of respiratory units [1389-1620 (1475±28.39)/plate] followed by R. purpureus [295-325 (310±3.69) / plate] and C. titillator [137-160 (148±2.52) /plate].
 | |
| Fig. 1-3: | SEM micrographs of posterior abdominal spiracles of 3rd instar larvae of C. titillator, O. ovis and R. purpureus (in front view) showing anal orifice (an), anal papillae (ap), dorsal and ventral spinules (asterisk), dorsal lip (dl), dorsal papillae (dp), fleshy processes (fr), Spiracular plate (spl), ventral lip (vl) and ventral papillae (vp). (1): C. titillator spiracles note the absence of fleshy process (fr). (2): O. ovis spiracles note larger dumpy shaped and fleshy process (fr). (3) R. purpureus spiracles note the absence of dorsal (dl) and ventral lips (vl) and large numerous dumpy-shaped fleshy processes (fr) |
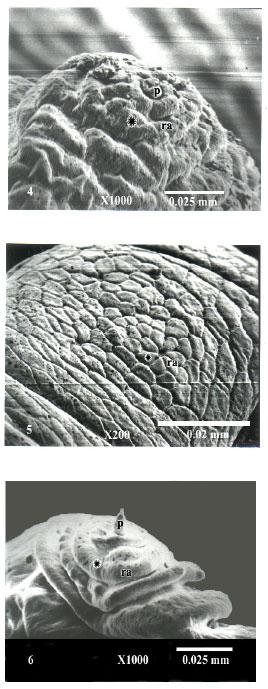 | |
| Fig. 4-6: | SEM micrographs of papillae distributed at the posterior spiracles of 3rd instar larvae of C. titillator, O. ovis and R. purpureus showing delineated cuticle (asterisk), raised circular area from which the sensory papillae originated centrally (ra) and papillae (p). (4): C. titillator papillae note the button-shaped papillae (p). (5): O. ovis papillae note reduced or absence of papillae. (6): R. purpureus papillae, note the cone-shaped papillae |
 | |
| Fig. 7-9: | SEM micrographs of spiraclular plate of 3rd instar larvae of C. titillator, O. ovis and R. purpureus showing ecdysal scar (es) and numerous respiratory units. (7): C. titillator notes no surrounding of spiracular plates (spl) to ecdysal scar (es). (8): O. ovis note the spiracular plate (spl) completely surrounded the ecdysal scar (es). (9): R. purpureus note the plate partially (spl) surrounded the ecdysal scar (es) |
 | |
| Fig. 10-12: | SEM micrographs of respiratory units (ru) of 3rd instar larvae of C. titillator, O. ovis and R. purpureus showing irregular situation of these units inside the spiracular plate. (10): C. titillator note the oval to kidney shaped respiratory units. (11): O. ovis note the oval-shaped respiratory units. (12): R. purpureus note the kidney-shaped respiratory units |
 | |
| Fig. 13-15: | SEM micrographs of fine structure of respiratory units of C . titillator, O. ovis and R. purpureus show the respiratory slits (t) and rima (rm). (13): C. titillator note the respiratory slit not zigzag line appearance having irregular trabecular tissue with no serration of rima (14): O. ovis note the respiratory slit zigzag-like. (15) R. purpureus note the respiratory slit not zigzag-like with a cap like structure over the slit (asterisk) |
| Table 1: | Morphometric measurements of spiracular plates and respiratory units of third larval instars of Cephalopina titillator, Rhinoestrus purpureus and Oestrus ovis and its respiratory unit numbers |
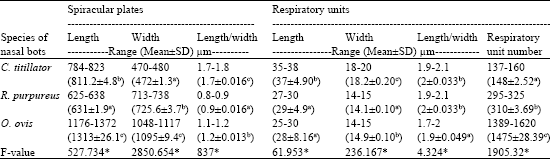 | |
| a, b and c letter(s) showing the significant differences between the measurements or number according Duncan test, *: Significant at (p<0.05) | |
DISCUSSION
This study described in details the fine structure of the posterior abdominal spiracles of 3rd instars of nasal bots; Cephalopina titillator, Oestrus ovis and Rhinoestrus purpureus. This performance allows us to differentiate these nasal flies. The previous literatures revealed no a comprehensive study was done on the posterior spiracles of C. titillator and R. purpureus larvae. However, few superficial studies were reported describing the whole larvae of O. ovis using scanning electron microscope (Guitton and Dorchies, 1993; Giannetto et al., 1999). Although, there was a single scanning electron microscopy study on R. purpureus usbekistanicus with simple description of posterior spiracle of the 3rd instar larvae, no previous work was performed on R. purpureus (Giannetto et al., 1999).
This study illustrated several fine structures on the posterior spiracles that could be used to identify these obligatory dipterous nasal bots. As reported by Fahmy (1991), Principato and Tosti (1988), Giannetto et al. (1999) and Guitton et al. (1996), the architecture of these posterior spiracles were morphologically similar to any other dipterous larvae. It located in a hallow depressed cuticle at the last posterior end of the larval body forming a dorsal and ventral lips jointed to each other enclosing a pair of spiracular plates inside. These cuticular lips were considered the first block barrier that prevented the adverse environmental conditions such as inflammatory exudates in case of rhinitis or high flow of cold air during breathing especially in racing horses. These unfavorable conditions lead the larvae to close the spiracles by these mobile lips. This adaptation of protection mechanism was similar to interpretation reported by Principato and Tosti (1988) for Gasterophilus larvae.
Present study demonstrated that the cuticle surrounding the posterior spiracles was provided with spinules and sensory papillae. The spinules were irregularly arranged in dorsal and ventral rows in C. titillator and O. ovis. However, these spinules were few or absent in R. purpureus. Moreover, the present study reported that the fine structure of the sensory papillae was also distinctly different and provided with additional features that could be used to identify these larval species morphologically. It was large and cone-shaped in R. purpureus, flat and button-shaped in O. ovis and reduced in C. titillator. These fine structures have not been previously described.
SEM revealed that the situation of spiracular plate together with its ecdysal scar had a characteristic configuration in differentiation of these larval species. The ecdysal scar was surrounded by the spiracular plate completely in O. ovis, or partially in R. purpureus but never surrounded by it in C. titillator. Similar observations were reported for Hypoderma species by Colwell et al. (1998). They found that the ecdysal scar was surrounded by the spiracular plate completely in H. actaeon and never surrounded by it in H. diana but the ecdysal scar located it at its median edge.
The present study demonstrated that spiracular plate exhibited numerous respiratory units or holes which being prominent and scattered irregularly inside it. This result was in agreement with that reported by Fahmy (1991) for C. titillator, Guitton et al. (1996) for R. usbekistanicus and Giannetto et al. (1999) for O. ovis.
This study found that the number of the respiratory units was of a significant taxonomic value. It estimated the largest number in O. ovis (1475/plate) followed by R. purpureus (310/plate) and C. titillator (148/plate). Moreover, SEM described in details the morphological features of the respiratory unit, which appeared oval to kidney-shaped having a longitudinal slit surrounded by rima. The fine structures of the respiratory slits were also of taxonomic value. It was zigzag-like with no serration of rima in O. ovis, linear or not zigzag-like having a few trabecular tissues without serration of rima in C. titillator. However, the slit was covered by a cap-like structure in R. purpureus. The results of C. titillator were similar to that reported by Fahmy (1991) except that the respiratory slit appeared a zigzag-like. This difference may be due to that the respiratory slit in C. titillator was wider and provided with trabecular tissue, which may be or not as zigzag-like in its appearance. Unfortunately, no previous description of the respiratory slits of O. ovis and R. purpureus were found.
Further surface ultrastructural studies were still needed for the posterior spiracles of first and second larval instars of these nasal bots to clarify the actual taxonomic differences between these larval species.
REFERENCES
- Colwell, D.D., 1989. Scanning electron microscopy of the posterior spiracles of cattle grubs Hypoderma bovis and Hypoderma lineatum. Med. Vet. Entomol., 3: 391-398.
CrossRefDirect Link - Colwell, D.D., F.J. Martinez-Moreno, A. Martinez-Moreno, S. Hernandez-Rodriguez et al., 1998. Comparative scanning electron microscopy ofthird-instar Hypoderma sp. (Diptera: Estridae). Med. Vet. Entomol., 12: 181-186.
PubMed - Giannetto, S., V. Santoro and S. Pampiglione, 1999. Scanning electron microscopy of Oestrus ovis larvae (Diptera: Oestridae): Skin armour and posterior spiracles. Parasite, 6: 73-77.
CrossRefDirect Link - Guitton, C., P. Dorchies and S. Morand, 1996. Scanning electron microscopy of larval instars and imago of Rhinoestrus usbekistanicus Gan, 1947 (Oestridae). Parasite, 3: 155-159.
CrossRefDirect Link - Hall, M. and R. Wall, 1995. Myiasis of humans and domestic animals. Adv. Parasitol., 35: 257-334.
CrossRefPubMedDirect Link - Innocenti, L., P. Lucchesi and F. Giorgi, 1997. Integument ultrastructure of Oestrus ovis (L.) (Diptera:Oestridae) larvae: Host immune response to various cuticular components. Int. J. Parasitol., 27: 495-506.
CrossRef - Principato, M. and M. Tosti, 1988. Scanning electron microscope observations on the anterior thoracic and post-abdominal spiracles of Gasterophilus larvae (Diptera: Gasterophilidae). Int. J. Parasitol., 18: 191-196.
CrossRefDirect Link - Zayed, A.A. and M. Hilali, 1993. Studies on Rhinoestrus purpureus larvae infesting donkeys in Egypt. J. Equine Vet. Sci., 13: 92-95.
CrossRef - Zayed, A.A., M. Hilali and T.M. El Metenawy, 1993. Studies on Rhinoestrus purpureus (Diptera: Oestridae) larvae infesting donkeys (Equus asinus) in Egypt: Incidence and seasonal variations. J. Equine Vet. Sci., 13: 46-49.
CrossRefDirect Link - Zayed, A.A., 1998. Localization and migration route of Cephalopina titillator (Diptera: Oestridae) larvae in the head of infested camels (Camelus dromedaries). Vet. Parasitol., 80: 65-70.
CrossRef