
A.P. Ajaykumar
Division of Insect Biochemistry and Physiology, Department of Zoology, University of Calicut, Kerala-673 635, India
M. Gokuldas
Division of Insect Biochemistry and Physiology, Department of Zoology, University of Calicut, Kerala-673 635, India
International Journal of Biological Chemistry
Year: 2011 | Volume: 5 | Issue: 2 | Page No.: 127-135







ABSTRACT
In insects, the retrocerebral complex comprises the Corpora Cardiaca (CC) and the Corpora Allata (CA). The CC act as a neurohaemal organ, storing and releasing neuropeptides produced in the brain. The CC also contains intrinsic glandular cells that produce and release several neuropeptides including Adipokinetic Hormones (AKHs). The present investigation reveals the presence of adipokinetic neuropeptides in the coffee locust, Aularches miliaris. The corpora cardiaca extract was fractionated by HPLC, monitoring the eluants both at 210 and 280 nm. Fractions were collected, dried and tested for adipokinetic activity by a homologous in vivo bioassay. Three UV absorbance peaks were found to contain materials with significant adipokinetic activity. MALDI-MS analysis of the extract indicated that the molecular mass, 1167.16Da, is similar to the already known AKH from the insect, Phymateus morbillosus. MALDI-MS/MS analysis confirmed that its primary structure is exactly similar to the structures reported for the Phymo-AKH-I (pE-L-N-F-T-P-N-W-G-S-NH2). Since, A. miliaris is a pest of economically important plants, the knowledge of primary structure of its AKH may be helpful in designing mimetic antagonistic and agonistic of these peptides which may be useful for the control of this pest as well as pests of related groups, employing biotechnological tools.
PDF Abstract XML References Citation
How to cite this article
A.P. Ajaykumar and M. Gokuldas, 2011. Elucidation of the Primary Structure of an Adipokinetic Neuropeptide from the Coffee Locust, Aularches miliaris L. (Pyrgomorphidae: Orthoptera). International Journal of Biological Chemistry, 5: 127-135.
URL: https://scialert.net/abstract/?doi=ijbc.2011.127.135
URL: https://scialert.net/abstract/?doi=ijbc.2011.127.135
INTRODUCTION
Neuropeptides play important roles in cellular communication in vertebrates. This is also true for insects in which many physiological, developmental and behavioral processes are affected by neuropeptides produced in neurosecretory cells of the retrocerebral complex. In insects, the retrocerebral complex comprises the Corpora Cardiaca (CC) and the Corpora Allata (CA). The CC act as a neurohaemal organ, storing and releasing neuropeptides produced in the brain. The CC also contain intrinsic glandular cells that produce and release several neuropeptides including Adipokinetic Hormones (AKHs). AKH/RPCH (red pigment concentrating hormone) family is one of the best known groups of arthropod neuropeptides. This name comes from the first member of the family to be fully characterized, a chromatophorin (RPCH) from prawns (Fernlund and Josefsson, 1972). Common characteristics of the AKHs are: a chain length of 8 to 10 amino acids; the N-terminus blocked by pyroglutamic acid (pGlu); the C-terminus blocked by a carboxyamide; amino acids at positions 8 and 9 (when present) are tryptophan and glycine; most of the peptides are uncharged, but there are a few that have an aspartic acid at position 7; there are at least two aromatic acids, at position 4 mostly phenylalanine (but sometimes tyrosine) and at position 8 tryptophan and a few peptides have a third aromatic acid either at position 2 (Tyr or Phe) or at position 7 (Trp). Known actions of AKHs are, however, broader than their name implies. The peptides have been reported from most of the insect orders. It acts on the fat body to mobilize stored lipids (Ajaykumar and Gokuldas, 2009; Lorenz et al., 2009) and carbohydrates, activate glycogen phosphorylase, accumulate cAMP (Kodrik, 2008) and inhibit the synthesis of lipids (Gokuldas et al., 1988; Lorenz, 2001; Fabold et al., 2010), proteins (Kodrik, 2008) and RNA (Kodrik and Goldsworthy, 1995).
The major objective of the present investigation was to elucidate the primary structures of adipokinetic neuropeptides of the insect, A. miliaris. The primary structures of AKH/RPCH peptides of the insect were elucidated by MALDI-MS/MS in PSD mode. Although, fragmentation by MALDI-MS/MS in PSD mode did not produce all C-terminal and N-terminal ions of the peptides, bioassays of crude extracts of neurohaemal tissues, HPLC fractionations and subsequent bioassays of the fractions provides substantial evidence for peptide confirmation.
MATERIALS AND METHODS
Experimental insect: Adult insects of both sexes and unspecified age of the coffee locust, Aularches miliaris were collected from coffee plantations of Kodak district of Karnataka, India. They were brought to Calicut University campus and maintained in the insectary on a diet of Terminalia cattapa leaves. Food and faecal matters were removed daily. The insects for bioassay were used after one week.
Preparation of corpora cardiaca extract: Corpora cardiaca collected from both sexes of the adult insect, A. miliaris was used as the source of the hormone. Retrocerebral complexes were removed from the head of insects with the help of a pair of fine forceps under a stereozoom binocular microscope. The tissues were immediately put into ice cold 80% methanol (HPLC grade) and stored at -4°C until extraction. Tissues were sonicated for 1 min on ice with an ultrasonicator (Sonics and Materials, USA). The extracts were centrifuged at 4°C and 10,000 rpm for 10 min. The supernatants were collected into an eppendorf tube and vacuum dried (Savant, USA). The dried supernatants were stored at -4°C until used for HPLC separations, bioassay studies and mass spectrometric analysis.
High Performance Liquid Chromatography (HPLC) analysis: The dried extract made from the retrocerebral complexes from the insect was dissolved in 20 of 80% methanol (HPLC grade). The extract was filtered using a sample filtration unit with 0.45 μm filter paper (Millipore). The samples were directly injected into the instrument by a microsyringe (20 μL). HPLC separations were carried out using Shimadzu system (SPD M-10 AVP, LC-10 ATVP, LC-10 ATVP) with a reversed phase column (C18) 250 mm long, 4.6 mm i.d. The separation was done in a binary gradient from 43 to 53% solvent B in 20 min and then to 70% within a further 6 min with a flow rate of 1 mL min-1. Trifluoroacetic Acid (TFA) 0.01% in water (HPLC grade) was used as solvent A. Solvent B was 60% acetonitrile in solvent A. All the solvents were filtered through 0.45 μm filter paper. The eluants were monitored simultaneously at 210 and 280 nm using a Photo Diode Array (PDA) detector. One minute fractions starting from 4 min up to 20 min and that at 25 min were collected manually, dried using vacuum concentrator and were used for testing their hyperlipaemic activity. The HPLC profiles were exported into Microsoft Word file and were used for further analysis of the data.
The presence of AKH peptide, Phymo-AKH-I (GenScript Corp., USA) in the retrocerebral extracts of A. miliaris was investigated by analysing their elution pattern in HPLC. The synthetic peptide, Phymo-AKH-I (100 pmol) was injected into the HPLC instrument maintained in the same set up as before for the retrocerebral extracts of A. miliaris.
Detection of biological activity in the CC extract: The dried methanolic extract prepared was dissolved in insect saline to get a final concentration of one gland pair equivalent (gpe) per 5 μL. A sample of the extract (5 μL) was injected using a Hamilton microsyringe (10 μL) into the haemolymph of the accepter pyrgomorphid grasshopper, A. miliaris. The injection needle was thrust in the intersegmental membrane between thorax and first abdominal segment, ventrally. The hyperlipaemic effects of the extracts were measured by estimating change in the lipid content of the haemolymph after the injection of extract using phosphovanillin method. Haemolymph samples (2 μL) were collected directly from the cut end of the antenna into precalibrated capillary tubes and were then transferred into the bottom of test tubes (5 mL capacity). Haemolymph samples taken before injections (from one antenna) were taken as controls and 60 min after injection (from the other antenna) as experimentals. Similar experiments were carried out using 5 μL of insect saline instead of retrocerebral extract. Haemolymph samples thus collected were used for quantification of lipids.
Quantitation of haemolymph lipids: Total lipids in the haemolymph samples were determined using phosphovanillin reagent (Frings et al., 1972). Haemolymph samples collected (2 μL each) in various experiments were deposited into the bottom of test tubes. Concentrated sulphuric acid (50 μL) were added to these samples, heated in a boiling water bath for 10 min, cooled to room temperature and 2 mL of each phosphovanillin reagent were added. The tubes were thoroughly shaken to mix the content. Optical densities were measured within 5 min using UV-vis spectrophotometer at 540 nm against a reagent blank.
Matrix Assisted Laser Desorption Ionisation-time of Flight-Mass Spectrometry (MALDI TOF-MS): The dried extract of neurohaemal tissues of the insect, A. miliaris was used for mass spectrometric analysis. Mass spectrometric analysis were performed on an Ultra Flex mass spectrometer (Bruker Daltonics, Germany) in reflectron ion mode, using a 90 ns time delay and a 25 kV accelerating voltage in the positive ion (Na+) mode. The system utilized 50 Hz pulsed voltage laser, emitting at 337 nm. The ion source and the flight tube were kept at pressure of about 7x10-7 mbar by turbo molecular pump. The samples were prepared by mixing equal volumes of peptide solution and a saturated solution of the matrix, dihydroxybenzoic acid in 1:1 (v/v) acetonitrile: water mixture. A standard peptide mixture was used for external calibration.
Tandem-MS/MS: Tandem Mass Spectra (MS/MS) were acquired by selecting the precursor mass (1167.16 Da) with a 10 Da window and fragments were generated in Post Source Decay (PSD) mode. A single acquisition was a sum of 360 added shots to generate the MS/MS spectra. Mass spectra were analysed by using Flex-analysis software. The MS/MS data were interpreted by the aid of Peptide Fragmentation Ion Analyser-II (PFIA-II) software (http://hodgkin.mbu.IIsc.ernet.in/~pfia/PFIA-11.html).
Statistical analysis and data presentation: Results obtained from various hyperlipaemic bioassay experiments were expressed as Mean±SE values as well as percentage difference of the experimentals over controls (E/C%). The paired t-test and results of adipokinetic responses were performed with the use of SPSS Software (version 10). The graphical representation of change in lipid mobilization was plotted by Microsoft Excel programme. The data were obtained from the research work conducted in the Department of Zoology University of Calicut, Kerala, India and Indian Institute of Science, Bangalore, India from July 2008 to February 2009.
RESULTS
HPLC separation and biological activity of fractions: HPLC analysis of the methanolic extract of CC from A. miliaris showed three major absorbance peaks with retention times 8.75, 10 and 24 min, respectively (designated as 1, 2 and 3 in Fig. 1). The materials in the fractions collected during 9, 10 and 24 min showed significant hyperlipaemic effects, giving increases up to 38 (p<0.05), 19 (p<0.05) and 17% (p<0.05), respectively over the controls. The hyperlipaemic activities exhibited by the other fractions, viz., 8, 11, 12, 13, 14, 15, 16, 18, 19 and 20 min were not statistically significant (Fig. 2). The data were compared with the HPLC profile of retrocerebral extract and the results are summarized in the Fig. 3. As seen in the chromatogram (Fig. 3a, b) Phymo-AKH-1 was eluted at same retention time as that of the material having the retention time 8.75 min in the retrocerebral extract of A. miliaris.
MALDI-MS analysis: Figure 4 represents the MALDI-MS spectra of extracts of retrocerebral complexes of A. miliaris in Na+ mode. The results showed that the molecular ion peak with m z-1 at 1167.16 (Na+ mode) obtained represent a peptide with molecular mass 1144.53 Da. A similar peptide has been reported from other pyrgomorphid grasshoppers with code name, Phymo-AKH-I. The primary structure of the molecule was further elucidated by MALDI-MS/MS analysis in PSD mode.
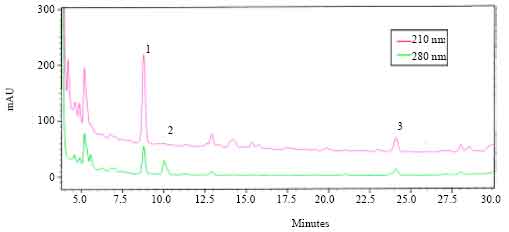 | |
| Fig. 1: | The HPLC profile of extracts of retrocerebral complexes of A. miliaris. The analysis was carried out on a C18 hiber column. The extract was run with a gradient of 43-53% B in 20 min and then to 70% B within a further 6 min (solvent A = 0.01% trifluoro acetic acid in water, solvent B = 60% acetonitrile in solvent A). The eluants were monitored at 210 and 280 nm |
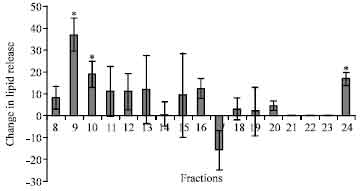 | |
| Fig. 2: | The HPLC fractions were collected and tested for hyperlipaemic activity. The change in total haemolymph lipid is represented as E/C% in bardiagram. (*)Indicates p<0.05 |
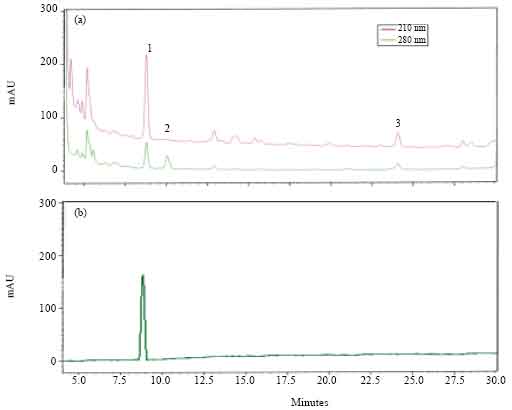 | |
| Fig. 3: | The HPLC profiles of crude corpora cardiaca extract of A. miliaris (a) and synthetic phymo-AKH-1 (b). The extract was run with a gradient of 43-53% B in 20 min (solvent A = 0.01% trifluoro acetic acid in water, solvent B = 60% acetonitrile in solvent A) |
MALDI MS/MS of precursor ion at 1167.16 m z-1: The MALDI-MS/MS fragmentation spectrum of precursor ion at 1167.16 m z-1 is given as Fig. 5. The theoretical fragmentation pattern was deduced from PFIA-II software. The N-terminal ‘b’ type ion fragments are b1 (112 m z-1), b3 (339 m z-1), b4 (486 m z-1), b5 (587 m z-1), b8 (007.4 m z-1) and b9 (1041 m z-1). The ‘a’ type ions are a2 (197 m z-1) and a5 (559 m z-1). The identified C-terminal y type fragment ions are y1 (105 m z-1), y3 (348 m z-1), y4 (462 m z-1), y5 (559 m z-1), y6 (660 m z-1) and y9 (1034)m z-1. The immonium ions of amino acids identified were glycine (30.03 m z-1), proline (70.06 m z-1), threonine (74.06 m z-1), leucine (80.09 m z-1) and phenylalanine (120.01 m z-1). Thus the primary structure of the precursor ion is derived as pE-L-N-F-T-P-N-W-G-S-NH2.
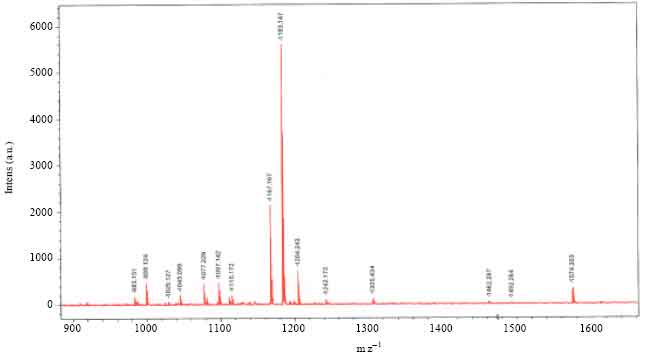 | |
| Fig. 4: | MALDI-MS spectrum of extract of corpora cardiaca of A. miliaris. The analysis was carried out in reflector positive (Na)+ mode with an accelaration voltage of 50 Hz pulsed N2 laser, emitting at 337 nm. Dihydroxy benzoic acid was used as matrix |
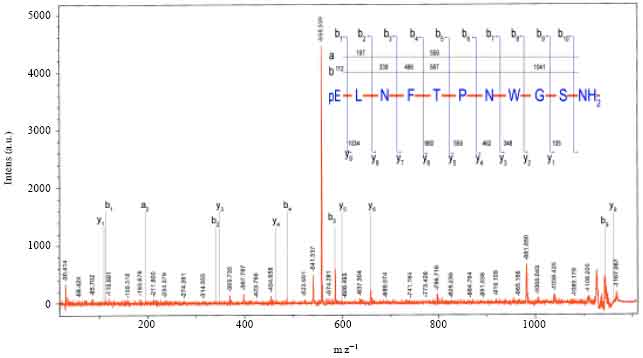 | |
| Fig. 5: | MALDI MS/MS spectrum of the ion (M+Na) = 1166.53 Da from A. miliaris. Inset shows the sequence assignment of the peptide, together with theoratical and calculated masses for b, y and a type fragment ions, obtained in the MS/MS spectrum |
DISCUSSION
The RP-HPLC separation and further bioassays of fractions of retrocerebral complexes of A. miliaris showed the presence of a few UV-absorbing peaks exhibiting biological activity (Fig. 1 and 2). The biologically active materials with retention times 0.75, 10 and 24 min in HPLC (Fig. 1) showed absorbance peaks at 210 and 280 nm in a PDA detector. Peptide bonds and the aromatic amino acids, give maximum absorbance at 210 and 280 nm respectively. All members of AKH/RPCH family of peptides have been shown to have a tryptophan residue at position 8 (Gade, 1997). The comparison of HPLC profiles of the synthetic Phymo-AKH-1 with that of A. miliaris retrocerebral complex extract showed that the extract contained materials having similar retention times (Fig. 3). This data indicate the presence of Phymo-AKH-1 in the retrocerebral complex extract of A. miliaris. The present study identified the primary structure of one adipokinetic peptide from the corpora cardiaca of the pyrgomorphid grasshopper, A. miliaris.It is a decapeptide of AKH/RPCH family. Similar peptide was first identified from a pyrgomorphid grasshopper Phymateus morbillosus and code named as Phymo-AKH-I with amino acid sequence pE-L-N-F-T-P-N-W-G-S-NH2. Gade et al. (1996) identified two other biologically active AKH peptides, Schgr-AKH-II and Phymo-AKH-III from the same insect. Even though the HPLC profile of retrocerebral extract of A. miliaris gave more than one biologically active peaks both at 210 and 280 nm (Fig. 1), we could not identify the molecular masses (933.43 Da; Schgr-AKH-II and 1072.5 Da; Phymo-AKH-III) corresponding to these peptides. Since regulation of intermediary metabolism in grasshoppers appears to be very complicated (Siegert et al., 2000), it is suggested that there may be other novel AKH peptides (Ajaykumar and Gokuldas, 2008) than those already detected that are involved in mobilizing energy reserves (lipids and carbohydrates) and this may be true with A. miliaris. However, the pyrgomorphid grasshopper Phymateus leprosus, contains only a single AKH, Phyle-CC. Hence, it will be worth investigating to find out the novel peptides whose biological activities have been confirmed but the structures have not been able to be figured out.
Most of the insect species found to have three or more AKHs, are mostly active fliers (Gade, 2006). Vroemen et al. (1998) pointed out that long distance flight in L. migratoria is attributed to the multiplicity of AKH neuropeptides in its CC. The coffee locust A. miliaris is not a continuous flier, it flies very short distances. Thus it is suggested that the AKHs in A. miliaris may be helpful for comparatively less energy requiring activities like, short distance flight, hopping and walking. Results from studies in P. apterus and G. bimaculatus are in agreement with the above suggestion, i.e., in these insects AKHs are useful for activities such as walking (Socha et al., 1999; Lorenz et al., 2004), the energy demand for which is comparatively less. In the present investigation, HPLC separation and further fractionation of extracts of Aularches retrocerebral complexes indicated the presence of more than one adipokinetic factor in them. However, by MALDI-MS analysis, we could not find any molecular mass corresponding to already known AKHs from other Orthopteran species.
More than fifteen AKH decapeptides were elucidated from various insect orders with various degrees of replacement of amino acids. The peptide Phymo-AKH-I is most similar in structure (90%) to Locmi-AKH-I (threonine is replaced by serine at 10th position) and Phyle-CC (threonine is replaced by asparagine at 3rd). The amino acid exchange at position 3 has been observed in many AKH/RPCH family peptides and is a conservative modification (Gade et al., 1994). The substitution at position 10, where four amino acid residues (Serine, Asparagine, Threonine and tyrosine) have been found to get exchanged in the AKH/RPCH peptides (Gade et al., 1994) cannot be explained easily by single step point mutation.
| Table 1: | Orthopteran insects which possess Phymo-AKH-I in their CC |
 | |
Phymo-AKH was identified earlier from two other insects, P. morbillosus and Dictyophorus spumens coming under the same insect family, Pyrgomorphidae (Table 1). These results show that the distribution of Phymo-AKH has some family specificity. Such data have been used as additional information to aid in the construction of phylogenetic trees by means of computer programme and protein parsimony algorithms. Gade and Heather (2005) analysed the phylogeny of the insect order Odonata, based on the primary structures of AKH peptides present in that group, two species, P. morbillosus and P. leprosus coming under the same genus (Phymateus) exhibited the presence of two different decapeptides (Phymo-AKH-I and Phyle-CC) in their CC. Since the data regarding AKH complements in different species of these groups are not completely available, further studies are essential to make conclusions regarding the phylogeny of this group.
ACKNOWLEDGMENT
We thank Council of Scientific and Industrial Research (CSIR), Government of India, for the award of scholarship to A.P. Ajaykumar., Mr. Subrahmanyam Prakash, Indian Institute of Science, Bangalore, for mass spectrometric analysis and equipment fund by KSCSTE Thiruvananthapuram, Kerala.
REFERENCES
- Ajaykumar, A.P. and M. Gokuldas, 2008. Elucidation of primary structures of three adipokinetic neuropeptides from the corpora cardiaca of the paddy pest Oxya nitidula (Orthoptera: Acrididae). Proceedings of the International Structural Neuroscience Conference on Peptides, Feb. 2-3, Nagpur, India, p: 16-16.
- Fabold, K., H.I.H. El-Damanhouri and M.W. Lorenz, 2010. Age-dependent cyclic locomotor activity in the cricket, Gryllus bimaculatus and the effect of adipokinetic hormone on locomotion and excitability. J. Comp. Physiol. A: Neuroethol. Sens. Neural Behav. Physiol., 196: 271-283.
CrossRefDirect Link - Fernlund, P. and L. Josefsson, 1972. Crustacean color-change hormone: Amino acid sequence and chemical synthesis. Science, 177: 173-175.
CrossRef - Frings, C.S., T.W. Fendley, R.T. Dunn and C.A. Queen, 1972. Improved determination of total serum lipids by the sulfo-phospho-vanillin reaction. Clin. Chem., 18: 673-674.
CrossRefPubMedDirect Link - Gade, G., 1997. The explosion of structural information on insect neuropeptides. Fortschr. Chem. Org. Naturst., 71: 1-128.
PubMed - Gade, G. and G.H. Heather, 2005. The adipokinetic hormones of Odonata: A phylogenetic approach. J. Insect Physiol., 51: 333-341.
PubMed - Gade, G., 2006. Unusual complement of three AKH octapeptides from in two species of grasshoppers (Caelifera). Eur. J. Entomol., 103: 297-304.
Direct Link - Gade, G., R. Kellner and K.L. Rinehart, 1996. Pyrgomorphid grasshoppers of the genus Phymateus contain species-specific decapeptides of the AKH/RPCH family regulating lipid-mobilization during flight. Physiol. Entomol., 21: 193-202.
CrossRef - Gokuldas, M., P.A. Hunt and D.J. Candy, 1988. The inhibition of lipid synthesis in vitro in the locust, Schistocerca gregaria, by factors from the corpora cardiaca. Physiol. Entomol., 13: 43-48.
CrossRefDirect Link - Kodrik, D., 2008. Adipokinetic hormone functions that are not associated with insect flight. Physiol. Entomol., 33: 171-180.
CrossRefDirect Link - Kodrik, D. and G.J. Goldsworthy, 1995. Inhibition of RNA synthesis by adipokinetic hormones and brain factor(s) in adult fat body of Locusta migratoria. J. Insect Physiol., 41: 127-133.
CrossRefDirect Link - Lorenz, M.W., 2001. Synthesis of lipids in the fat body of Gryllus bimaculatus: Age-dependency and regulation by adipokinetic hormone. Arch. Insect Biochem. Physiol., 47: 198-214.
CrossRefDirect Link - Lorenz, M.W., R. Zemek, D. Kodrik and R. Socha, 2004. Lipid mobilization and locomotor stimulation in Gryllus bimaculatus (Ensifera, Gryllidae) by topically applied adipokinetic hormone. Physiol. Entomol., 29: 146-151.
CrossRef - Siegert, K.J., R. Kellner and G. Gaede, 2000. A third active AKH is present in pyrgomorphid grasshopper Phymateus morbilosus and Dictyophorus spumens. Insect Biochem. Mol. Biol., 30: 1061-1067.
Direct Link - Socha, R., D. Kodrik and R. Zemek, 1999. Adipokinetic hormone stimulates insect locomotor activity. Naturwissenschaften, 88: 85-86.
CrossRefDirect Link - Vroemen, S.F., D.J.V. Horst and W.J.A.V. Marrewijk, 1998. New insights into adipokinetic hormone signaling. Mol. Cell. Endocrinol., 141: 7-12.
CrossRef