
Ismaeel Bin-Jaliah
Department of Physiology, College of Medicine, King Khalid University, P.O. Box 641, Abha, Aseer, Kingdom of Saudi Arabia
Journal of Medical Sciences
Year: 2008 | Volume: 8 | Issue: 1 | Page No.: 54-61







ABSTRACT
The purpose of this study was to examine the behavior of glutathione peroxidase (GPx) activities and Free Radicals (FR) production in the brain and the lung during graded hyperoxia exposure. Twenty-four adult male rats, matched with age and body weigh, were randomly assigned to four groups. The first group served as control and the second, third and fourth were exposed to hyperoxia for 24, 48 and 72 h, respectively. Following the exposure period for each group animals were sacrificed and both lungs and brain tissues were homogenized for GPx and FR determinations. GPx activity was determined by Randox protocol (Randox, UK) and FR was determined using dROM method (H and D, Italy). Results showed that mean ( ± SD) GPx activity in the lungs increased from the baseline control of 12898.33 ± 6034.77 to 20083.62 ± 2734 (U L-1) during hyperoxia exposure for 24; then dropped to 5467.77 ± 1159.53 and 8271.80 ± 1347.67 (U L-1) during hyperoxia exposure for 48 and 72 h, respectively. Whereas mean ( ± SD) GPx activity in the brain increased from the baseline control of 5467.80 ± 2852.65 to 13841.72 ± 1245.67 and 14594.82 ± 6711.44 (U L-1), during hyperoxia exposure for 24 and 48 h, respectively; then dropped to 4346.17 ± 343.34 (U L-1), during 72 h exposure. The sustained increased in GPx up-to 48 hr in the brain provided evidence of delayed protection against ROS. The average ( ± SD) FR production in the lung increased from the baseline control of 176.67 ± 33.79 to 274.33 ± 33.37, 260.00 ± 62.54 and 320.00 ± 114.91 (U L-1) during hyperoxia exposure for 24, 48 and 72 h, respectively. The average ( ± SD) FR production in the brain increased from the baseline control of 73.33 ± 20.18 to 132.17 ± 21.77 during hyperoxia exposure for 24 h and then dropped to 94.33 ± 14.56 and 65.33 ± 21.12, during 48 and 72 h, respectively. Tukey-Kramer multiple comparisons between the lung and the brain showed that the lungs had higher rate of FR formation at all levels of hyperoxia exposures, which suggest more mechanisms that had contributed to FR formation in pnueumocyte, as compared with neurocyte. Based on the results of the present study antioxidant supplements are recommended for traumatic brain injury and hypoxemia lung injury patients subjected to oxygen therapy.
PDF Abstract XML References Citation
How to cite this article
Ismaeel Bin-Jaliah, 2008. Comparison of Glutathione Peroxidase Activity and Free Radicals Production in the Lungs and the Brain of Rats During Graded Hyperoxia. Journal of Medical Sciences, 8: 54-61.
DOI: 10.3923/jms.2008.54.61
URL: https://scialert.net/abstract/?doi=jms.2008.54.61
DOI: 10.3923/jms.2008.54.61
URL: https://scialert.net/abstract/?doi=jms.2008.54.61
INTRODUCTION
Oxygen toxicity had attracted the attention in physiology since the early work by Bean (1945) and Comroe et al. (1945). Hyperoxia inducts reactive oxygen species (Heberlein et al., 2000; Hitka et al., 2003; Haffor and Al-Johany, 2005) which alter cellular components and thus impair various ionic conductance`s that regulate cell excitability and exchangeability (Colton and Colton, 1986; Upham et al., 1997; Bickford et al., 1999). Studies of hyperoxia confirmed that ROS are reported to target certain neurotransmitter and neuromodulator systems and thus to alter chemical synaptic function (Zhang et al., 1995; Bitterman and Bitterman, 1998; Dean and Mulkey, 2000; Mulkey et al., 2003). Following exposure to hyperoxia, the lungs show morphologic changes that are similar to pulmonary inflammation, atelectasis and oedema formation, leading to irreversible loss of respiratory function. Furthermore, lung inflammation is associated with infiltration of circulating neutrophils (Crapo, 1986; Jankov et al., 2003; Jafari et al., 2004) with potential source for ROS formation.
Glutathione Peroxidase (GPx), is a free radical scavenger, initially reported that neutralized the highly toxic hydroxyl radical (Paglia and Valentine, 1967), an observation which has been confirmed by several authors (Allen and Balin, 2003; Pollack et al., 2005; Al-Johany and Haffor, 2007). Previous studies demonstrated that GPx is critical antioxidant in the peroxidase system of cytochrome-C, quinones and ascrobate that prevented mitochondrial swelling which are accompanied by the production of hydrogen peroxide and malonaldhyde and intermediate of free radicals initiated lipid peroxidation (Hunter et al., 1964; Lesnefsky et al., 2001; Haffor et al., 2002; Haffor, 2004; Haffor and Al-Johang, 2005; Haffor and Alhazza, 2007).
The question that arises is how fast it takes for ROS intermediates to react with membrane lipids and start a series of pathologic antioxidants response in the brain, as compared with the lung tissues. The clinical relevance of this issue relates to the controversial question of whether hyperoxic exposure intervals contribute to irreversible deleterious in the CNS control of pulmonary vascular effects or an earlier direct pneumocyte phagocyte infiltration mechanism. The purpose of this study was to examine the changes in glutathione peroxidase activity (GPx) and free radicals (FR) production in the brain and the lung tissues during graded hyperoxia exposure for 24, 48 and 72 h. In particular, we emphasize the use of hyperoxia under normal baric O2 as in intensive care model for studying the impact on the cellular mechanisms by which acute oxidative stress (ROS) can potentially alters neuronal and pneumocyte physiological activities. Some of this work has been published previously in abstract form (Bin-Jaliah, 2007).
MATERIALS AND METHODS
Experimental design and hyperoxia exposure: The experiments were conducted during the spring of 2007 in the College of Science, King Saud University, as part of collaborative initiative with the aim to establish a High Altitude and Stress Physiology Laboratory (HASPL). Twenty-four adult wister albino male rats, Rattus norvigicus, matched with age and body weigh were randomly assigned to four groups, six animals each. The first group served as control and the second, third and fourth were exposed to hyperoxia for 24, 48 and 72 h, respectively. Animals of the experimental groups were placed in a closed box that has an inlet flow which was connected to 100% O2 tank, medical grade, on which a regulator was connected to maintain flow at 5 L min-1 (LPM). The out flow of the regulator passed through a humidifier in order to saturate the inspired air with H2O. The outlet ventilation rate of the box was adjusted at 5 LPM to ensure that the concentration of oxygen in the box remains equal to 100% O2 and maintain normal flow and normal barometric pressure at 767 mmHg. The temperature inside the box was adjusted at room temperature (22-24° C). Animals of the control and experimental groups were sacrificed and the lungs and brain were isolated and homogenized immediately in 0.9 saline solutions (4:1 ratio).
Glutathione peroxidase: Samples from all groups were used for the determination of GPx activities using Randox protocol (Randox, England). This method is based on the detoxification of hydrogen peroxide by the oxidation of reduced glutathione according to the following reaction:
Free radical determination: Free radicals production was measured, using the d-ROMs-2 test kits (Health and Diagnostic, Italy) according to the manufacturer`s instructions. The test measures the levels of hydroperoxides (R-OOH) which are generated by peroxidation of biological compounds; lipid, amino acids, nucleic acids. This test is based on the principle of the ability of hydrogen peroxides to generate free radicals after reacting with some transitional metals (Fe2+/Fe3+), according to Fenton`s Reaction as follows:
H2O2 + Fe++ = *OH + OH– + Fe++ |
Thus, the hydrogen peroxides of biological sample (whole blood) generate free radicals (alcoxy and peroxyl radicals) after exposure to a transitional metal (Fe++/Fe+++). When a correctly buffered chromogen substance (N, N-diethyl-phenylenediamine) lead to the reduction of hydrogen peroxides which in turns colored as radical cation. Color intensity was read using spectrophotometer with peak absorbance of 505 nm. In the d-ROMs test, results were expressed in CARR units (CARR U). One CARR U relates to 0.08 mg H2O2/100 mL.
Statistical analysis: Mean group differences for the dependent variables; glutathione peroxidase (GPx) and free radicals (FR) were evaluated using one-way analysis of variance (ANOVA) to reveal the main effect of each group on the dependent variables. Tukey-Kramer multiple comparisons were used to compare differences between each means pairs.
RESULTS
The mean final body weights ( ± SD) of the four groups; control, hyperoxia-24 h, hyperoxia-48 h and hyperoxia-72 h; at the end of the experiment were 185.67 ± 6.41, 188.66 ± 11.39, 192.33 ± 10.09 and 186.83 ± 12.35 g, respectively. Results of paired t-test indicated that values were not significantly (p>0.05) different from body weights prior to the experiment.
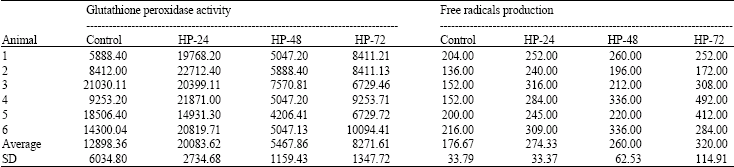
Exposure to hyperoxia resulted in increasing mean ( ± SD) GPx activity from the baseline control of 12898.33 ± 6034.77 to 20083.62 ± 2734 (U L-1), during hyperoxia exposure for 24; then dropped to 5467.77 ± 1159.53 and 8271.80 ± 1347.67 (U L-1), during hyperoxia exposure for 48 and 72 h, respectively. The average ( ± SD) FR production in the lung increased from the baseline control of 176.67 ± 33.79 to 274.33 ± 33.37, 260.00 ± 62.54 and 320.00 ± 114.91 (CARR) during hyperoxia exposure for 24, 48 and 72 h, respectively (Table 1).
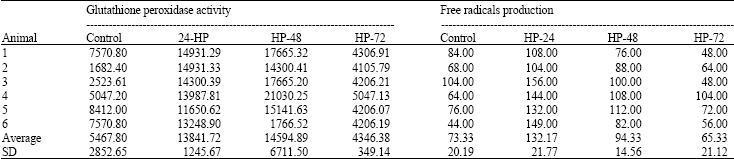
GPx activity in the brain increased, from the mean baseline control of 5467.80 ± 2852.65 to 13841.72 ± 1245.67 and 14594.82 ± 6711.44 (U L-1), during hyperoxia exposure for 24 and 48 h, respectively; then dropped to 4346.17+343.34 (U L-1), during 72 h exposure. The mean ( ± SD) FR production in the brain increased from the baseline control of 73.33 ± 20.18 to 132.17 ± 21.77 during hyperoxia exposure for 24 h and then dropped to 94.33 ± 14.56 and 65.33 ± 21.12, during 48 and 72 h, respectively (Table 2).
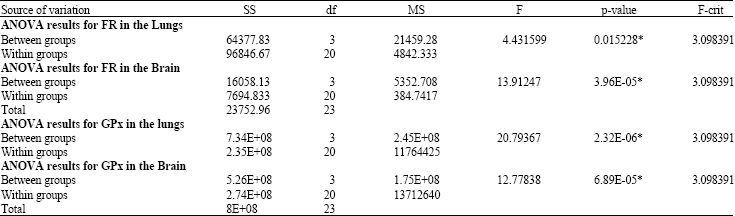
Results of one-way (ANOVA) analysis of variances (Table 3) showed significant (p<0.05) differences among groups` means in both lungs and brain for glutathione peroxidase (GPx) activities and free radicals (FR) productions. Post-hoc Tukey-Kramer multiple comparisons procedures (Table 4) were conducted to simultaneously examine comparisons between all possible pairs of group means. When hyperoxia exposure administered for 24 h, it elevated GPx activity significantly (p<0.05), then dropped significantly (p<0.05) at 48 h in the lungs and remained lower following 72 h exposure but the difference was not statistically significantly (p>0.05) lower than control group. In the brain, when hyperoxia was administered for 24 h, it elevated GPx significantly (p<0.05) and remained higher significantly (p<0.05) following hyperoxia exposure for 48 h, then dropped to control value following 72 h of hyperoxia exposure. Despite the observed progressive rise in FR production in the lungs, with increasing hyperoxia exposure but the difference was significantly (p<0.05) higher following hyperoxia exposure for 72 h only. In the brain, when hyperoxia administered for 24 h FR production increased significantly (p<0.05), then dropped following exposure for 48 and 72 h, but the difference was not statistically significant (p>0.05), as compared with control group. Figure 1a and b displays and summarizes the behavioral mean changes for GPX activity and FR production in the lungs and the brain during control and hyperoxia exposure for 24, 48 and 72 h.
| Table 1: | Glutathione peroxidase activity (U L-1) and free radicals production (Carr) in the lungs |
 | |
| Table 2: | Glutathione peroxidase activity (U L-1) and free radicals production (Carr) in the brain |
 | |
| Table 3: | One way analysis of variance (ANOVA) results |
 | |
| *: p<0.05 | |
| Table 4: | Tukey-Kramer Multiple comparisons for GPx and FR in the lungs and brain |
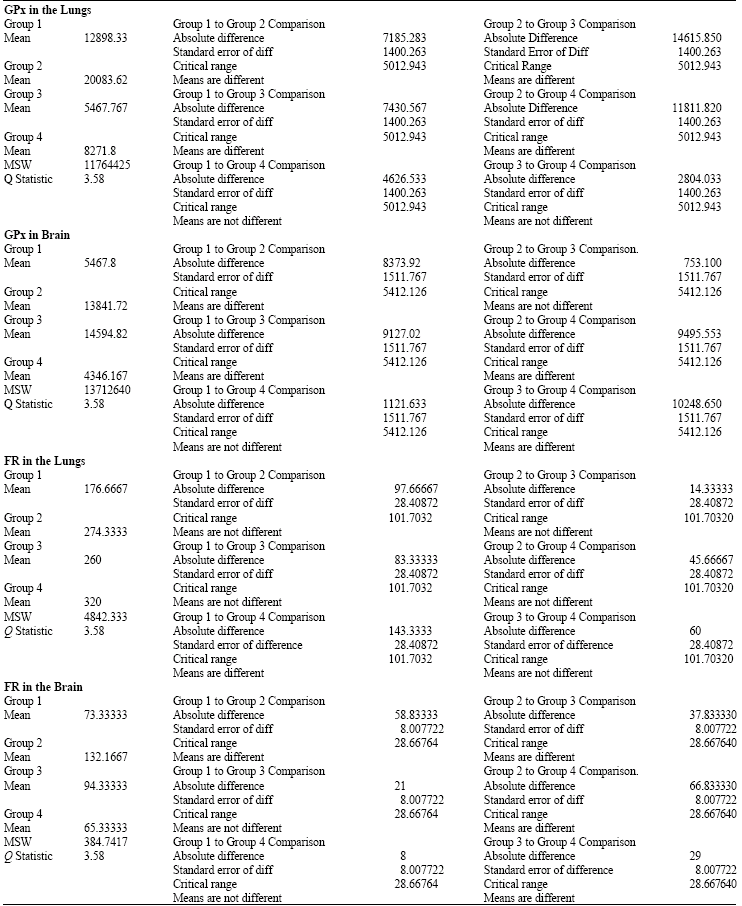 | |
| *Group 1 = Control, Group 2 = Hyperoxia exposure for 24 h, Group 3 = Hyperoxia exposure for 48 h and Group 4 = hyperoxia exposure for 72 h | |
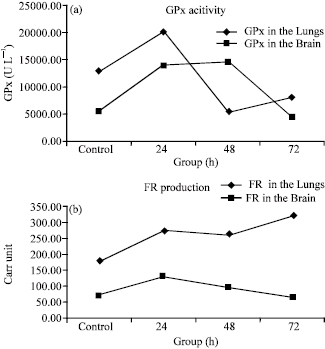 | |
| Fig. 1: | The Behavior of mean GPx in the lungs and brain (a) and FR production in the lungs and brain (b) during hyperoxia exposure |
DISCUSSION
Hyperoxia is believed to generate Reactive Oxygen Species (ROS) and inhibit antioxidants defense. O2 toxicity is believed to occur when the body`s antioxidant defenses are overwhelmed by increased production of ROS. Included in ROS list are superoxide, hydrogen peroxide, hydroxyl radicals and peroxynitrite at high levels of PtiO2 (Demchenko et al., 2001, 2003; Elayan et al., 2000; Torbati et al., 1992). Herein, the present study showed that glutathione peroxidase (GPx) in the lungs was overwhelmed following 24 h of hyperoxia exposure. In the brain, GPx was overwhelmed after 48 h of hyperoxia exposure.
It is clear that dysfunctional mitochondria result in releasing its contents such as oxidative enzymes and hydrogen peroxides to the cytoplasm, in attempt to prevent swelling. When the rate of release of mitochondrial contents exceeds elimination rate of antioxidants defense system, ROS accumulate and FR productions rise. GPx catalyzes specifically detoxification of hydrogen peroxide (Paglia and Valentine, 1967). The severity of hyperoxic-induced cellular injury is time and dose dependent (Hayatdavoudi et al., 1981; Barry and Crapo, 1985; Crapo et al., 1994).
The effects of hyperoxia on the central nervous system (CNS) and risk of bronchopulmonary dysplasia in infants or adult respiratory distress syndrome in adults begins with exposure period over 8 hours (Arieli, 1998; Chavko et al., 1998; Demchenko et al., 2001). In healthy adult risk begins after 48 h (Comroe et al., 1945). Acute exposure to hyperoxia causes tissue and cellular damages in the brain (Huang et al., 2000; Gerstner et al., 2006). In addition, ethanol intake is associated with adaptive changes in the antioxidant defense enzymes such as increased levels of neural superoxide dismutase and catalase (Montoliu et al., 1994). Furthermore, chronic ethanol intake decreases the levels of the GSH (Guerri and Grisolia, 1980; Montoliu et al., 1994) and tocopherol content of the cerebellum (Rouach et al., 1991). Many clinicians have successfully reduced oxygen toxicity by the antioxidant redoxiredoxin administration (Kim et al., 2003) and selenium supplementation (Ebert et al., 2006). Others used nitro oxide to prevent cell apoptosis in the lungs (Iben et al., 2000; Howlett et al., 1999). In the present study, GPx sustained its rise in attempt to defend the integrity of the pneumocyte till 24 h of hyperoxia exposure and up-to 48 h in the brain.
Although increased generation of ROS is evident in lung epithelia cells in vitro within 30-60 min of hyperoxia (Manautou and Carlson, 1991; Sanders et al., 1993; Parinandi et al., 2003), clinical use of normobaric hyperoxia for several hours is frequently considered harmless or even recommended to reduce the risk of post-surgical wound infections (Neubauer et al., 1994; Greif et al., 2000; Belda et al., 2005) and head injury (Brown et al., 1988). In humans, the first respiratory symptoms have been reported after 6 h of oxygen exposure (Comroe et al., 1945) and ultrastructural alterations such as epithelial cell swelling are evident within 14 h but with hyperoxia of 70% O2 (Kapanci et al., 1972). In rats, animals die within 60-72 h of exposure to 100% O2, whereas an FIO2 of 0.85 is sublethal but may cause platelet accumulation within 3 days and increase lung weight within 5 days of exposure (Crapo et al., 1980; Barry and Crapo, 1985; Tibbles and Edelsberg, 1996). Herein, results of the present study clearly showed reduction in GPx and higher rate of FR generation beyond 24 h of hyperoxia exposure (100% O2, medical grade) but did not result in animals` death up-to 72 h.
The severity of hyperoxic lung injury is time- and dose dependent (Crapo et al., 1980; Barry and Crapo, 1985). As compared with 95% O2 , an FIO2 of 0.7 results in less ROS formation but does not protect from lung injury when applied over longer periods because rats exposed to 60% O2 for 7 days showed reduced lung compliance and perivascular oedema formation (Cragg et al., 1986; Caldwell et al., 1966; Nishio et al., 1998) and 14 day exposure to 60% O2 causes low-grade epithelial injury and interstitial fibrosis in baboons (Crapo et al., 1994). Besides lung epithelial ROS generation, capillary endothelial cells were identified as the source of hyperoxia-induced ROS production (Kuebler et al., 2000). Herein, results of the present study showed a steady rise in ROS generation, along with increasing exposure period, yet beyond the onset of lung tissue injury at 24 h, which reflected additional phagocytes defense mechanism of the alveolar macrophages which in turn contributed additively to ROS generation.
CONCLUSIONS
The results of the present study along with the data showing that GPx and FR levels increased in the lungs and the brain tissues but these changes were delayed and reversed in the brain. Steady elevated of ROS formation, with increasing exposure period, in the lungs reflected added pulmonary vascular effects directly related to pneumocyte phagocyte infiltration mechanism, perhaps ROS-related chemotaxic mechanism.
ACKNOWLEDGMENT
The author thanks Dr. Al-Said Haffor from the College of Science, King Saud University for his sincere help in data management, analysis and review of this manuscript.
REFERENCES
- Al-Johany, A.M. and A.S.A. Haffor, 2007. Effects of cold temperature on the activities of glutathione peroxidase, lactate dehydrogenase and free radicals production in uromastyx aegyptids. J. Medical Sci., 7: 408-412.
CrossRefDirect Link - Allen, R.G. and A.K. Balin, 2003. Effects of oxygen on the antioxidant responses of normal and transformed cells. Exp. Cell Res., 289: 307-316.
Direct Link - Arieli, R., 1998. Latency of oxygen toxicity of the central nervous system in rats as a function of carbon dioxide production and partial pressure of oxygen. Eur. J. Applied Physiol. Occup. Physiol., 78: 454-459.
CrossRefDirect Link - Barry, B.E. and J.D. Crapo, 1985. Patterns of accumulation of platelets and neutrophils in rat lungs during exposure to 100% and 85% oxygen. Am. Rev. Respir. Dis., 132: 548-555.
CrossRefPubMedDirect Link - Belda, F.J., L. Aguilera, A. Garcia, J. Alberti, R. Vicente and L. Ferrandiz et al., 2005. Supplemental perioperative oxygen and the risk of surgical wound infection: A randomized controlled trial. J. Am. Med. Assco., 294: 2035-2042.
Direct Link - Bickford, P.C., K. Chadman, B. Williams, B. Shukitt-Hale, D. Holmes, G. Taglialatela and J. Joseph, 1999. Effect of normobaric hyperoxia on two indexes of synaptic function in fisher 344 rats. Free Radic. Biol. Med., 26: 817-824.
CrossRefDirect Link - Bitterman, N. and H. Bitterman, 1998. L-Arginine-NO pathway and CNS oxygen toxicity. J. Applied Physiol., 84: 1633-1638.
Direct Link - Brown, J.A., M.C. Preul and A. Taha, 1988. Hyperbaric oxygen in the treatment of elevated intracranial pressure after head injury. Pediatr. Neurosci., 14: 286-290.
CrossRefDirect Link - Caldwell, P.R.B., W.L. Lee, H.S. Schildkraut and E.R. Archibald, 1966. Changes in lung volume, diffusing capacity and blood gases in men breathing oxygen. J. Applied Physiol., 21: 1477-1483.
Direct Link - Chavko, M., J.C. Braisted and A.L. Harabin, 1998. Effect of MK-801 on seizures induced by exposure to hyperbaric oxygen: Comparison with AP-7. Toxicol. Applied Pharmacol., 151: 222-228.
CrossRefDirect Link - Colton, C.A. and J.S. Colton, 1986. The action of oxygen and oxygen at high pressure on inhibitory transmission. Brain Res., 364: 151-158.
CrossRefDirect Link - Comroe, Jr. J.H., R.D. Dripps, P.R. Dumke and M. Deming, 1945. Oxygen toxicity: The effect of inhalation of high concentrations of oxygen for twenty-four hours on normal men at sea level and at a simulated altitude of 18,000 feet. JAMA., 128: 710-717.
CrossRefDirect Link - Cragg, P.A., D.B. Drysdale and J.H. Hamilton, 1986. Ventilation in intact and glossopharyngeal nerve sectioned anaesthetized rats exposed to oxygen at high pressure. J. Physiol., 370: 489-499.
CrossRefDirect Link - Crapo, J.D., B.E. Barry, H.A. Foscue and J. Shelburne, 1980. Structural and biochemical changes in rat lungs occurring during exposures to lethal and adaptive doses of oxygen. Am. Rev. Respir. Dis., 122: 123-143.
CrossRefDirect Link - Crapo, J.D., 1986. Morphologic changes in pulmonary oxygen toxicity. Annu. Rev. Physiol., 48: 721-723.
CrossRefDirect Link - Crapo, J.D., G. Hayatdavoudi, M.J. Knapp, P.J. Fracica, W.G. Wolfe and C.A. Piantadosi, 1994. Progressive alveolar septal injury in primates exposed to 60% oxygen for 14 days. Am. J. Physiol., 267: L797-L806.
Direct Link - Dean, J.B. and D.K. Mulkey, 2000. . Continuous intracellular recording from mammalian neurons exposed to hyperbaric helium, oxygen, or air. J. Applied Physiol., 89: 807-822.
Direct Link - Demchenko, I.T., A.E. Boso, A.R. Whorton and C.A. Piantadosi, 2001. Nitric oxide production is enhanced in rat brain before oxygen-induced convulsions. Brain Res., 917: 253-261.
Direct Link - Demchenko, I.T., D.N. Atochin, A.E. Boso, J. Astern, P.L. Huang and C.A. Piantadosi, 2003. Oxygen seizure latency and peroxynitrite formation in mice lacking neuronal or endothelial nitric oxide synthases. Neurosci. Lett., 344: 53-56.
Direct Link - Ebert, R., M. Ulmer, S. Zeck, J. Meissner-Weigl, D. Schneider, H. Stopper, N. Schupp, M. Kassem and F. Jakob, 2006. Selenium supplementation restores the antioxidative capacity and prevents cell damage in bone marrow stromal cells in vitro. Stem Cells., 24: 1226-1235.
Direct Link - Elayan, I.M., M.J. Axley, P.V. Prasad, S.T. Ahlers and C.R. Auker, 2000. Effect of hyperbaric oxygen treatment on nitric oxide and oxygen free radicals in rat brain. J. Neurophysiol., 83: 2022-2029.
Direct Link - Gerstner, B., C. Buhrer, C. Rheinlander, O. Polley and A. Schuller et al., 2006. Maturation-dependent oligodendrocyte apoptosis caused by hyperoxia. J. Neurosci. Res., 84: 306-315.
Direct Link - Greif, R., O. Akca, E.P. Horn, A. Kurz and D.I. Sessler, 2000. Supplemental perioperative oxygen to reduce the incidence of surgical wound infection. Outcomes research group. N. Engl. J. Med., 342: 161-167.
Direct Link - Guerri, C. and S. Grisolia, 1980. Changes in glutathione in acute and chronic alcohol intoxication. Pharmacol. Biochem. Behavior., 13: 53-61.
CrossRefDirect Link - Haffor, A.S., 2004. Effects of O2 breathing on cardiac mitochondrial, got and free radical production. J. Med. Sci., 42: 164-169.
CrossRefDirect Link - Haffor, A.S. and A.M. Al-Johany, 2005. Effect of heat stress, hypoxia, hypoxia-hyperoxia on free radical production in mice, Mus Musculus. J. Med. Sci., 5: 89-94.
CrossRefDirect Link - Haffor, A.S.A. and I.M. Alhazza, 2007. Effects of lead and hyperoxia on lactate dehydrogenase activity in rats. J. Medical Sci., 71: 452-456.
CrossRefDirect Link - Hayatdavoudi, G., J.J. O'Neil, B.E. Barry, B.A. Freeman and J.D. Crapo, 1981. . Pulmonary injury in rats following continuous exposure to 60% O2 for 7 days. J. Applied Physiol., 51: 1220-1231.
Direct Link - Heberlein, W., R. Wodopia, P. Bartsch and H. Mairbaurl, 2000. Possible role of ROS as mediators of hypoxia-induced ion transport inhibition of alveolar epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol., 278: 640-648.
Direct Link - Hitka, P., M. Vizek and J. Wilhelm, 2003. Hypoxia and reoxygenation increase H2O2 production in rats. Exp. Lung Res., 29: 585-592.
Direct Link - Howlett, C.E., J.S. Hutchison, J.P. Veinot, A. Chiu, P. Merchant and H. Fliss, 1999. Inhaled nitric oxide protects against hyperoxia-induced apoptosis in rat lungs. Am. J. Physiol., 277: L596-L605.
Direct Link - Huang, K.L., J.N. Wu, H.C. Lin, S.P. Mao, B.H. Kang and F.J. Wan, 2000. Prolonged exposure to hyperbaric oxygen induces neuronal damage in primary rat cortical cultures. Neurosci. Lett., 293: 159-162.
Direct Link - Iben, S.C., I.A. Dreshaj, C.F. Farver, M.A. Haxhiu and R.J. Martin, 2000. Role of endogenous nitric oxide in hyperoxia-induced airway hyperreactivity in maturing rats. J. Applied Physiol., 89: 1205-1212.
Direct Link - Jafari, B., B. Ouyang, L.F. Li., C.A. Hales and D.A. Quinn, 2004. Intracellular glutathione in stretch-induced cytokine release from alveolar type-2 like cells. Respirology, 9: 43-53.
Direct Link - Jankov, R.P., L. Johnstone, X. Luo, B.H. Robinso and A.K. Tanswell, 2003. Macrophages as a major source of oxygen radicals in the hyperoxic newborn rat lung. Free Radic. Biol. Med., 35: 200-209.
Direct Link - Kapanci, Y., R. Tosco, J. Eggermann and V.E. Gould, 1972. Oxygen pneumonitis in man: Light-and electron-microscopic morphometric studies. Chest, 62: 162-169.
CrossRefDirect Link - Kim, H.S., Y. Manevich, S.I. Feinstein, J.H. Pak, Y.S. Ho and A.B. Fisher, 2003. Induction of 1-cys peroxiredoxin expression by oxidative stress in lung epithelial cells. Am. J. Physiol. Lung Cell Mol., 285: L363-L369.
CrossRefDirect Link - Kuebler, W.M., K. Parthasarathi, P.M. Wang and J. Bhattacharya, 2000. A novel signaling mechanism between gas and blood compartments of the lung. J. Clin. Invest., 105: 905-913.
Direct Link - Lesnefsky, E.J., S. Moghaddas, B. Tandler, J. Kerner and C.L. Hoppel, 2001. Mitochondrial dysfunction in cardiac disease: Ischemia-reperfusion, aging and heart failure. J. Mol. Cell Cardiol., 33: 1065-1089.
Direct Link - Manautou, J.E. and G.P. Carlson, 1991. Ethanol-induced fatty acid ethyl ester formation in vivo and in vitro in rat lung. Toxicology, 70: 303-312.
CrossRefDirect Link - Montoliu, C., S. Valles, J. Renau-Piqueras and C. Guerri, 1994. Ethanol-induced oxygen radical formation and lipid peroxidation in rat brain: Effect of chronic alcohol consumption. J. Neurochem., 63: 1855-1862.
CrossRefDirect Link - Mulkey, D.K., R.A.III. Henderson, R.W. Putnam and J.B. Dean, 2003. Hyperbaric oxygen and chemical oxidants stimulate CO2/H+-sensitive neurons in rat brain stem slices. J. Applied Physiol., 95: 910-921.
Direct Link - Neubauer, R.A., S.F. Gottlieb and N.H. Pevsner, 1994. Hyperbaric oxygen for treatment of closed head injury. Southern Med. J., 87: 933-936.
PubMedDirect Link - Nishio, K., Y. Suzuki, T. Aoki, K. Suzuki and A. Miyata et al., 1998. Differential contribution of various adhesion molecules to leukocyte kinetics in pulmonary microvessels of hyperoxia-exposed rat lungs. Am. J. Respir. Crit. Care Med., 157: 599-609.
Direct Link - Paglia, D.E. and W.N. Valentine, 1967. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med., 70: 158-169.
CrossRefPubMedDirect Link - Parinandi, N.L., M.A. Kleinberg, P.V. Usatyuk, R.J. Cummings and A. Pennathur et al., 2003. Hyperoxia-induced NAD(P)H oxidase activation and regulation by MAP kinases in human lung endothelial cells. Am. J. Physiol. Lung Cell Mol. Physiol., 284: L26-L38.
Direct Link - Koo, H.C., J.M. Davis, Y. Li, D. Hatzis and H. Opsimos et al., 2005. Effects of transgene expression of superoxide dismutase and glutathione peroxidase on pulmonary epithelial cell growth in hyperoxia. Am. J. Physiol.-Lung Cell Mol. Physiol., 288: L718-L726.
CrossRefDirect Link - Sanders, S.P., J.L. Zweier, P. Kuppusamy, S.J. Harrison, D.J. Bassett, E.W. Gabrielson and J.T. Sylvester, 1993. Hyperoxic sheep pulmonary microvascular endothelial cells generate free radicals via mitochondrial electron transport. J. Clin. Invest., 91: 46-52.
CrossRef - Tibbles, P.M. and J.S. Edelsberg, 1996. Hyperbaric-oxygen therapy. N. Engl. J. Med., 334: 1642-1648.
CrossRefDirect Link - Torbati, D., D.F. Church, J.M. Keller and W.A. Pryor, 1992. Free radical generation in the brain precedes hyperbaric oxygen-induced convulsions. Free Radic. Biol. Med., 13: 101-106.
CrossRef - Upham, B.L., K.S. Kang, H.Y. Cho and J.E. Trosko, 1997. Hydrogen peroxide inhibits gap junctional intercellular communication in glutathione sufficient but not glutathione deficient cells. Carcinogenesis, 18: 37-42.
CrossRefDirect Link - Zhang, J., A.D. Sam, B. Klitzman and C.A. Piantadosi, 1995. Inhibition of nitric oxide synthase on brain oxygenation in anesthetized rats exposed to hyperbaric oxygen. Undersea. Hyperb. Med., 22: 377-382.
PubMed