
M. Gokuldas
Division of Insect Biochemistry and Physiology, Department of Zoology, University of Calicut, Kerala, 673635, India
A.P. Ajaykumar
Division of Insect Biochemistry and Physiology, Department of Zoology, University of Calicut, Kerala, 673635, India
D. Umadevi
Division of Insect Biochemistry and Physiology, Department of Zoology, University of Calicut, Kerala, 673635, India
V.S. Binitha
Division of Insect Biochemistry and Physiology, Department of Zoology, University of Calicut, Kerala, 673635, India
K.M. Ismail
Division of Insect Biochemistry and Physiology, Department of Zoology, University of Calicut, Kerala, 673635, India
K.A. Rasheed
Division of Insect Biochemistry and Physiology, Department of Zoology, University of Calicut, Kerala, 673635, India
Journal of Entomology
Year: 2013 | Volume: 10 | Issue: 2 | Page No.: 95-102







ABSTRACT
Small neuropeptides of the adipokinetic hormone/red pigment concentrating hormone family (AKH/RPCH), is one of the important groups of peptides that regulate physiological homeostasis in insects. The adipokinetic neuropeptide of Hieroglyphus banian (Order: Orthoptera; Family: Acrididae) was isolated from corpora cardiaca by HPLC; the chromatographic fractions were examined for adipokinetic activity in the plant bug, Iphita limbata in vivo. Two fractions that peaked at retention times of 9.94 and 11.81 min, respectively were found to be significantly active in mobilization of lipids. One of the biologically active peaks (9.94 min) had identical retention time as that of synthetic Locusta migratoria-Adipokinetic Hormone-I (Locmi-AKH-I). These analyses confirmed the presence of Locmi-AKH-I in the corpora cardiaca of H. banian. This data can be additional information to aid in the construction of phylogenetic trees of the insect order Orthoptera, by means of computer programme and protein parsimony algorithms.
PDF Abstract XML References Citation
Received: September 19, 2012;
Accepted: December 24, 2012;
Published: April 27, 2013
How to cite this article
M. Gokuldas, A.P. Ajaykumar, D. Umadevi, V.S. Binitha, K.M. Ismail and K.A. Rasheed, 2013. Identification of an AKH/RPCH Family Peptide in the Rice Grasshopper, Hieroglyphus banian (Acrididae: Orthoptera). Journal of Entomology, 10: 95-102.
DOI: 10.3923/je.2013.95.102
URL: https://scialert.net/abstract/?doi=je.2013.95.102
DOI: 10.3923/je.2013.95.102
URL: https://scialert.net/abstract/?doi=je.2013.95.102
INTRODUCTION
The vertebrate hypothalamo/hypophyseal system is analogous to neurosecretory X-organ/sinus gland complex in the eyestalks of crustaceans and the intrinsic neurosecretory cells of the corpora cardiaca of insects. The hormones of X-organ-sinus gland complex are known to regulate reproduction, metabolism, osmoregulation, chromatic adaptation and growth (Shuranova et al., 2006; Velmurugan et al., 2008). Similarly, the Adipokinetic hormones (AKHs) in insects act on the fat body to mobilize stored lipids and carbohydrates, activate glycogen phosphorylase, accumulate cAMP (Goldsworthy, 1983) and inhibit the synthesis of lipids (Gokuldas et al., 1988; Lorenz, 2001; Fabold et al., 2010), proteins (Carlisle and Loughton, 1979; Kodrik, 2008) and RNA (Kodrik and Goldsworthy, 1995). Common characteristics of the adipokinetic hormone/red pigment concentrating hormone (AKH/RPCH) family of peptides include, a chain length of 8-10 amino acids; the N-terminus blocked by pyroglutamic acid (pGlu); the C-terminus blocked by a carboxamide; amino acids at positions 8 and 9 (when present) are tryptophan and glycine; most of the peptides are uncharged. There are at least two aromatic acids, at position 4 mostly phenylalanine (but sometimes tyrosine) and at position 8 tryptophan and a few peptides have a third aromatic amino acid either at position 2 (Tyr or Phe) or at position 7 (Trp) (Kodrik, 2008). The actions of AKHs are however, broader than what their name implies. The peptides have been reported from most of the insect orders (Kodrik, 2008). The importance of body fat regulation is emphasized by dysfunctions resulting in obesity and lipodystrophy in humans. Packaging of storage fat in intracellular lipid droplets and various molecules and mechanisms guiding storage fat mobilisation, are conserved between mammals and insects (Gronke et al., 2007). New approaches in molecular biology should be made to exploit the fat mobilizing effect of AKH in order to target this lipid mobilizing hormone effectively for therapeutic purposes for controlling obesity. Such an indication was given by the work done by Schacter and Schacter (2005).
The major objective of the present investigation was to elucidate the primary structures of adipokinetic neuropeptides of H. banian. It is a sporadic pest of paddy in India causing serious damage to paddy cultivation. Knowledge of peptides has been thought to be useful in developing non-toxic, biotechnology-based tools for pest management. The primary structures of AKH/RPCH peptides of the insect were elucidated by HPLC analysis, hyperlipaemic bioassay of the fractions separated on HPLC and comparison of HPLC profiles of synthetic AKHs with that of native peptides.
MATERIALS AND METHODS
Experimental insects: The rice grasshopper, H. banian were collected locally from the paddy fields of Kannur and Malappuram districts of Kerala, India, during the months of August 2009-March 2010). After collection they were kept in plastic containers with perforated lids. In the laboratory, they were immediately subjected to hormone extraction. Adult female plant bug, Iphita limbata were collected locally from around the Calicut University campus. A stock culture of insect, maintained in the laboratory on a diet of sprouted green gram (Phaseolus aureus) and ripe banana at a room temperature of 27±2°C and 25% RH, were used for bioassay experiments.
Preparation of hormone extract: Corpora cardiaca collected from both sexes of adult grasshoppers were used as the sources of the hormone. Heads from the insects were removed and placed with dorsal side up and mouth parts away from us, were cut longitudinally between the eyes into two unequal halves. The smaller part was discarded and from the larger part, corpora cardiaca-allata complexes (retrocerebral complexes) were removed with the help of a pair of fine forceps under a stereo zoom binocular microscope. The tissues were immediately put into ice cold 80% methanol (HPLC grade) and stored at -4°C until extraction. Tissues were sonicated for 1 min on ice with an ultrasonicator (Sonics and Materials, USA). The extracts were centrifuged at 4°C and 10,000 rpm for 10 min (Plastocraft, India). The supernatants were collected into an Eppendorf tube and vacuum dried (Savant, USA). The dried supernatants were stored at -4°C until HPLC separations and bioassay.
Hyperlipaemic in vivo bioassay: Since, experimental insect, H. banian was not readily available for bioassay, I. limbata, a phytophagus bug which has been proved to be an ideal test animal for hyperlipaemic bioassay, have been used here for regular bioassay (crude retrocerebral extract of H. banian as well as fractions collected from HPLC). The dried HPLC fractions (1-20 min) and retrocerebral extracts were redissolved in insect saline; 5 μL of these fractions were injected using a 10 μL Hamilton syringe into the experimental insect through the laterodorsal intersegmental membrane. The injected samples of crude retrocerebral extract (5 μL) contained different concentrations (1, 2, 3, 4, 5, 6 and 7 gland pair equivalent (gpe)). Haemolymph samples were taken at pretreatment and 60 min after the injection. The lipid levels in the samples were determined by spectrophotometric method (Frings et al., 1972) (n = 6), using UV-vis spectrophotometer (Shimadzu) at 540 nm.
HPLC analysis: The dried extract was redissolved in 20 μL of 80% methanol (HPLC grade). The extract was filtered using a sample filtration unit with 0.45 μm (Millipore) filter. The samples (20 μL) were directly injected into the instrument by a microsyringe (Hamilton). HPLC separations were carried out using Shimadzu system (SPD M 10 AVP, LC 10 ATVP, LC 10 ATVP) with a reversed phase column (C18) 250 mm long, 4.6 mm i.d. The separation was done in a binary gradient from 43-53% solvent B in 20 min with a flow rate of 1 mL min-1. Trifluoroacetic acid (TFA) 0.01% in water (HPLC grade) was used as solvent A, solvent B was 60% acetonitrile in solvent A. The eluants were monitored at 210 nm. One minute fractions (1-20 min) were collected manually, dried by vacuum concentrator and were used for testing their hyperlipaemic activity.
The presence of peptides similar to an already known AKH peptide, Locmi-AKH-I (GenScript Corp., USA), in the retrocerebral extracts of H. banian was investigated by analysing the elution pattern on HPLC. The synthetic peptide, Locmi-AKH-I (100 pmol) was injected into the HPLC instrument maintained in the same set up as before for the retrocerebral extracts of H. banian. The HPLC profiles of the peptides were overlaid with that of the retrocerebral extract of H. banian for comparison.
Statistical analysis and data presentation: Values obtained from hyperlipaemic bioassay experiments were expressed as mean±standard error values as well as percentage difference of the experimental over controls (E/C%). The paired t-test for the adipokinetic responses were performed with SPSS Software (version 10). The graphical representation of change in lipid mobilization (E/C%) was plotted by Microsoft Excel programme.
RESULTS AND DISCUSSION
The hyperlipaemic effects of the extracts of H. banian were determined by the change in the lipid content after the injection of extract. The result of the extract is summarized in Table 1. The retrocerebral extract with a concentration of 5 gpe/5 μL induced maximal hyperlipaemic response, 45%. A gradual decrease in lipid release was noticed with the peptide above 5 gpe. The hyperlipaemic effects induced by other concentrations were 9 (1 gpe), 19 (2 gpe), 28 (3 gpe), 20 (4 gpe) and 16 (6 gpe) per cent, respectively. Earlier studies demonstrated significant hyperlipaemic response by the fat body of I. limbata injected with corpora cardiaca (CC) extracts of Spodoptera mauritia (Kumari and Gokaldas, 2001), I. limbata (Rasheed and Gokuldas, 2002; Ajaykumar and Gokuldas, 2009, 2011a) Oryctes rhinoceros (Ajaykumar and Gokuldas, 2011b) and O. nitidula (unpublished data).
| Table 1: | Amount of lipid released into the haemolymph of I. limbata injected with the retrocerebral extracts of H. banian |
 | |
| Values are Mean±SEM (n = 6), The haemolymph lipids were measured before (control) and 60 min after (experiment) injection of retrocerebral extracts of H. banian, *Indicates p<0.05, E/C: Percent increase of experiment over control | |
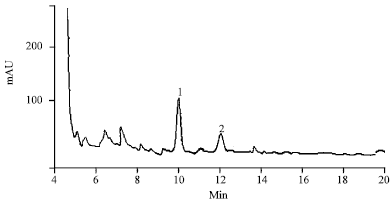 | |
| Fig. 1: | The HPLC profile of extracts of retrocerebral complexes of H. banian. The analysis was carried out on a C18 Hibar column. The extract was run with a gradient of 43-53% B in 20 min, 1: 0.01% trifluoroacetic acid in water (A), 2: 60% acetonitrile in solvent A, The eluants were monitored at 210 nm |
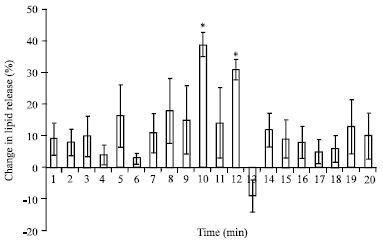 | |
| Fig. 2: | Hyperlipaemic activity of HPLC fraction. The change in total haemolymph lipid is represented as E/C%, *p<0.05 |
Since regulation of intermediary metabolism in grasshoppers appears to be very sophisticated (Siegert et al., 2000), it is suggested that there may be other novel AKH peptides than those already detected that are helpful for mobilizing energy reserves (lipids and carbohydrates) and this may be true with H. banian. It will be worth investigating to find out the novel peptides whose biological activities have been confirmed but the structure has not been able to be figured out.
Analysis of the extract of retrocerebral complexes of H. banian using HPLC showed two prominent absorption peaks with retention times 9.94 and 11.81 min, respectively monitored at 210 nm (Fig. 1). The hyperlipaemic effects of materials in the fractions were tested by injecting the reconstituted fractions into female I. limbata. The materials in the fractions 10 and 12 min showed (Fig. 2) significant adipokinetic activities with increase of lipids by 39 and 31% over the controls, respectively (p<0.05), none of the other fractions were active at statistically significant levels. Locmi-AKH-1 was eluted at the same retention time as that of the material having the retention time 9.94 min in the retrocerebral extract of H. banian.
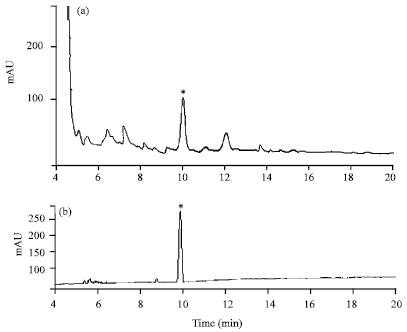 | |
| Fig. 3(a-b): | The HPLC profile of extracts of corpora cardiaca of H. banian (a) Compared with synthetic Locmi-AKHI-I and (b) Monitored at 210 nm, The samples were run in a gradient of 43-53% B in 20 min (solvent A: 0.01% of trifluoroacetic acid in water, solvent B: 60% acetonitrile in solvent A, Eluants were monitored at 210 nm, *Indicates peaks of interest |
| Table 2: | The insects of Acrididae sub-families which possess the adipokinetic hormones, Locmi-AKH-I |
 | |
The materials in the fraction eluted at 11.81 min, which also showed significant hyperlipaemia, is to be analysed to structural and detailed functional studies in order to find out whether it is already reported AKH/RPCH member (Fig. 3). This result indicates the presence of Locmi-AKH-I in the retrocerebral extract of H. banian.
Most of the grasshoppers and locust species, in which AKH occur, including the rice grasshopper H. banian in the present investigation, contain one decapeptide (Locmi-AKH-I, Phymo-AKH-I, Phyle-CC, Rommi-CC or Locmi-HrTH) in their corpora cardiaca (Table 1). It also been seen that the AKHs Locmi-AKH-I is present in most of the subfamilies of family Acrididae investigated so far (Table 2). The present investigations in the rice grasshopper, H. banian substantiate the above results, i.e., the insect possesses, Locmi-AKH-I in its CC (Table 3).
| Table 3: | Adipokinetic decapeptides identified from Orthoptera |
 | |
Thus, it is assumed that the genes coding for this AKH is highly conserved in the subfamilies of Acrididae during the course of evolution. Phylogenetic studies using molecular markers (Mitochondrial DNA) have been successfully carried out in Horseshoe crab (Kamaruzzaman et al., 2011) and cereal aphids (Helmi and Khafaga, 2011). Gade and Marco (2009) analysed the phylogeny of ensifera (Insecta, Orthoptera) and Odonata (Gade et al., 2003) using the structural information of adipokinetic neuropeptides in conjunction with morpho-anatomical characters, these data are interpreted in a phylogenetic framework. Similar investigations may be helpful in the construction of phylogenetic tree of the order Orthoptera.
CONCLUSION
The present investigation demonstrated the presence of adipokinetic neuropeptide hormone Locmi-AKH-I in the retrocerebral extract of rice grass hopper, Hieroglyphus banian. With the advent of techniques such as MALDI-TOF/MS-MS, elucidation of insect neuropeptides has been of late interest among insect endocrinologists. Structural and functional similarities of these peptides have enthused them to draw phylogenetic trees to assign relationships. At the same time, similarity in the amino acid sequence of these metabolically crucial peptide hormones also prompted molecular biologists to explore the possibility of employing genetic engineering and biotechnology to develop methods to bring in hormone imbalance in insects leading to self-killing. Structural information also enables to synthesize non-toxic agonists and antagonists that can disrupt hormone balance. If more of insect pest species have similar peptides, development of a few such compounds can be targeted at a variety of pests. Therefore, more and more pest species are to be screened for the presence and structural similarity of the metabolic peptide hormones in them.
ACKNOWLEDGMENTS
We thank Dr. G. Gäde, University of Cape Town, South Africa, for his valuable suggestions. The authors gratefully acknowledge the financial support from the UGC, New Delhi (to MG) and equipment fund from KSCSTE, Thiruvananthapuram, Kerala.
REFERENCES
- Ajaykumar, A.P. and M. Gokuldas, 2011. Elucidation of the primary structure of an adipokinetic neuropeptide from the coffee locust, Aularches miliaris L. (Pyrgomorphidae: Orthoptera). Int. J. Biol. Chem., 5: 127-135.
CrossRefDirect Link - Ajaykumar, A.P. and M. Gokuldas, 2011. Primary structure of an adipokinetic neuropeptide from the rhinoceros beetle, Oryctes rhinoceros L (Coleoptera: Dynastidae). Ann. Neurosci., 18: 100-104.
CrossRefDirect Link - Carlisle, J.A. and B.G. Loughton, 1979. Adipokinetic hormone inhibits protein synthesis in Locusta. Nature, 282: 420-421.
CrossRefDirect Link - Fabold, K., H.I.H. El-Damanhouri and M.W. Lorenz, 2010. Age-dependent cyclic locomotor activity in the cricket, Gryllus bimaculatus and the effect of adipokinetic hormone on locomotion and excitability. J. Comp. Physiol. A: Neuroethol. Sens. Neural Behav. Physiol., 196: 271-283.
CrossRefDirect Link - Frings, C.S., T.W. Fendly, R.T. Dunn and C.A. Queen, 1972. Improved determination of total serum lipid by sulpho-phosphovanillin reaction. Clin. Chem., 18: 673-674.
PubMedDirect Link - Gade, G., 1988. Studies on the hypertrehalosaemic factor from the corpus cardiacum/corpus allatum complex of the beetle, tenebrio molitor. Comparat. Biochem. Physiol. Part A: Physiol., 91: 333-338.
CrossRef - Gade, G., R. Kellner and K.L. Rinehart, 1996. Pyrgomorphid grasshoppers of the genus Phymateus contain species-specific decapeptides of the AKH/RPCH family regulating lipid-mobilization during flight. Physiol. Entomol., 21: 193-202.
CrossRef - Gade, G., H.G. Marco and L. Desutter-Grandcolas, 2003. A phylogenetic analysis of the adipokinetic neuropeptides of Ensifera. Physiol. Entomol., 28: 283-289.
CrossRefDirect Link - Gokuldas, M., P.A. Hunt and D.J. Candy, 1988. The inhibition of lipid synthesis in vitro in the locust, Schistocerca gregaria, by factors from the corpora cardiaca. Physiol. Entomol., 13: 43-48.
CrossRefDirect Link - Gronke, S., G. Muller, J. Hirsch, S. Fellert and A. Andreou et al., 2007. Lipolytic control of body fat storage and mobilisation in Drosophilla. Plos Biol., 5: 1248-1256.
Direct Link - Helmi, A. and A.F. Khafaga, 2011. Molecular fingerprinting of certain cereal aphids in Egypt (Hemiptera: Sternorrhyncha: Aphididae) using RAPD and ISSRs markers. J. Entomol., 8: 327-340.
CrossRefDirect Link - Kamaruzzaman, B.Y., B.A. John, K. Zaleha and K.C.A. Jalal, 2011. Molecular phylogeny of horseshoe crab. Asian J. Biotechnol., 3: 302-309.
CrossRefDirect Link - Kodrik, D. and G.J. Goldsworthy, 1995. Inhibition of RNA synthesis by adipokinetic hormones and brain factor(s) in adult fat body of Locusta migratoria. J. Insect Physiol., 41: 127-133.
CrossRefDirect Link - Kodrik, D., 2008. Adipokinetic hormone functions that are not associated with insect flight. Physiol. Entomol., 33: 171-180.
CrossRefDirect Link - Lorenz, M.W., 2001. Synthesis of lipids in the fat body of Gryllus bimaculatus: Age-dependency and regulation by adipokinetic hormone. Arch. Insect Biochem. Physiol., 47: 198-214.
CrossRefDirect Link - Shuranova, Z.P., Y.M. Burmistrov, J.R. Strawn and R.L. Cooper, 2006. Evidence for an autonomic nervous system in decapod crustaceans. Int. J. Zool. Res., 2: 242-283.
CrossRefDirect Link - Siegert, K.J., R. Kellner and G. Gaede, 2000. A third active AKH is present in pyrgomorphid grasshopper Phymateus morbilosus and Dictyophorus spumens. Insect Biochem. Mol. Biol., 30: 1061-1067.
Direct Link - Stone, J.V., W. Mordue, K.E. Batley and H.R. Morris, 1976. Structure of locust adipokinetic hormone, a neurohormone that regulates lipid utilisation during flight. Nature, 263: 207-211.
PubMedDirect Link - Taub-Montemayor, T.E., K.D. Linse, J.W. Jr. Kent, K.J. Min and M.A. Rankan, 2002. Discovery and characterization of Melanoplus sanguinipes AKH-II by combined HPLC and mass spectrometry methods. J. Biomol. Tech., 13: 219-227.
PubMedDirect Link - Velmurugan, S., P. Soundarapandian N.J. Samuel and S. Rajagopal, 2008. Activity of hyperglycemic hormone in the eyestalk of commercially important crab Portunus pelagicus (Linnaeus). J. Fish. Aquatic. Sci., 3: 77-81.
CrossRefDirect Link - Gade, G. and H.G. Marco, 2009. Peptides of the adipokinetic hormone/red pigment-concentrating hormone family with special emphasis on Caelifera: Primary sequences and functional considerations contrasting grasshoppers and locusts. Gen. Comp. Endocrinol., 162: 59-68.
CrossRefPubMedDirect Link