
Hashem Al-Sheikh
Department of Biology, College of Science, King Faisal University, Al-Hassa, Saudi Arabia
Hani M.A. Abdelzaher
Department of Biology, College of Sciences, Al-Jouf University, Sakaka, Saudi Arabia
Research Journal of Microbiology
Year: 2012 | Volume: 7 | Issue: 2 | Page No.: 82-100







ABSTRACT
Increase the productivity of crops is a matter of concern for workers in the field of crop science and plant diseases. Pythiaceous fungi are one of the biggest sources of injury to seedling crop plants, especially if environmental conditions were not favorable for plant growth and favorable for the disease prevalence. Seventy four isolates of virulent Pythium aphanidermatum were obtained from rhizosphere soil of maize, potato, tomato and wheat plants cultivated in five farms in Al-Jouf and a farm in Al-Hassa, Saudi Arabia. Sequencing of rDNA-ITS of the isolated P. aphanidermatum including the 5.8SrDNA were analyzed to confirm the species identification. Isolates of P. aphanidermatum started to produce zoospores after 72 h incubation at 20, 25, 30, 35 and 40°C. Production of zoospores was inhibited at pH 4 and prompted within the pH range 5-8 with optimum at pH 7. All isolates were able to produce zoospores between -0.13 to -0.27 MPa but not above this range. The highest zoospore production was found at -0.27 MPa but it was inhibited by 0.5% NaCl. The optimum concentrations for zoospores production were between 0.1 and 0.2% gm NaCl. Oospores produced between 10°C and 45°C but not at 50°C. Optimum temperatures for oospores production were between 30-35°C. Oospores production happened over a range of pH 5-9 with Optimum values between 6.5-7.5. Oospores produced at -0.13 to -1.65 MPa osmotic potentials with the optimum production rate at -0.27 to -0.47 MPa. Oospores produced at 0.1-1% NaCl with the optimum production rate at 0.3-0.4%.
PDF Abstract XML References Citation
Received: February 18, 2012;
Accepted: April 16, 2012;
Published: June 21, 2012
How to cite this article
Hashem Al-Sheikh and Hani M.A. Abdelzaher, 2012. Materials for Pythium Flora of Saudi Arabia (I) Occurrence, Pathogenicity and Physiology of Reproduction of Pythium aphanidermatum (Edson) Fitzp. Isolated from North and East Regions of Saudi Arabia. Research Journal of Microbiology, 7: 82-100.
URL: https://scialert.net/abstract/?doi=jm.2012.82.100
URL: https://scialert.net/abstract/?doi=jm.2012.82.100
INTRODUCTION
Pythium species belong to oomycetes that grow and thrive in wet environments. They are a distinct group and have unique morphology and physiology. These fungi are characterized by the diversity and their presence all over the world with wide host range (El-Hissy et al., 2001; Abdelzaher, 2009). Pythium commonly occur in cultivated and non-cultivated soils (El-Naghy et al., 2002). Its species occur most abundantly in cultivated soils near the root regions particularly in the superficial soil layers but less commonly in non-cultivated or acid soils. Dry deserts, dry forests and salt media are generally poor in Pythium species (Al-Sheikh and Fathi, 2010; Abdelzaher, 2009; Elnaghy et al., 2010; Moharam, 2010). Their survival in soil occurs as saprophytic mycelial growth or in the form of resistant oospores. Its activity is promoted by high soil moisture as well as by any other soil factors that may give this genus a competitive advantage against other soil microorganisms. It qualifies as a primary and opportunistic invader of plant tissues; it conquests rapidly but can soon be overcome by competitors. In general, Pythium species are very common in rhizosphere soil (Alhussaen et al., 2011; Amein, 2006; Kerkeni et al., 2007; Elnaghy et al., 2010; Moharam, 2010).
Pythium taxonomy has been basically relying on morphological features such as size and shape of sporangia, antheridia or oogonia (Van der Plaats-Niterink, 1981; Dick, 1990; Moustafa et al., 2010; Al-Sheikh and Abdelzaher, 2010a, b). Recently, Molecular characteristics become very important to confirm the identification and classification of fungi (Paul, 2003; Moustafa et al., 2010). The Internal Transcribed Spacer (ITS) region of the rDNA gene sequences has become the useful way in this respect and can be used for identifying or detecting different Pythium spp. (Paul, 2002a, b; Levesque and de Cock, 2004).
In 1990, it was known to have more than 130 species (Paul et al., 2005) and increased thereafter to more than 200 species (Levesque and de Cock, 2004).
Investigation of this genus in Saudi Arabia has begun recently. Very little investigations were done concerning some diseases caused by Pythium spp. (Sunboul, 2001, 2006). IMI (records for geographical unit Arabian Peninsula) reserves an isolate of Pythium aphanidermatum which isolated from Al-Madinah and was associated with Lycopersicon esculentum L. in 1983 (Herb IMI 291421). In addition, some investigators isolated some Pythium species from the infected and rhizosphere of healthy plants grown in Saudi Arabia but they did not provide descriptions and illustrations of the pure cultures of the isolated Pythium spp. and identification based on morphological and molecular criteria (Sunboul, 2001; Molan, 2009).
Studies on Pythium, particularly in the rhizosphere soil of different plants, have been carried out by several researchers all over the world. In Saudi Arabia, very little attention has been paid for isolation and identification of this important fungus. Except for few studies performed in certain mycological laboratories, no comprehensive study has been performed along this line. Therefore, Saudi Arabia is a virgin area for studying Pythium and researches should be done to fill this gap.
In an attempt to fill this gap, this study was undertaken to obtain information about the occurrence of Pythium species in different localities in Saudi Arabia particularly in the rhizosphere zone of some crop plants cultivated in these areas. Pathogenicity of the isolated Pythium species using cucumber was tested, a full description of the isolated species based on morphological and molecular criteria was provided and physiology of zoospores formation and oospores production were done. This investigation is from a sere of work concerning a comprehensive study of Pythium flora in Saudi Arabia.
MATERIALS AND METHODS
Survey points: Collections were made during spring of 2011. The practical work and experiments were done at laboratories of Ah-jouf University, Saudi Arabia, King Faisal University, Al-Hassa, Saudi Arabia and laboratory of Mycology, Department of Botany and Microbiology, College of Science, Minia University, Egypt.
Collection of rhizosphere soil from around roots of 4 crop plants was performed from the following six sites:
| • | Farms at the east of Laquayet (Sakaka) (29°58’56½N, 40°12’10½ E), cultivated with maize |
| • | A farm at the road between Sakaka and Domet Algandal (29°51’56½ N, 39°51’20½ E), cultivated with wheat |
| • | A farm at Sakaka city (29°58’56½ N, 40°12’50½ E), cultivated with potato |
| • | A farm located at 8 km west of Maiquo, Domet algandal (29°51’04½ N, 38°57’02½ E), cultivated with potato |
| • | A farm at Domet Algandal (29°48’58½ N, 39°52’20½ E) cultivated with potato |
| • | A farm at Quara village, Alahsa (25°23’64½ N, 49°38’25½ E), cultivated with tomato |
Collections were made in the spring of 2011.
Fungal isolation: Isolation of Pythium spp., from the rhizosphere soil from the plant roots was performed using the following method: 1- plant root were lifted gently from the soil and shaken by hand to remove large soil particles and then transferred into sterile plastic bags and taken to the laboratory to be used further for analysis on the same day or stored in a refrigerator at 4°C till used. 2- Soil particles were removed from the roots of each plant and placed on the surface of NARM [nystatin (10 mg L-1), ampicillin (250 mg L-1), rifampicin (10 mg L-1) and miconazole (1 mg L-1) in cornmeal agar (CMA)] (Morita and Tojo, 2007; Senda et al., 2009) for isolation of Pythium species, selectively, in 9 cm diam Petri plates at 4 corners in each plate. Four replicates for each plant were made. The emerging hyphal tips were transferred to Water Agar (WA) for further purification from bacterial contamination as follows: distal end of a colony growing on the NARM medium was cut and re-inoculated on 2.5-3% (WA) in a Petri-dish to obtain a colony of about 1 cm diameter. The whole agar medium in the same Petri-dish and incubated until the colony reached before the edge of the dish wall. During this procedure, the non-contaminated mycelia penetrated the agar medium and reached its top. Thin pieces of the agar containing a single hyphal tip of the fungus were taken from the margin of the colony and transferred to Corn Meal Agar (CMA) slants for maintaining the fungus and to CMA plates supplemented with 500 μg mL-1 wheat germ oil to check the formation of sexual structures (Abdelzaher, 2009).
Identification of Pythium spp.
Morphological studies: For purification and morphological identification of isolated pythia, the NARM selective medium was found to be effective in inhibiting the development of bacteria in Pythium cultures whilst not affecting Pythium itself. The position, shape and size of sporangia, the formation of zoospores and the position, shape and size of antheridia, oogonia and oospores were determined in grass blade culture (Senda et al., 2009). Autoclaved grass blades were placed on 2% Water Agar (WA) inoculated with each isolate. After incubation for 2 days at 25°C, colonized grass blades were transferred to autoclaved distilled water and were incubated for 1-14 days at 20°C to follow the fungal development. Morphological identification was done using the keys of Van der Plaats-Niterink (1981) and Dick (1990), as well as the original description of each species isolated.
Molecular studies
DNA extraction: Mycelia were grown in V8 agar medium at 25°C for 7 days or until adequate growth was observed. To extract the total genomic DNA, mycelia from the edge of Pythium colony from a culture plate were suspended in 200 μL of PrepMan Ultra Sample Preparation Reagent (Applied Biosystems, CN, USA) in a 2.0 mL microcentrifuge tube. Samples were vortexed for 10-30 sec and then heated for 10 min at 100°C in dry thermo unit or water bath. Samples were spin for 30 min at 15000 g. Supernatants were transferred into a new microcentrifuge tubes and were ready for (PCR) amplification by the polymerase chain reaction (Senda et al., 2009).
DNA amplification and sequencing: The nuclear rDNA region of the internal transcribed spacer (ITS), including the 5.8S rDNA, was amplified with the universal primers ITS4 (5’ T C C T C C G C T T A T T G A T A T G C 3’) and ITS5 (5’ G G A A G T A A A A G T C G T A A C A A G G 3’). Depending on the experiment, sometimes, primers of ITS1 (5’ T C C G T A G G T G A A C C T G C G G 3’) and ITS2 (5’G C T G C G T T C T T C A T C G A T G C 3’) were used as described and modified by Martin (2000). The amplicons were 700-900 bp long. On the other hand, 563 bp of the cox II gene was amplified in certain Pythia with the primer pair FM66 (5’ TAGGATTTCAAGATCCTGC 3’) and FM58 (5’ CCACAAATTTCACTACATTGA 3’) (Martin, 2000). Amplification of the sequencing template was carried out with DNA Thermal Cycler 2700 (Applied Biosystems) with a cycling profile of pre-PCR at 94°C for 5 min, followed by denaturation at 94°C for I min, I min primer annealing at 55°C for ITS, 52°C for cox II and elongation at 72°C, 2 min for 40 cycles, with a 7 min extension at 72°C after the final cycle. To check the presence of PCR products, 5 μL of the PCR reaction mixture was loaded in 2% L03 (Takara Bio) agarose gel, electrophoresed at 100 V, 20-30 min and stained with ethidium bromide. The sequencing templates were purified with GenElute PCR Clean-up kit (Sigma Chemical Co., St Louis, Missouri, USA) following the manufacturer’s instructions. Sequencing was performed with BigDye Terminator v3.1 Cycle Sequencing Reaction kit (Applied Biosystems) using the same primers in the initial PCR step. After purifying the sequencing reaction mixture through ethanol precipitation it was run on ABI 3100 DNA Sequencer (Applied Biosystems).
Temperature-growth relations: Minimum, optimum and maximum temperatures of each fungus are concluded on cornmeal agar (CMA; 17 g L-1, BD-BBLTM) inoculated with 5 mm dam discs from stock cultures on CMA plates. All plates were incubated at 25°C for 24 h before starting growth determinations. Cardinal temperatures were evaluated by measuring growth at different temperatures (2, 5, 10, 15, 20, 25, 30, 35, 40, 43 and 45 °C).
Pathogenicity test
In agar bottles: A new test was designed in which pathogenicity was evaluated in water agar as a medium for seed germination. One hundred mL of water agar (2%) was poured each in 250 mL Erlenmeyer flasks and then sterilized by autoclaving. Cucumber seeds (Cucumis sativus L.) which proved to be highly susceptible to damping-off disease by Pythium species (Abdelzaher, 2009) were sterilized by surface disinfection using Clorox 2% for 3 min and then washing three times by sterilized distilled water followed by 1 min in 70% ethyl alcohol and finally three times using sterilized distilled water. Seeds were germinated to form radicles and plumules for 2 days at 25°C and selected viable ones were used, thereafter. In each Erlenmeyer flask, three cucumber seeds were planted for damping-off test. Three discs of each fungal isolate taken from actively growing margin of Pythium colonies grown on CMA medium were added to each flask containing cucumber seeds under aseptic conditions. All inoculated flasks were then incubated in a growth cabinet (Precision, United States) at 25°C with 12 h photoperiod (91 μmol m-2 S-1). Damping-off was determined as the difference between seedlings emergence in non-inoculated controls and inoculated one.
In pots: For preparation of inocula, a new method was developed. Five grams of grass blade leaf segments (0.5x1 cm) and 2 g glucose were moistened by adding distilled water (10 mL) each in 250 mL Erlenmeyer flask. After autoclaving at 121°C for 20 min, each flask was inoculated with three disks (7 mm dam) of agar inoculated with growing margins of Pythium spp. obtained from CMA culture medium. The inoculated conical were held at 25°C for 10 days. In pre-emergence damping-off tests, the inoculum concentration of 2.5%, was obtained by mixing thoroughly the 1 g of colonized grass leaf segments in the Erlenmeyer flask with 50 g of oven-dried (70-80°C for 2 days) clay loam soil using a sterilized mortar and pestle. 2.5 gm of this mixture were added to 97.5 g of clay loam soil which had been sterilized by autoclaving at 121°C for 60 min (pH 7.1 after autoclaving) and kept in plastic bag for 2-3 weeks at room temperature with 25% water content prior to use (CL soil). The experiments were carried out in a growth cabinet (Precision, United States) at 25°C with 12 h photoperiod (91 μmol m-2 S-1). Healthy and diseased seedlings were counted at regular intervals until the development of two true leaves in non-inoculated control. Pre-emergence damping-off was determined as difference in emergence between healthy control soil and diseased soil.
Factors affecting zoospores production by six isolates of P. aphanidermatum
Effect of temperature: Grass blades of Echinochloa colonum L. were cut into 3x10 mm pieces and then autoclaved at 121°C for 20 min. Different species of Pythium were inoculated on CMA medium and after the fungal colonies reached about 4 cm diam, the autoclaved Echinochloa colonum leaf blades were laid over each colony in contact with the actively growing margin and incubated at 25°C. After 1 day of incubation the colonized grass blades were transferred to Petri dishes (7 cm diam) containing 10 mL of sterilized distilled water at temperatures of 5, 10, 15, 20, 25, 30, 35 and 40°C to determine the effect of temperature on zoospore production.
Effect of pH: To study the effect of hydrogen ion concentration on zoospore production, the colonized (Echinochloa colonum) leaf blades were transferred to Petri dishes (7 cm diam) containing 10 mL of 0.025 M MES [2-(N-morpholino) ethanesulfonic acid] buffer which had been adjusted to pH values of 4 to 7 with 1N HCl or 1N NaOH. CHES [2(cyclohexylamino) ethanesulfonic acid] buffer was used to make pH values of 8 and 9 with the aid of 1N HCl or NaOH. The solution and buffers were sterilized by filtration.
Effect of osmotic potential: To study the effect of osmotic potential on zoospore production, the colonized (Echinochloa colonum) leaf blades were transferred to Petri dishes (7 cm diam) containing 10 mL of sterilized mannitol solution dissolved in distilled water which is inactive to Pythium, of different osmotic potentials (-0.27, -0.47 and -1.00 MPa). The osmotic potential was determined with a Dew point Microvoltmeter (HR-33T; Wescor, logan, USA). Distilled water was used as test solution.
Effect of salinity: To study the effect of salinity colonized (E. colonum) leaf blades were transferred to Petri-dishes containing 10 mL portions of autoclaved NaCl solutions with different concentrations of 0.1, 0.2, 0.3, 0.4, 0.5, 1, 2 and 3% g.
Factors affecting oospores production by six isolates of P. aphanidrmitium
Effect of temperature: To examine the effect of temperature on oospores production, Pythium species were cultured to produce oospores at different temperatures ranging from 5-45°C for three weeks in 100 mL Erlenmeyer flasks containing 10 mL of clarified V-8 juice medium containing 20% V8 vegetable juice, V/V and 0.25% CaCO3 and was clarified by centrifugation at 13,200 g for 30 min. (V-8) (Moharam, 2010). Oospores suspensions were then obtained by mincing mycelial mats in a blender for 3 min. Because average oospores diameter of P. aphanidermatum was 20 μL, respectively, the resulting suspension was filtered through a sieve (15 μL pore diameter for P. aphanidermatum) in order to produce a suspension of oospores reasonably free of hyphal fragments. The number of mature (vital contents and intact walls) oospores were counted and related to the examined criteria.
Effect of pH: The effect of pH on oospores production was conducted using 2-(N-morpholino) ethanesulfonic acid (MES) buffer (Moharam, 2010). Double strength of the buffer and V-8 juice medium were separately autoclaved at 121°C for 15 min and the cooled buffer was then added to an equal volume of the medium to give a final buffer concentration of 50 mM and adjusted between 4.5 and 9 at 0.5 pH unit intervals using 1N HCl or 1N NaOH. Flasks were incubated at 28°C for three weeks in the dark.
Effect of osmotic potential: In the osmotic potential experiment, mannitol which was found not to be utilized by Pythium as a nutrient (Moharam, 2010) was added to V-8 juice to obtain potentials of -0.13, -0.27, -0.47, -1.00, -1.65 and -3.40 MPa according to Robinson and Stokes’s formula (Moharam, 2010). Flasks were incubated at 28°C for three weeks in the dark the osmotic potential was determined with a Dew point Microvoltmeter (HR-33T; Wescor, logan, USA). Distilled water was used as test solution.
Effect of salinity: In the salinity experiment, NaCl solutions with different concentrations of 0.1, 0.2, 0.3, 0.4, 0.5, 1, 2 and 3% gm was added to V-8 juice according to Robinson and Stokes’s formula (Moharam, 2010). Flasks were incubated at 28°C for three weeks in the dark.
RESULTS
Occurrence of Pythium in rhizosphere soil of the studied plants: Seventy four isolates belonging to P. aphanidermatum were isolated from the rhizosphere soil of 4 crop plants (maize, potato, tomato and wheat) cultivated in 6 farms located as described previously. The 15, 13, 13, 12, 11, 10 isolates of this fungus were obtained from the rhizosphere soil of maize (sit 1), wheat (site 2), potato (sit 3), potato (site 4), potato (site 5) and tomato (site 6), respectively.
Results also show that P. aphanidermatum was the only species in the rhizosphere soil of the studied plants. P. aphanidermatum, representing 100% of the total Pythium isolations were obtained from the rhizosphere soil of the studied plants.
Description of Pythium aphanidermatum: One isolate representing (isolates 1-6) the isolates obtained from each site was choused to be studied in the physiology section of this study.
Morphological characterization of P. aphanidermatum were done using a single isolate (isolate 1) which characterized as follows.
Pythium aphanidermatum (Edson) Fitzp.(MU0001) (Plate 1): Colonies on cornmeal agar have a cottony aerial mycelium while on potato-carrot agar they have some loose aerial mycelium without a special pattern (Plate 2a-c). Main hyphae are up to 10 μm wide. Zoosporangia consist of terminal complexes of swollen hyphal branches of varying length and up to 22 μm wide. Zoospores are formed at 15-30°C. Encysted zoospores have a diameter of 12 μm. Oogonia are terminal, globose, smooth and of (20-) 22-25(-26) μm (av. 24 μm) diam.
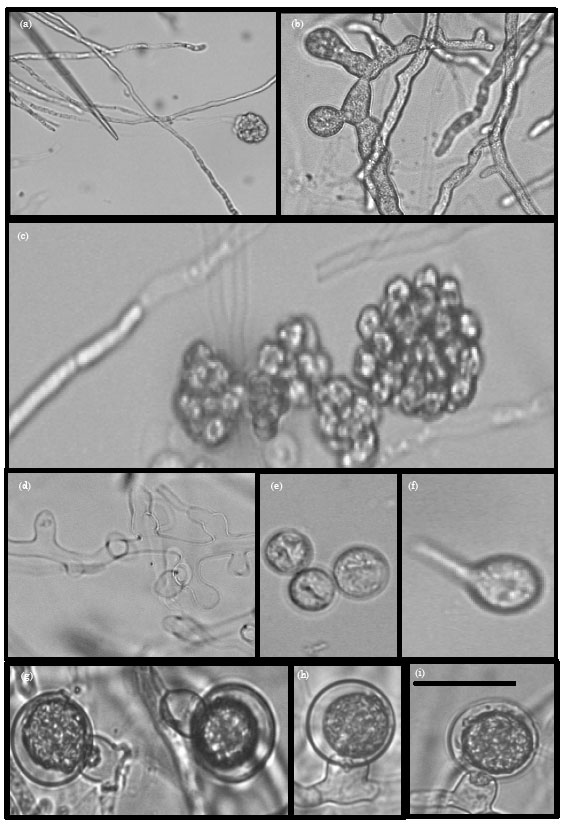 | |
| Plate 1(a-i): | :Morphology of Pythium aphanidermatum. (a) A toruloid zoosporangium with vesicles, (b) A young toruloid zoosporangium, (d) An empty toruloid zoosporangium, (c, e-f) Encysted and germinating zoospores, (g-i) Young oogonia and intercalary and Bar on photo (i) is equal 20 μm |
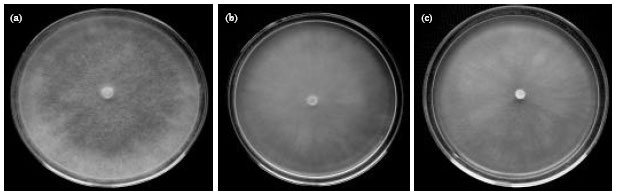 | |
| Plate 2(a-c): | :Growth habits of P. aphanidermatum (isolate I, MU0001) on Biolife-PDA (a) PCA (b) Difco-CMA and (c) at 25°C for 5 days |
 | |
| Fig. 1: | Nuclear rDNA region of the internal transcribed spacer (ITS), including the 5.8S rDNA, of P. aphanidermatum, All of the six isolates had the same sequence |
Antheridia are mostly intercalary, sometimes terminal, broadly sac-shaped, 11-15 μm long and 9-15 μm wide, 1(-2) per oogonium and monoclinous or diclinous. Oospores are aplerotic, (19-)20-23 μm (av. 21 μm) in dam and their walls are 1-2 μm thick. Cardinal temperatures: minimum 10°C, optimum 35 °C, maximum 42 °C. Daily growth rate on Bacto-CMA was 19 mm.
Molecular characterization of P. aphanidermatum was done for isolates (1-6): Nuclear rDNA region of the Internal Transcribed Spacer (ITS), including the 5.8S rDNA, of P. aphanidermatum is shown in Fig. 1.
Cardinal temperatures of P. aphanidermatum: As shown in Table 1 P. aphanidermatum showed temperature maxima higher than 45°C. The optimum temperature for all isolates lied between (30 and 35°C). The minimum temperature supporting mycelial growth of P. aphanidermatum was around 10°C.
| Table 1: | Linear growth of six isolates of Pythium aphanidermatum isolated from rhizosphere soil of tested plants cultivated in Al-Jouf and Al-Hassa at various temperatures |
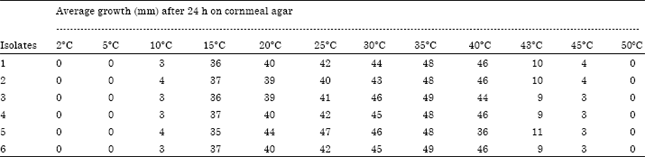 | |
| Growth is the mean of five replicates and no significant difference in diameter within replicates of each measurement was observed | |
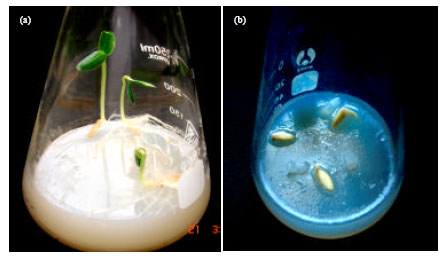 | |
| Plate 3(a-b): | Erlenmeyer flasks containing 2% water agar infested with Pythium aphanidermatum (isolate I, MU0001), (a) No Pythium inocula in the control flask (b) Pre-emergence damping-off of cucumber seeds |
Pathogenicity tests
In agar bottles: Pathogenicity (pre-emergence damping-off) of the six isolates of P. aphanidermatum was tested on cucumber seeds. All of the tested isolates of P. aphanidermatum were highly pathogenic to cucumber seeds causing 100% damping-off (Plate 3).
In pot experiment: Pathogenicity tests (pre-emergence damping-off) were performed also in pots using 2.5% inoculum concentration of the tested P. aphanidermatum added to soil and incubated for 2-3 weeks before cucumber seeds were cultivated. All of the tested isolates of P. aphanidermatum proved to be highly pathogenic to cucumber seeds causing 100% damping-off (Plate 4).
Factors affecting zoospores production of six isolates of P. aphanidermatum.: Influence of temperature on zoospores production, Fig. 2 shows six isolate of P. aphanidermatum started to produce zoospores after 72 h incubation at 20, 25, 30, 35 and 40°C. At lower temperature (15°C), the fungus produced zoospores after 72 h, till the end of the experimental period whereas at high temperatures production declined drastically.
 | |
| Plate 4(a-b): | Clay sandy soil in plastic pots infested with Pythium aphanidermatum (isolate I, MU0001), (a) No Pythium inocula in the control pot, (b) Pre-emergence damping-off of cucumber seeds |
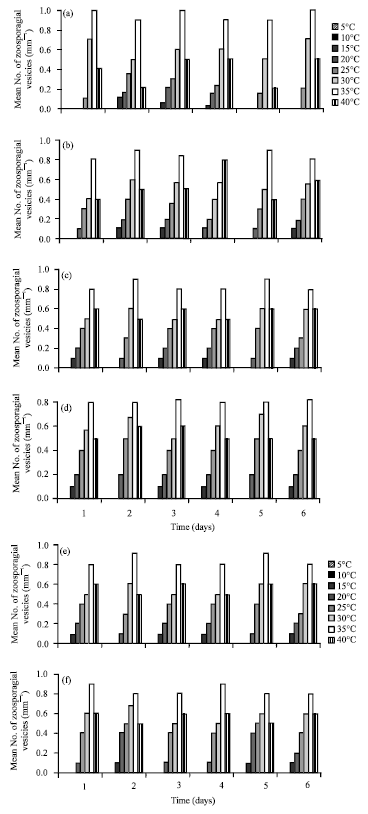 | |
| Fig. 2(a-f): | Effect of temperature on zoospore production by six isolates of P. aphanidermatum, (a) Isolate 1, (b) Isolate 2, (c) Isolate 3, (d) Isolate 4, (e) Isolate 5 and (f) Isolate 6 |
 | |
| Fig. 3(a-f): | Effect of hydrogen ion concentration on zoospore production by six isolates of P. aphanidermatum, (a) Isolate 1, (b) Isolate 2, (c) Isolate 3, (d) Isolate 4, (e) Isolate 5 and (f) Isolate 6 |
Influence of hydrogen ion concentration on zoospores production: All isolates produce zoospores within the pH range 5-8. Optimum pH for zoospore production was 7 for all fungi used. There was no zoospore production at pH 4 and 5 for all the tested pythia (Fig. 3).
Influence of osmotic potential on zoospores production: Figure 4 represents the effect of osmotic potential on zoospore production by the species tested. Distilled water (zero osmotic potential) was favorable for good production by tested fungi. In distilled water containing mannitol was added to adjust osmotic potential between -0.13 to -1.00 MPa. All isolates were able to produce zoospores between -0.13 to -0.27 MPa but not above this range. The highest zoospore production was found at -0.27 MPa for all species tested.
Influence of salinity on zoospores production: Figure 5 shows that production of zoospores by all of the isolates were inhibited by 0.5% NaCl. The optimum concentration for zoospore production were between 0.1 and 0.2% g NaCl.
Factors affecting oospore production
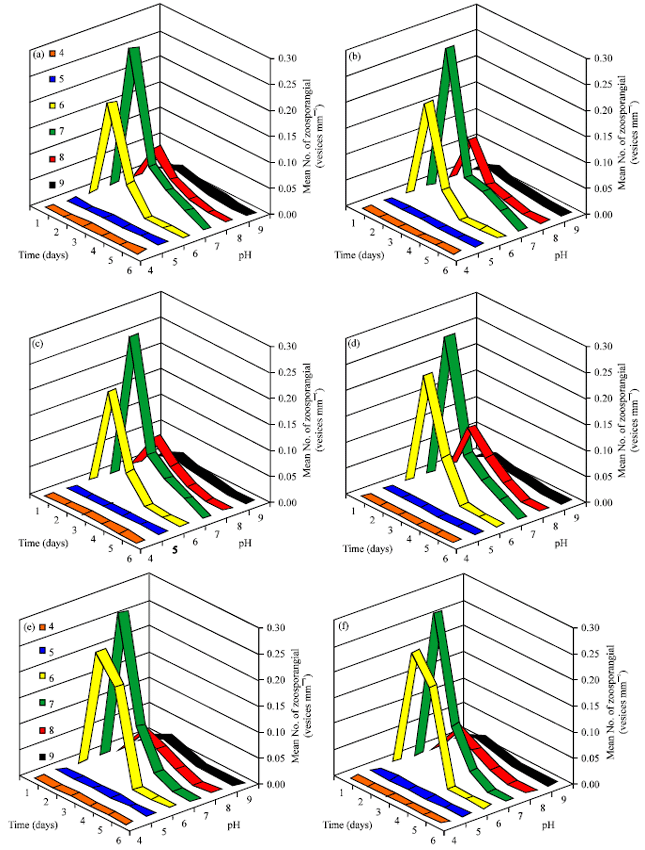
Influence of temperature on oospores production: As shown in Fig. 6, oospores produced by six isolates of P. aphanidermatum over a temperature range between 10 and 45°C but not at 5°C. Optimum temperatures were between 30-35°C for oospores production by the six isolates of P. aphanidermatum.
Influence of hydrogen ion concentration on oospores production: As shown in Fig. 7, oospores production occurred over a range of pH 5-9. Optimum pH values were noticed between 6.5-7.5 for all species tested.
Influence of osmotic potential on oospores production: The effect of osmotic potential on oospores production at 25°C is indicated in Fig. 8. The isolates showed similar responses. Oospores produced at -0.13 to -1.65 MPa with the optimum production rate at -0.27 to -0.47 MPa.
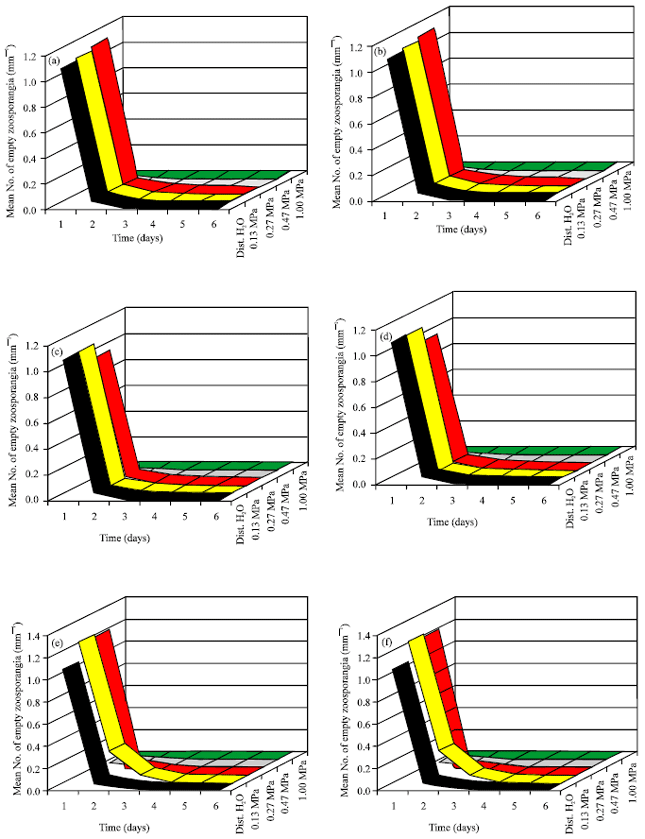 | |
| Fig. 4(a-f): | Effect of osmotic potential (MPa) on zoospore production by six isolates of P. aphanidermatum at 25°C, (a) Isolate 1, (b) Isolate 2, (c) Isolate 3, (d) Isolate 4, (e) Isolate 5 and (f) Isolate 6 |
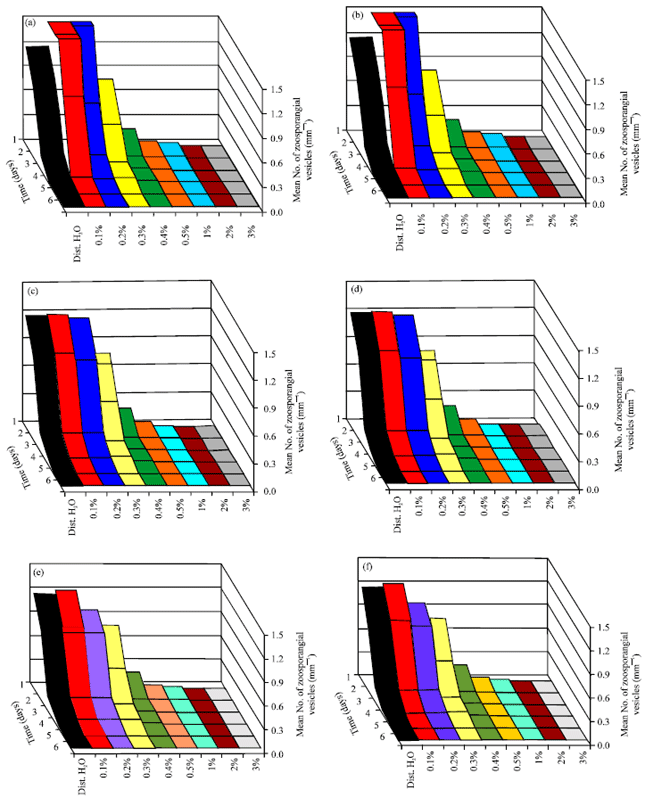 | |
| Fig. 5(a-f): | Effect of salinity (NaCl%) on zoospore production by six isolates of P. aphanidermatum at 25°C, (a) Ioslate 1, (b) Ioslate 2, (c) Ioslate 3, (d) Ioslate 4, (e) Ioslate 5 and (f) Ioslate 6 |
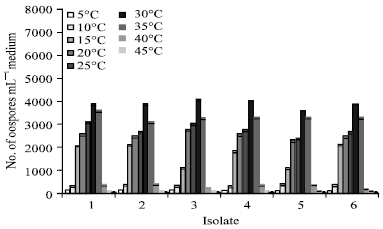 | |
| Fig. 6: | Effect of temperature on oospores production by six isolates of P. aphanidermatum. grown on V-8 juice medium for 21 days in the dark |
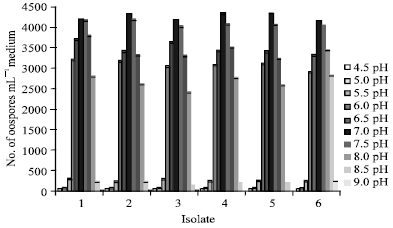 | |
| Fig. 7: | Effect of hydrogen ion concentration on oospores production by six isolates of P. aphanidermatum grown on V-8 juice medium for 21 days at 25°C in the dark |
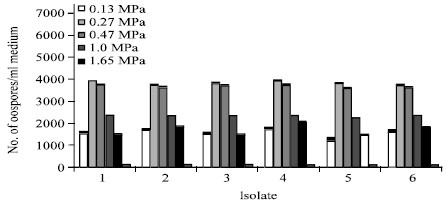 | |
| Fig. 8: | Effect of osmotic potential on oospores production by six isolates of P. aphanidermatum grown on V-8 juice medium for 21 days at 25°Cin the dark |
Influence of salinity on oospores production: The effect of salinity on oospores production at 25°C is indicated in Fig. 9. Oospores produced at 0.1-1% NaCl with the optimum production rate at 0.3-0.4% NaCl. No oospores produced over the value of 1% NaCl.
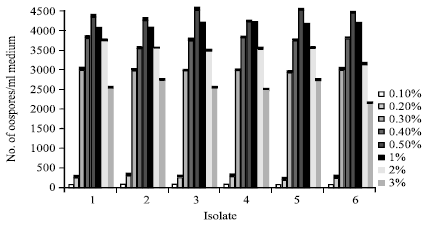 | |
| Fig. 9: | Effect of salinity NaCl on oospores production by six isolates of P. aphanidermatum grown on V-8 juice medium for 21 days at 25°C in the dark |
DISCUSSION
P. aphanidermatum are typical plant parasites of plants in warm regions (Al-Sheikh and Abdelzaher, 2010a, b; Moharam, 2010). The results show that P. aphanidermatum was the only species in the rhizosphere soil of the different plans studies. The ITS region of the nuclear ribosomal DNA of our isolate of P. aphanidermatum is comprised of 777 bases and a BLAST search showed complete resemblance of 100% similarity (Genbank accession number (AB274404.1). Subsequently, based on morphological (Plate 1) and molecular criteria, this study confirms the identification of P. aphanidermatum for the first time in Saudi Arabia using molecular identification.
From previous studies, rhizosphere soil of various plants could be considered as favorable environment for the occurrence of Pythium spp., indicating that the plant root exudates enhance the growth and proliferation of Pythium spp. In general, Pythium may be considered as one of the main components of the rhizosphere soil mycoflora in warm and hot places but they were almost absent in desert soil (Nasser, 2003; Al-Sheikh, 2010; Al-Sheikh and Abdelzaher, 2010a, b; El-Naghy et al., 2002). This may be due to scarce of humidity of the soil as well as absence of organic matter which support the saprophytic ability of these pythia. Results here confirm occurrence of P. aphanidermatum in the rhizosphere soil of maize, potato, tomato and wheat, cultivated in five farms in Al-Jouf and a farm in Al-Hassa, Saudi Arabia. These two areas of Saudi Arabia are characterized by their arid and very hot condition in summer. Temperatures are often exceeded 50°C in summer in the shade. Isolates of P. aphanidermatum show ability to grow over 45°C which differ from cardinal temperatures of other isolates in other countries (Al-Sheikh, 2010). Therefore, isolates of P. aphanidermatum obtained from this study can tolerate and adapt to grow and establish in Saudi Arabian weather.
The pathogenicity of P. aphanidermatum was previously proved (Moustafa et al., 2010; Al-Sheikh, 2010). It can cause root rot and damping-off, stalk and rhizome rot, soft rot, fruit rot or cottony blight of Abelmoschus esculentus L., Basella alba L., Carica papaya L., Catharanthus roseus L.., Carthamus creticus L., Chrysanthemum indicum L., Citrus sp. L., conifers, corn, a number of other crops, crucifers, cucurbitaceae, leguminosae, Fragaria sp. L., Gossypium sp. L., grasses, Lactuca sativa L., Linum usitatissimum L., peppers, poinsettia, potato, sugar beet, sugar cane, Talinum fruticosum L., Tephrosia sp. Pers, tobacco and tomato (Van der Plaats-Niterink, 1981; Sunboul, 2006).
Results here revealed that the six isolated of P. aphanidermatum in this study similar in their virulence. P. aphanidermatum was highly pathogenic to cucumber germinating seeds causing 100% pre-emergence damping-off. Therefore, occurrence of P. aphanidermatum in rhizosphere soil of maize, potato, tomato and wheat cultivated if six farms in Saudi Arabia represent a very dangerous factor for causing diseases to these important crop plants. This virulent species of Pythium can cause diseases especially when environmental conditions favor disease prevalence.
Subsequently, studying some main factors (temperature, pH, osmotic potential and salinity) affecting asexual and sexual reproduction of P. aphanidermatum is important in order to shed light on life cycle of this important fungus and its manipulation in the soil under variable conditions. The asexual reproduction of Pythium species is effected by swimming zoospores liberated from a zoosporangium which is undifferentiated from the vegetative thallus in the lower forms but cut off as an enlarged, more or less spherical cell in the higher. As the genus invaded the land, the zoospores were gradually suppressed and in the higher forms the sporangium germinates as a whole by putting out a germ filament which grows into a new plant (Moustafa et al., 2009).
One of the important roles of zoospores is to serve for the dissemination of fungi. P. aphanidermatum produced zoospores at 15°C indicating optimum temperatures for zoospore formation is not necessarily coinciding with that of mycelial growth. while P. aphanidermatum could not produce zoospores at 50°C (Fig. 3). Acidic solutions (pH 4, 5) proved unsuitable for zoospore production of all fungi tested (Fig. 3). Results here (Fig. 5) confirms that Moderate osmotic potential appeared to favor zoospore production. This phenomenon is correlated with the influence of osmotic strength on the formation of the vesicles since at higher concentration levels, although fungi could produce vesicles, these failed to produce zoospores. Moustafa et al. (2010), in their study on the role of quality of irrigation water in dissemination of Pythium species to agricultural fields, concluded that no isolate had been detected in heavily polluted waters and had seldom been found in partially polluted places. For the above reason, it is speculated that highly polluted soil and water with high osmotic potential act as a barrier against zoospore production by the six tested Pythium isolates (Fig. 4). A concentration of 0.2% NaCl was found suitable for zoospore production. Above this level a drastic decrease in this process was recorded (Fig. 5). This phenomenon can be explained on the basis of the effect of salinity on vesicles formation. Pythium vesicle has a thin membrane sensitive to the surrounding medium. This permeable membrane needs a special NaCl concentration for complete differentiation. A level of 0.2% NaCl matched the ideal concentration and appeared to favor zoospore production. It can be concluded that soil quality influence mycelial and zoospore production by Pythium sp. highly acidic, heavily polluted and saline soil water extracts do not support mycelial and zoospore formation by these fungi. However, a generalization cannot be made and variation can occur throughout pythia.
In case of sexual reproduction, studying oospores production of some isolated pythia are considered important as survival structures. These oospores can remain viable after 8 months to 12 years in the soil (Elnaghy et al., 2010). When the soil conditions become favorable, oospores germinate to produce huge amount of disease inducing elements which spread from place to place causing infestation. The production of oospores is dependent on several factors including temperature, hydrogen ion concentration, osmotic potential and salinity. Results here indicate that the most favorable temperatures for oospore production were 30-35°C by the tested fungi. It could be postulated that the very cold winter and very hot summer retard oospore production of such fungi (Elnaghy et al., 2010). This can be explained that the high temperature in the daytime in summer in the desert lead to the death of vegetative structures of the fungus and retard oospore production but this could be accelerated during the night. For these reasons, cultivation in Al-Jouf and Al-Hassa areas of Saudi Arabia in spring and early summer encourage disease prevalence by P. aphanidermatum but during summer plants grown under trees in the shade could be infected. Oospore produced over a broad pH range with an optimum around neutral. Previous studies revealed that Pythium spp. non-tolerant to acid soils (Moustafa et al., 2009). In this study, no oospores have been produced below pH 5. It could be expected that acidic soil retard oospores production. Pythium species have been absent from heavily polluted places and seldom found in partially polluted locations (Elnaghy et al., 2010). It would be suggested that, high-polluted locations with osmotic potentials above -1.65 MPa represent inhibitive media toward oospore production. Oospores produced at 0.1-1% NaCl with the optimum production rate at 0.3-0.4% NaCl. No oospores produced over the value of 1% NaCl. For this reason, saline soil represents a barrier to oospore production of P. aphanidermatum.
Results here are compatible with the results of many previous researches (Cliquet and Tirilly, 2002; Elnaghy et al., 2010). Oospores which represent the overwintering stage of P. aphanidermatum seem to exhibit the same behavior in their presence in any place and under any circumstance.
Further studies on Pythium species in soil of Saudi Arabia should be done in order to establish the science of this important fungus.
ACKNOWLEDGMENT
The authors are thankful to King Faisal University for partially supporting this project (small grant No. 1210001).
REFERENCES
- Alhussaen, K., E.I. Hussein, K.M. Al-Batayneh, M. Al-Khatib and W. Al Khateeb et al., 2011. Identification and controlling Pythium sp. infecting tomato seedlings cultivated in Jordan Valley using garlic extract. Asian J. Plant Pathol., 5: 84-92.
CrossRefDirect Link - Al-Sheikh, H. and H.M.A. Abdelzaher, 2010. Differentiation between two isolates of Pythium ultimum var. ultimum isolated from diseased plants in two different continents. J. Biol. Sci., 10: 306-315.
CrossRefDirect Link - Al-Sheikh, H. and H.M.A. Abdelzaher, 2010. Isolation of Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii from agricultural soil and their biological activity against Pythium spinosum, the damping-off organism of soybean. J. Biol. Sci., 10: 178-189.
CrossRefDirect Link - Al-Sheikh, H. and A.A. Fathi, 2010. Ecological studies on lake Al-Asfar (Al-Hassa, Saudi Arabia) with special references to the sediment. Res. J. Environ. Sci., 4: 13-22.
CrossRefDirect Link - Amein, T., 2006. Soil-borne pathogens infecting sugar beet in Southern Sweden. Plant Pathol. J., 5: 356-361.
CrossRefDirect Link - Cliquet, S. and Y. Tirilly, 2002. Development of a defined medium for Pythium oligandrum oospore production. Biocontrol Sci. Technol., 12: 455-467.
CrossRefDirect Link - El-Hissy, F.T., M.S.M. Nassar, A.M. Khallil and F.F. Abdel-Motaal, 2001. Aquatic fungi recovered from water and submerged mud polluted with industrial effluents. J. Biol. Sci., 1: 854-858.
CrossRefDirect Link - El-Naghy, M.A., E.M. Fadl-Allah, H.M.A. Abdelzaher and S.A. Moharam, 2002. Rhizosphere pythia of some plants grown in Egypt. Bull. Fac. Sci. Assiut Univ., 31: 149-157.
Direct Link - Kerkeni, A., M. Daami-Remadi, N. Tarchoun and M.B. Khedher, 2007. In vitro and in vivo suppression of Pythium ultimum the causal agent of the cucumber damping-off by some compost fungi. Asian J. Agric. Res., 1: 50-58.
CrossRefDirect Link - Levesque, C.A. and W.A. de Cock, 2004. Molecular phylogeny and taxonomy of the genus Pythium. Mycol. Res., 108: 1363-1383.
PubMedDirect Link - Martin, F.N., 2000. Phylogenetic relationships among some Pythium species inferred from sequence analysis of the mitochondrially encoded cytochrome oxidase II gene. Mycologia, 92: 711-727.
Direct Link - Molan, Y.Y., 2009. Detection of presumptive mycoparasites in soil placed on host-colonized agar plates in Riyadh Region, Saudi Arabia. Asian J. Plant Pathol., 3: 22-26.
CrossRefDirect Link - Morita, Y. and M. Tojo, 2007. Modifications of PARP medium using fluazinam, miconazole and nystatin for detection of Pythium spp. in soil. Plant Dis., 91: 1591-1599.
CrossRefDirect Link - Nasser, L.A., 2003. Distribution of zoosporic and terrestrial fungi in accumulated rainfall water in Abha, South Western region, Saudi Arabia. J. Biol. Sci., 3: 843-853.
CrossRefDirect Link - Paul, B., 2002. ITS1 region of the rDNA of Pythium proliferatum, a new species: Its taxonomy and its comparison with related species. FEMS Microbiol. Lett., 206: 191-196.
Direct Link - Paul, B., 2002. ITS region of Pythium canariense sp. Nov, its morphology and its interaction with Botrytis cinerea. EMS Microbiol. Lett., 208: 135-141.
Direct Link - Paul, B., 2003. Pythium carbonicum, a new species isolated from a spoil heap in northern France, the ITS region, taxonomy and comparison with related species. FEMS Microbiol. Lett., 219: 269-274.
Direct Link - Paul, B., K. Bala, S. Gognies and A. Belarbi, 2005. Morphological and molecular taxonomy of Pythium longisporangium sp. Nov. isolated from the Burgundian region of France. FEMS Microbiol. Lett., 246: 207-212.
CrossRef - Senda, M., K. Kageyama, H. Suga and C.A. Levesque, 2009. Two new species of pythium, P. senticosum and P. takayamanum, isolated from cool-temperate forest soil in Japan. Mycologia, 101: 439-448.
PubMed - Sunboul, Y.H., 2006. Solarizaton of commercial peat in transparent polyethylene bags and its effect on survival of some plant pathogenic fungi. JKAU Sci., 18: 1-11.
Direct Link - Al-Sheikh, H., 2010. Two pathogenic species of Pythium: P. aphanidermatum and P. diclinum from a wheat field. Saudi J. Biol. Sci., 17: 347-352.
CrossRefPubMedDirect Link