
L.N. Ireri
Division of Vector Borne and Neglected Tropical Diseases, Ministry of Public Health and Sanitation, P.O. Box 1905-60100, Embu, Eastern Province, Kenya
J. Kongoro
Department of Zoological Sciences, School of Pure and Applied Sciences, Kenyatta University, P.O. Box 43844, Nairobi, Kenya
P. Ngure
School of Science, Engineering and Health, Daystar University, P.O. Box 44400-00100, Nairobi, Kenya
K.S. Sum
Pyrethrum Board of Kenya, P.O. Box 420-20100, Nakuru, Kenya
W. Tonui
Centre for Biotechnology Research and Development, Kenya Medical Research Institute, P.O. Box 54840-00200, Nairobi, Kenya
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 6 | Page No.: 581-587







ABSTRACT
Breeding sites of sand flies are complex; consequently using conventional larvicides to control larvae is not realistic and need improved strategies. A study was therefore conducted to evaluate susceptibility of immature stages of Phlebotomus duboscqi Neveu Lemaire (Diptera: Psychodidae), to two pyrethrin formulations incorporated in larval food; an emulsifiable concentrate (0.5%) used as a mosquito larvicide and a dusting powder (0.2%) normally used against crawling insects. The concentrate was prepared into 0.2, 0.5 and 1 mg mL-1 and each mixed with 1g of larval food, dried overnight and introduced into vials containing 10 larvae per instar. Similar concentrations of pyrethrin powder were prepared, mixed with 1 g of larval food then kept overnight and introduced in similar vials as above. All the experiments were done in triplicate and mortality recorded daily. The larvae showed significant mortality (p<0.05) effects to the two formulations at 48 h of exposure. Second and 3rd instars were more susceptible to Moskil® than Pylarvex™ with LD50 of 0.23 and 0.3 mg mL-1 for 2nd instars and 0.1 and 0.14 mg mL-1 for 3rd instars in 0.2 mg bioassays, respectively. However, there was no significant difference (p>0.05) in mortality between formulations and instars but significantly (p<0.05) differed between concentrations used. None of the larval stages managed to molt to the next stage, hence no pupal stages were observed. The results of this study demonstrate the clear entomological efficacy of pyrethrin formulations in larval food and should be considered for use in integrated leishmaniasis vector control programmes.
PDF Abstract XML References Citation
Received: March 06, 2011;
Accepted: June 04, 2011;
Published: August 12, 2011
How to cite this article
L.N. Ireri, J. Kongoro, P. Ngure, K.S. Sum and W. Tonui, 2011. Insecticidal Properties of Pyrethrin Formulations against Immature Stages of Phlebotomine Sand Flies (Diptera: Psychodidae). Journal of Entomology, 8: 581-587.
DOI: 10.3923/je.2011.581.587
URL: https://scialert.net/abstract/?doi=je.2011.581.587
DOI: 10.3923/je.2011.581.587
URL: https://scialert.net/abstract/?doi=je.2011.581.587
INTRODUCTION
Larviciding is a key strategy used in many disease vector control programmes around the world. Phlebotomine sand fly larvae are difficult to find since they occupy obscure terrestrial habitats and only scarce information is available on their ecology. Sand flies are considered to play important roles as vectors of Leishmania, Bartonella and phleboviruses that afflict humans and other vertebrates in tropical and subtropical regions of the world (Feliciangeli, 2004; Kakarsulemankhel, 2004). The difficulty of finding breeding sites is an important constraint in vector control measures available to leishmaniasis control programmes, application of conventional larvicides not being a practical alternative (Ashford, 1999; Mascari et al., 2007). Unlike in most biting Dipteran flies, development of sand flies takes place in a wide variety of terrestrial habitats rather than aquatic microhabitats (Modi and Tesh, 1983), making it difficult to design and implement control strategies targeting the immature stages. Larvae of some phlebotomines have been found in specialized niches such as Lu. ovallesi on buttress roots of trees in Panama; P. celiae, P. martini in termite hills and P. duboscqi in animal burrows in Kenya; P. longipes and P. pedifer in caves and among rocks in Eastern Africa (Ngumbi et al., 1998; Feliciangeli, 2004). These are rich in organic matter which larvae feed on passing through four instars, pupate and eventual adult emergence. They play a major role in the ecology of phlebotomine sand flies (Ngumbi et al., 1998). Unlike unknown sites, these can be targeted for the application of larvicides. Spraying of conventional insecticides targeting the immature stages of sand flies has not been successful because application of insecticides into rodent burrows does not reach the microhabitats due to complexities and lengths of the tunnels (Karapet’ian et al., 1983; Mascari et al., 2007). Development of new efficacious formulations and methods are in dire need and especially use of powdered formulations and fumigants. Powdered and aqueous insecticidal formulations have been used successfully to control insect pests of economic and medical importance and can as well be used in sand fly control. Pyrethrins are normally used to fortify commercial preparations of household insecticides and are low in toxicity to mammals; effective against many insects and are easily biodegradable to harmless substances. Pyrethrins persist much longer in enclosed quarters than they do out in the open. Previous studies have shown that pyrethrins persists for 60 h after treatment on horizontal surfaces, two weeks after treatment on airborne particles and over two months in carpet dust (Class and Kintrup, 1991). A recent study however has shown that low pyrethrin concentrations incorporated in sucrose baits is lethal to adult Phlebotomus duboscqi (Ireri et al., 2010). Other materials including pyrethroids have been successfully evaluated against sand flies (Al-Barrak, 2005). The aim of this study was to evaluate the potential of natural pyrethrin, abundantly grown in Kenya, in combinations with artificial larval food to control sand fly larvae. The study was conducted from June 2007 to December 2009. In Kenya leishmaniases are transmitted by P. martini and P. duboscqi in several foci.
MATERIALS AND METHODS
Larvae for bioassay: Larvae for the experiments were obtained from rearing female Phlebotomus duboscqi got from the Centre for Biotechnology Research and Development in sectaries in Kenya Medical Research Institute Nairobi. These were blood fed using Syrian golden hamsters, Mesocricetus auratus which were regularly introduced in the rearing cages. Engorged females were maintained with corn syrup, Karo® drops placed on top of the screened oviposition jar lids and allowed to lay eggs. Larvae when hatched were fed on a mixture of an equal amount of rabbit chow and droppings, fermented and ground to fine powder until they attained desired stages.
Test materials: Two commercial products of Chrysanthemum cinerariefolium, pyrethrin EC 0.5% w/v (Pylarvex™) and pyrethrin 0.2% w/w (Moskil® dusting powder) were acquired from the manufacturer, Pyrethrum Board of Kenya, of P.O. Box 420, Nakuru, Kenya. The pyrethrin concentrate is normally used as mosquito larvicide while pyrethrin powder is normally used for controlling crawling insects.
Bioassays: The technique used was as previously described (Luitgards-Moura et al., 2000) and similar to the one generally applied for the bioassays of Spodoptera larvae (Ikbal et al., 2007). Ten P. duboscqi larvae were gently placed into four triplicate series of vials using a camel hair brush wetted in distilled water to avoid stress and damage. The experiment was arranged such that first triplicate series contained 1st instar larvae, the second triplicate 2nd instar larvae, the third triplicate contained the 3rd instar larvae and the fourth triplicate contained the 4th instar larvae in each vial. Measures of one gram each of larval food prepared from a fungal growth obtained from rabbit droppings-chow mixture were meticulously mixed with prepared concentrations of 0.2, 0.5 and 1.0 mg g-1 of pyrethrin powder and sprinkled into the vials. Similar concentrations were prepared from the pyrethrin EC. These were mixed with a gram of larval food each, thoroughly homogenized, spread on a sheet of paper and let to dry under shade overnight. The food mixtures were sprinkled into the vials each day as the larvae feed. The larvae that fed on larvae food alone in the first bioassay formed the control group while in the second; food was wetted and homogenized with distilled water, dried under shade and introduced into the vials containing the larvae. The larvae were observed under binocular microscope daily. Mortality defined as luck of response to prodding with a blunt pin after 15 sec was recorded. Abnormal behavior, morphological characteristics, evidence of feeding and the presence of frass in the experimental vials were also noted (Mascari et al., 2007). The experiments were monitored and mortality recorded daily for analysis. Mean lethal dosage designated LD50, was determined every 24 h of exposure.
Statistical analysis: All data were entered into Microsoft excel program. Data on the larvae mortality were subjected to computerized Probit analysis (PROBIT model: (PROBIT (p)) = Intercept + BX): for LD50 values on all bioassays (Finney, 1971). Confidence limits for effective dosages were set at 95% probability level. Variation effects between different larval instars were compared using ANOVA (Edwards, 1960).
RESULTS
Observations done throughout the exposure period to pyrethrin which act on insect’s nervous system revealed that the larvae fed and moved about normally in the first 24 h in both formulations.
Mortality of P. duboscqi larvae due to 0.5% pyrethrin (Pylarvex™) in food at 48 h of exposure: This product had the most rapid mortality to first and fourth instar larvae than the powdered pyrethrin. More deaths were noted in 1st instar larvae than others in 48 h of exposure with LD50 of 0.19 mg mL-1. It was however observed that 4th instar larvae fed less avidly than all the others but had more mortality than 4th instar larvae in the powdered pyrethrin bioassay which had a higher LD50 value (1.33 mg g-1). Larval mortality did not significantly differ in the various stages of the instars (F = 0.1, p = 0.962). Probit analysis showed varied larvicidal activity values for 1st, 2nd, 3rd and 4th instar larvae to this product (Table 1). Second instar larvae had generally low mortality at 48 h than all the others in the assays. After 48 h mortality increased and was dose dependent.
Mortality of larvae due to Moskil® pyrethrin powder at 48 h of exposure: Moskil is a relatively known pyrethrin in the Kenyan market used mainly in the control of many crawling insects including coach roaches, fleas, chicken louse and ants and had generally significantly less (p<0.05) LD50 like Pylarvex. The LD50 values for second and third instar larvae in Pyrethrin and Moskil bioassays were comparable and were between 0.1 and 0.29 mg mL-1.
| Table 1: | Larvicidal activity of liquid pyrethrin (Pylarvex) against first, second, third and fourth instars larvae of P. duboscqi when incorporated in larval food at 48 of exposure |
 | |
| LD: Lethal dosage (set at 95% confidence level), between instars F = 0.1, p = 0.962 | |
| Table 2: | Larvicidal activity of powdered pyrethrin (Moskil) against first, second, third and fourth instars larvae of P. duboscqi when incorporated in larval food at 48 h of exposure |
 | |
| LD: Lethal dosage (set at 95% confidence level), between instars F = 2.0, p = 0.173 | |
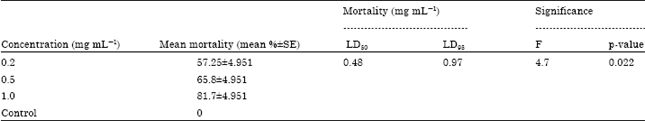
| Table 3: | Mortality of P. duboscqi larvae when fed on larval food mixed with different concentrations of powdered pyrethrin (Moskil) concentrations |
 | |
| LD: Lethal dosage (set at 95% confidence level), overall mean = 51.25%, coefficient of variation -64.4% | |
Using Moskil pyrethrin powder, this study established there was no significant difference in the mortality of larval stages of sand flies (F = 2.0, p = 0.173) at 48 h of exposure. However, 3rd instars were the most affected stage of the larvae with a mortality mean of 70%). Analysis also showed varying mortality to the powdered formulation (Table 2).
Concentrations of the Moskil® pyrethrin powder used: Observations observed on the larvae when exposed to different concentrations of Moskil revealed a similar manner of toxicity to that of Pyrethrin. Larvae were still active after 24 h but became lethargic in movement there after until they died. It was found out that there was significant effect of concentrations of the moskil pyrethrin powder used (F = 4.7, p = 0.022). The higher the concentration of the test material the more larval mortality was experienced. Concentration of 1.0 mg g-1 had the highest mean of 81.7% followed by 0.5 mg g-1 with a mean mortality of 65.8% and subsequently by 0.2 mg g-1 with a mean mortality of 57.25%. No mortality was observed in the control group. The overall mean was 51.25% as shown in Table 3.
DISCUSSION
The results obtained in the evaluation of pyrethrum product showed that first instar larvae were more susceptible to pylarvex™ EC than moskil® powder. This could be attributed to permeability and homogeneity of the active ingredients in the larval food as compared to the physical mixing of the powdered formulation and the larval food. It was observed that first and 4th instar larvae fed less avidly than 2nd and 3rd instars. The behaviour of 4th instar larvae to feed reluctantly could be attributed to the fact that they were able to withstand long hours without feeding in preparation to undergo pupation or other factors not tested here. There was no mortality of 4th instars in the first 24 h in the powdered pyrethrin bioassays and also in the 0.5 mg g-1 group at 48 h. Mortality in all the other instars was significantly high (p<0.05) in both pyrethrin-larval food preparations. At 72 and 96 h of exposure, results of the two formulations were however comparable and had more than 80% mortality in some instars. Comparing the two formulations, dilute emulsifiable concentrates were fast in action than powders but not statistically significant. The larvae showed evident physiological changes twenty four hours after feeding on the food mixtures. In all the bioassays, all the larval instars succumbed to the preparations before molting to the next stage; hence pupal stages were not realized. This is in support of a study conducted by Mascari et al. (2007) which found out that no pupation or adult emergence occurred when larvae of P. papatasi were fed feces of hamsters containing novaluron. The larvae gradually shortened and changed from translucent to opaque, looking decrepit and scarcely feeding until they died. The controls were followed up to pupation and eventual adult emergence successfully, depicting the potential of pyrethrin combination with other materials to control immature stages of sand flies. This study is also in support with similar works using Bacillus sphaericus Neide toxins fed to sand fly adults while in sucrose and were subsequently eaten by larvae produced significant larval death (p<0.01) (Robert et al., 1997) in the laboratory set up. Other insecticidal compounds have been evaluated and found similarly effective and comparable against sand fly larvae (Mascari et al., 2010). There is no literature available on the use of pyrethrin against sand fly larvae and hence comparative data could not be obtained. However, similar extracts of botanical origin with similar properties have been evaluated against other organisms including mosquito larvae (Vinayagam et al., 2008; Shonouda and Mehanney, 2000; Chakkaravarthy et al., 2011) which died as larvae in higher concentrations, soil nematodes (Katooli et al., 2010) and as antimicrobial agents (Mariita et al., 2011). Use of such material could form efficient means of controlling sand flies by exploiting their habitat, intandem rendering them unreceptive. Control and prevention of leishmaniases depends on the prevailing local epidemiological characteristics. As a rule of thumb, control measures are the result of breaking one or more elements in the life cycle (Alvar et al., 2006), in this case we targeted the larval stages. There is no single method that can be used for all situations and one method may be successful in one place but fail in another and thereby necessitating change and/or modification of application methods. In addition, cost effectiveness has to be considered before adopting a certain method (Tonui, 2006) and pyrethrins are cheap and easily acquired. Moreover, control measures should always be revised and evaluated. Although some trials target reservoirs by eliminating the rodents, destruction of the animals’ food sources and /or ploughing out burrows and treatment of dogs, measures targeting the vector needs to be modified to embrace and circumvent possible vector resistance to available insecticides which has been witnessed in some cases lately.
CONCLUSIONS
Due to limited information on vector competence, seasonal abundance patterns, species association, disciplines and development, designing new control strategies may entail vigorous studies to embrace the current trend in vector bionomics (Rassi et al., 2005). Only limited information is available regarding bioassays on phlebotomine sand fly larvae. In conclusion pyrethrum products showed potent properties and should be formulated into fine powder, incorporated into dry powdered plant parts or animal waste products as food baits and blown into breeding sites. Research into possible incorporation into rodent feeds or termite baits for use in feed-through practices to control sand fly larvae are warranted.
ACKNOWLEDGMENTS
We wish to thank Kasili Sichangi, Chris Anjili, Phillip Ngumbi, Josyline Cirindi and Stella Kepha of Centre for Biotechnology Research and Development, KEMRI; Obadiah Mucheru and Paul Kamau of Ministry of Public Health and Sanitation for their valuable suggestions on set up and organizational support.
This study received financial support from the International Foundation for Science (IFS) Karlavägen 108, 5th floor SE-115 26 Stockholm, Sweden and the Organization for the Prohibition of Chemical Weapons, The Hague (OPCW), through Dr. Willy Tonui of KEMRI, Nairobi, Kenya [Grant No. F/3894-1].
REFERENCES
- Feliciangeli, M.D., 2004. Natural breeding places of phlebotomine sandflies. Med. Vet. Entomol., 18: 71-80.
PubMedDirect Link - Kakarsulemankhel, J.K., 2004. Composition of the Phlebotominae, fauna (Diptera, Psychodidae) in Balochistan, Pakistan. J. Biological Sci., 4: 391-392.
CrossRefDirect Link - Ashford, R.W., 1999. Cutaneous leishmaniasis: Strategies for prevention. Clin. Dermatol., 17: 327-332.
CrossRefPubMedDirect Link - Mascari, T.M., M.A. Mitchell, E.D. Rowton and L.D. Foil, 2007. Evaluation of novaluron as a feed-through insecticide for control of immature sand flies (Diptera: Psychodidae). J. Med. Entomol., 44: 714-717.
PubMedDirect Link - Modi, G.B. and R.B. Tesh, 1983. A simple technique for mass rearing Lutzomyia longipalpis and Phlebotomus papatasi (Diptera: Psychodidae) in the laboratory. J. Med. Entomol., 20: 568-569.
PubMedDirect Link - Ngumbi, P.M., L.W. Lrungu, L.I. Robert, D.M. Gordon and J.L. Githure, 1998. Abundances and nocturnal activities of phlebotomine sandflies (Diptera: Psychodidae) in termite hills and animal burrows in Baringo district, Kenya. Afr. J. Health Sci., 5: 28-34.
PubMedDirect Link - Karapet'ian, A.B., L.N. Dzhabarov, R.I. Mamigonova and E.A. Sabatov, 1983. Use of D-20 aerosol insecticide smoke pots in controling burrow sandflies. Med. Parazitol. (Mosk), 4: 78-81.
PubMedDirect Link - Class, T.J. and J. Kintrup, 1991. Pyrethroids as household insecticides: Analysis, indoor exposure and persistence. Fresenius J. Anal. Chem., 340: 446-453.
CrossRefDirect Link - Ireri, L., O. Mucheru, P. Ngure, S. Kepha, A. Kimutai, J. Cirindi and W. Tonui, 2010. Potential effects of pyrethrin incorporated in sucrose baits against phlebotomus duboscqi neveu lemaire (Diptera: Psychodidae) in leishmaniasis control strategies. WebmedCentral Parasitology., 1: 1-8.
Direct Link - Al-Barrak, A.S., 2005. A study on abundance and control of sandflies (Diptera: Psychodidae) in Al-Gassim region, Saudi Arabia. Pak. J. Biol. Sci., 8: 326-329.
CrossRefDirect Link - Luitgards-Moura, J.F., E.G.C. Bermudez and M.G. Rosa-Freitas, 2000. related to productivity for four generations of Lutzomyia longipalpis laboratory colony Mem. Inst. Oswaldo Cruz., 95: 251-257.
PubMedDirect Link - Robert, L.L., M.J. Perich, Y. Schlein, R.L. Jacobson, R.A. Wirtz, P.G. Lawyer and J.I. Githure, 1997. Phlebotomine sand fly control using bait-fed adults to carry the larvicide Bacillus sphaericus to the larval habitat. J. Am. Mosq. Control Assoc., 13: 140-144.
PubMedDirect Link - Mascari, T.M., M.A. Mitchell, E.D. Rowton and L.D. Foil, 2010. Evaluation of juvenile hormone analogues as rodent feed-through insecticides for control of immature phlebotomine sandflies. J. Med. Entomol.
CrossRefDirect Link - Vinayagam, A., N. Senthilkumar and A. Umamaheswari, 2008. Larvicidal activity of some medicinal plant extracts against malaria vector Anopheles stephensi. Res. J. Parasitol., 3: 50-58.
CrossRefDirect Link - Shonouda, M.L. and S.M. Mehanney, 2000. New botanical derivatives, used in medicinal preparations, showing bioactive action on insect pests II: Effect on the vectors Culex pipiens and Musca domestica larvae. Pak. J. Biol. Sci., 3: 1039-1041.
CrossRefDirect Link - Chakkaravarthy, V.M., T. Ambrose, S. Vincent, R. Arunachalam, M.G. Paulraj, S. Ignacimuthu and G. Annadurai, 2011. Bioefficacy of Azadirachta indica (A. juss) and Datura metel (Linn.) leaves extracts in controlling Culex quinquefasciatus (Diptera: Culicidae). J. Entomol., 8: 191-197.
CrossRefDirect Link - Katooli, N., E.M. Moghadam, A. Taheri and S. Nasrollahnejad, 2010. Management of root-knot nematode (Meloidogyne incognita) on cucumber with the extract and oil of nematicidal plants. Int. J. Agric. Res., 5: 582-586.
CrossRefDirect Link - Mariita, R.M., C.K.P.O. Ogol, N.O. Oguge and P.O. Okemo, 2011. Methanol extract of three medicinal plants from samburu in northern kenya show significant antimycobacterial, antibacterial and antifungal properties. Res. J. Med. Plant, 5: 54-64.
CrossRefDirect Link - Alvar, J., S. Croft and P. Olliaro, 2006. Chemotherapy in the treatment and control of leishmaniasis. Adv. Parasitol., 61: 223-274.
CrossRef - Tonui, W.K., 2006. Situational analysis of leishmaniases research in Kenya. Afr. J. Health Sci., 13: 7-21.
PubMedDirect Link - Rassi, Y., E. Javadian, A. Nadim, A. Zahraii and H. Vatandoost et al., 2005. Phlebotpmus (Larroussius) kandelakii the principal and proven vector of visceral leishmaniasis in North West of Iran. Pak. J. Biol. Sci., 8: 1802-1806.
CrossRefDirect Link - Ikbal, C., B.H. Mounia and B.H. Habib, 2007. Toxicity experiments of the saponic extract of Cestrum Parqui (Solanaceae) on some insect spices. J. Entomol., 4: 113-120.
CrossRefDirect Link