
Bo Luo
Department of Biochemistry, Luzhou Medical College, Luzhou 646000, Sichuan, China
Huailin Xiong
Department of Human Anatomy, Luzhou Medical College, Luzhou 646000, Sichuan, China
Yingyu Mao
Department of Pathogenic Biology, Luzhou Medical College, Luzhou 646000, Sichuan, China
Haiqing Yu
Research Center for Preclinical Medicine, Luzhou Medical College, Luzhou 646000, Sichuan, China
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 4 | Page No.: 258-262







ABSTRACT
Currently, it is obscure whether Toll-like Receptor 2 (TLR2) is related to the pathogenesis and pathophysiology of Paederus Dermatitis (PD). This study focused on the analysis of TLR2 mRNA expression in spleen of PD mice. Pederin was extracted at -20°C by dipping six female Paederus fuscipes Curtis in 1 mL of 70% ethanol for one month. Mouse abdominal hairs were denuded gently using 4% Na2S. By evenly extending 20 μL pederin solutions in 8 mm diameter skin, PD were induced in mice. Erythema and edema symptoms were scored. At 44 h after induction, the expression of TLR2 mRNA in spleen was analyzed using a reverse transcription and semi-quantitative polymerase chain reaction (SQ-PCR) with GAPDH mRNA as a reference. Normal mice with unhaired abdominal skin were used as controls. In this study, erythema and edema symptoms were observable, especially at 20 h after pederin application. The relative quantity of TLR2/GAPDH mRNA was 1.32±0.213 and 0.896±0.036 in healthy group and PD group, respectively. The expression level of TLR2 mRNA in spleen was significantly decreased (p<0.05) after treatment with pederin. These results suggested that pederin is a Th1 (T helper cell 1) inducing agent and there are Th1 cytokines in PD mice in early stage after contact, however, down-regulation of TLR2 mRNA expression is most likely related to Th2 (T helper cell 2) cytokines. The pathogenesis and pathophysiology In PD mice are closely associated with Th1 and Th2 responses, especially with cytokines excretion. This hypothesis needs to be investigated further.
PDF Abstract XML References Citation
Received: April 04, 2012;
Accepted: May 19, 2012;
Published: July 04, 2012
How to cite this article
Bo Luo, Huailin Xiong, Yingyu Mao and Haiqing Yu, 2012. Expression of Toll-Like Receptor 2 mRNA in Spleens of Paederus Dermatitis Mice. Journal of Biological Sciences, 12: 258-262.
DOI: 10.3923/jbs.2012.258.262
URL: https://scialert.net/abstract/?doi=jbs.2012.258.262
DOI: 10.3923/jbs.2012.258.262
URL: https://scialert.net/abstract/?doi=jbs.2012.258.262
INTRODUCTION
Paederus Dermatitis (PD) is an irritant contact dermatitis provoked by beetles in genus Paederus (Coleoptera: Staphylinidae). Pederin (C25H45O9N) released from the abdomen, especially in female insect, is an effective vesicant, often leading to a PD outbreak with initial linear erythema in skin after contact (Wang et al., 2004; Mammino, 2011). Also, this amide is easily dissolved in ethanol (Liu and Liang, 2003; Liu et al., 2004; Singh and Yousuf, 2007). The species commonly causing PD are Paederus fuscipes Curtis in China (Yang et al., 2001; Wang et al., 2004). P. fuscipes beetles are nocturnal and attracted by incandescent and fluorescent lights and it has an active climax in September which is identical to the peak time of PD presentation (Liu et al., 2004). Without biting or stinging, the beetle releases coelomic fluid immediately when it was accidentally brushed against or crushed over the skin. The potent vesicant provokes a reaction on the skin about 24 h after contact (Mammino, 2011). Therefore, it is difficult to put prevention of this inflammatory disease if one’s skin is exposed.
Toll-like receptor 2 (TLR2) is a transmembrane receptor with an extracellular domain and an intracellular signaling domain (Jarrousse et al., 2006). By recognizing a group of microbial-derived molecules which is also called pathogen-associated molecular patterns (PAMPs), it plays a crucial role in activation of the innate immune response (Zhang and Mao, 2011). TLR2 triggers the MYD88 dependent pathway and involve the early phase of NF-κB activation which induced a variety of inflammatory cytokines (Kim, 2005). It is expressed widely in monocytes, macrophages and dendritic cells and is one most important factor in linking innate and adaptive immunity (Pasare and Medzhitov, 2004). TLR2 is closely related to inflammatory skin disease (Jarrousse et al., 2006; Zhang and Mao, 2011; Kaplan et al., 2012),however, we still know very little about the expression pattern and its effects on pathogenesis and pathophysiology of PD.
Semi-quantitative polymerase chain reaction (SQ-PCR) has been shown to be useful in mRNA expression analysis (Mekki et al., 2006; Kavousi et al., 2009; Valiellahi et al., 2009; Markova et al., 2010; Ngamniyom et al., 2011). The aim of this study was to investigate the expression level of TLR2 and discuss its molecular mechanisms involved in the progression of PD. To this end, TLR2 gene expression was detected in spleens of PD mice.
MATERIALS AND METHODS
The study was conducted from November 2011 to February 2012 at Luzhou city, Research Center for Preclinical Medicine of Luzhou Medical College.
Beetle materials and pederin preparation: P. fuscipes beetles used in this study were collected under fluorescent lights at night from Luzhou city of Sichuan province. Adult P. fuscipes were carefully identified by Dr. Haiqing Yu. The voucher specimens (accession number PFTY110901) have been deposited at the Medicinal Botanical Association of Zhongshan Mountain (MBAZM), Luzhou Medical College. Sex identification was followed by descriptions of Liu et al., (2004). Pederin was extracted in 70% ethanol as suggested by Wang et al. (2004). In this study, six female P. fuscipes were dipped in 1 mL 70% ethanol at -20°C for one month. Subsequently, beetle bodies were gently comminuted by adding quartz sands. After centrifugation at 12000 rpm for 5 min at room temperature, supernatant was transferred to a new 1.5 mL tube and was stored at -20°C for future use.
Experimental mice and PD model: Adult (8-10 weeks old) male KM mice (Kunming, Outbred Mice, SPF grade) were purchased from experimental animal center of Luzhou Medical College. Water and food were provided ad libitum. Abdominal skin was denuded gently using 4% Na2S as suggested by Tie et al. (2009). Mice were raised two days after denudation and those who had intact skin were used. Eight mice were divided randomly into two groups: control group (n = 3) and PD group (n = 5). PD mice were induced by evenly extending 20 μL pederin solutions on abdominal skin of 8 mm in diameter. The observable skin reaction was read at 14, 20, 38 and 44 h after application and valued according to Draize’s scale (Draize, 1959) using double-blind trial. At 44 h after treatment with pederin, spleens were isolated for RNA extraction.
Total RNA extraction and cDNA synthesis: Total RNA was extracted using the RNAsimple Total RNA Kit (TIANGEN BIOTECH, Co., Ltd, Beijing, China). Fresh spleens (about 100 mg) were put into a 1.5 mL RNase-free tube which contained 0.5 mL lysis buffer RZ and was ground with 1 mL RNase-free pipette tip on ice immediately. The subsequent protocol was slightly modified by shortening centrifugation time and ignoring all optional or repetitive steps. The concentration and purity of total RNA were detected immediately using ND-1000 (NanoDrop Technologies, USA). cDNA was synthesized quickly using BioBRK RT Kit (BioBRK Co., Ltd. Chengdu, China). According to the manufacturer’s instruction, reverse transcription was performed in a total volume of 20 μL containing 1 μg RNA, 2.5 μM of Oligo (dT) 18, 1xRT buffer, 1 mM of each dNTP, 100 unit of ReverTra Ace, 5 unit of Super-RI, 0.5 μL of RT-Enhancer and RNase free water to the final volume. The reaction mixture was incubated in 42°C for 40 min, followed by 5 min at 99°C to inactivate the enzyme. Synthesized cDNAs were diluted five times with sterile water and stored in -20°C until use.
Semi-quantitative PCR: To avoid DNA contaminants, forward primer (5’-AAACAACTTACCGAAACCTCAG-3’) and reverse primer (5’-AAGACTGCCCAGAGAATAA AAG-3’) of TLR2 were designed from two adjacent exons respectively using DNAMAN (Lynnon Biosoft, version 5.2.9 Demo) based on GenBank data (NM_011905.3). The expected TLR2 product is 350 bp in size. GAPDH is used as control and its control primers F/R (forward primer: 5’-ACCACAGTCCATGCCATCAC-3’ and reverse primer: 5’-TCCACCACCCTGTTGCTGTA-3’) generate 450 bp amplicon (BioBRK). The PCR reaction was carried out in a total volume of 20 μL containing 1xTaq PCR MasterMix (BioBRK), 0.5 μM of each primer, 8 μL diluted cDNA and sterile water to the final volume. Simultaneous, PCR reactions for TLR2 and GAPDH were carried out in a Mastercycler 5331 (Eppendorf, Germany) in different tubes and PCR cycle numbers were optimized respectively to evade the plateau. The reaction cycling condition consisted of an initial denaturation at 94°C for 5 min, followed by optimized cycles of 30 sec at 94°C, 30 sec at 54°C, 30 sec at 72°C and final extension of 10 min at 72°C.
Data analysis: The amplified PCR products were resolved by electrophoresis on 1.5% agarose gel in 1xTAE buffer. Gels were visualized by 0.5 μg mL-1 ethidium bromide staining and the images were documented using the ChemiDoc XRS (Bio-Rad, USA). Based on trace tracking, amplified bands were quantified with Quantity One 4.4 (Bio-Rad, USA). Lanes were subtracted background and width of band detection was artificially corrected. TLR2 and GAPDH trace qty ratio was used to calculate the relative mRNA expression levels. The t-test was used to compare the statistic significance of relative quantity between control group and PD group.
RESULTS
Grading values were marked and mean scores for mice in PD group after application were listed in Table 1. Visible erythema and edema were recorded at 14 h. The most serious symptoms were observed at 20 h and then skin reactions were eased at 38 and 44 h.
PCR amplifications of TLR2 and GAPDH were nearly in platform at 33 and 24 cycles (Fig. 1). Therefore, 32 and 23 cycle numbers for TLR2 and GAPDH amplifications were selected, respectively. PCR reactions of TLR2 and GAPDH generated expected amplicons which were used in relative quantity analysis (Fig. 2).
| Table 1: | Mean values for edema and erythema skin reactions in PD group mice |
 | |
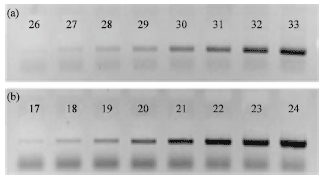 | |
| Fig. 1(a-b): | Optimization of PCR cycle numbers for (a) TLR2 and (b) GAPDH amplifications, Number in each lane are the cycle numbers |
 | |
| Fig. 2(a-b): | cDNA amplifications of (a) TLR2 and (b) GAPDH. M, DL2000 DNA ladder (bp), +: pederin treated, -: Pederin non-treated |
At 44 h after induction, the mRNA expression of TLR2/GAPDH was 1.32±0.213 and 0.896±0.036 in healthy group and PD group respectively. The levels of TLR2 mRNA in spleen were significantly decreased (p<0.05) after treatment (Fig. 3).
DISCUSSION
T helper cells (Th cells) play a crucial role in the immune system. Th1 and Th2 cells excrete cytokines, such as IFN-γ and IL4 which are inflammatory agents and anti-inflammatory agents respectively (Berger, 2000). Therefore, Th1 and Th2 responses are closely associated with inflammation. In this study, erythema and edema symptoms were observed, especially at 20 h after pederin application. This indicated that pederin was a Th1 inducing agent and there were Th1 cytokines in PD mice in early stage after contact.
In cultured human keratinocytes, Curry et al. (2003) revealed that TLR2 mRNA is not significantly enhanced or is decreased in some cases after treatment with different cytokines. Also, no immunoreactivity was detected for TLR2. Compared to normal skin, Jarrousse et al. (2006) found that the expression of TLR2 is not increased in keratinocytes of atopic dermatitis and psoriasis patients via immunohistochemistry detection. Similarly, expression level of TLR2 mRNA was significantly decreased in PD mice in the current study.
Hartgers et al. (2008) indicated that different pathogens might induce different regulation of TLR expression.
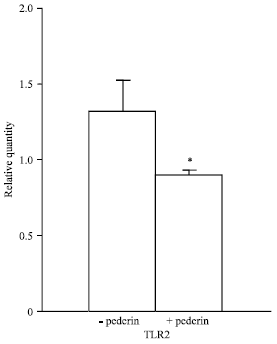 | |
| Fig. 3: | TLR2 expression level based on relative quantity of TLR2/GAPDH, *TLR2 mRNA was significantly down-regulated after pederin |
Mueller et al. (2006) showed that Th2 cytokines down-regulate TLR expression and function in human intestinal epithelial cells. This suggested that down-regulation of TLR2 mRNA at 44 h after pederin treatment is most likely related to the increase of Th2 cytokines. However, it needs to be investigated further.
Overall, pederin use is easy and effective to induce PD in mice and TLR2 expression is down-regulated in inflammatory skin diseases. The pathogenesis and pathophysiology in PD mice are closely associated with Th1 and Th2 responses, especially with cytokines excretion.
ACKNOWLEDGMENTS
The authors contribute equally to the present work. We are thankful to the Youth Foundation, the National Natural Science Pre-Research Foundation and the Scientific Research Starting Foundation for Doctors of Luzhou Medical College (No. 409, 676 and 606).
REFERENCES
- Berger, A., 2000. Th1 and Th2 responses: What are they? Br. Med. J., 321: 424-424.
CrossRefDirect Link - Curry, J.L., J.Z. Qin, B. Bonish, R. Carrick and P. Bacon, 2003. Innate immune-related receptors in normal and psoriatic skin. Arch. Pathol. Lab. Med., 127: 178-186.
PubMedDirect Link - Hartgers, F.C., B.B. Obeng, Y.C. Kruize, M. Duijvestein and A. de Breij, 2008. Lower expression of TLR2 and SOCS-3 is associated with Schistosoma haematobium infection and with lower risk for allergic reactivity in children living in a rural area in Ghana. PLoS Negl. Trop. Dis., 2: 227-227.
Direct Link - Jarrousse, V., G. Quereux, S. Marques-Briand, A.C. Knol and A. Khammari, 2006. Toll-like receptors 2, 4 and 9 expression in cutaneous T-cell lymphoma (mycosis fungoides and Sezary syndrome). Eur. J. Dermatol., 16: 636-641.
Direct Link - Kaplan, D.H., B.Z. Igyarto and A.A. Gaspari, 2012. Early immune events in the induction of allergic contact dermatitis. Nat. Rev. Immunol., 12: 114-124.
PubMedDirect Link - Kavousi, H.R., H. Marashi, J. Mozafari and A.R. Bagheri, 2009. Expression of phenylpropanoid pathway genes in chickpea defense against race 3 of Ascochyta rabiei. Plant Pathol. J., 8: 127-132.
CrossRefDirect Link - Kim, J., 2005. Review of the innate immune response in acne vulgaris: Activation of toll-like receptor 2 in acne triggers inflammatory cytokine responses. Dermatology, 211: 193-198.
CrossRefPubMedDirect Link - Liu, C.L., M.Y. Yu, Q. Zhao and L. Huang, 2004. Research on Paederus fuscipes curtis' regular life and human's dermatitis. Q. For. By-Prod. Specialty China, 5: 24-26.
Direct Link - Mammino, J.J., 2011. Paederus dermatitis: An outbreak on a medical mission boat in the Amazon. J. Clin. Aesthet. Dermatol., 4: 44-46.
PubMed - Markova, T., M. Tabuchi, B. Alexieva, E. Nikolova, Y. Aragane and H. Higashino, 2010. Rolipram inhibits phosphorylation and activation of ERK/MAP kinase signalling pathways in allergen-activated human peripheral mononuclear cells. Int. J. Pharmacol., 6: 600-607.
CrossRefDirect Link - Mekki, D.M., B. Liang-yong, W. Jin-yu, Y. Yan, L. lin-chuan, Y. Yabo and H.H. Musa, 2006. Differential expression levels assessment of avian E2A gene in two Chinese indigenous duck breeds (Anas platyrhynchos) by semi-quantitative RT-PCR. Int. J. Poult. Sci., 5: 895-899.
CrossRefDirect Link - Mueller, T., T. Terada, I.M. Rosenberg, O. Shibolet and D.K. Podolsky, 2006. Th2 cytokines down-regulate TLR expression and function in human intestinal epithelial cells. J. Immunol., 176: 5805-5814.
PubMed - Ngamniyom, A., W. Magtoon, Y. Nagahama and Y. Sasayama, 2011. Expression levels of bone morphogenetic protein 2b in fins of adult japanese medaka (Oryzias latipes) exposed to sex steroid hormones. J. Fish. Aquat. Sci., 6: 119-129.
CrossRefDirect Link - Singh, G. and A.S. Yousuf, 2007. Paederus dermatitis. Indian J. Dermatol. Venereol. Leprol., 73: 13-15.
CrossRefPubMedDirect Link - Valiellahi, E., A. Niazi and M. Farsi, 2009. Semiquantitative RT-PCR analysis to assess the expression levels of Wcor14 transcripts in winter-type wheat. Biotechnology, 8: 323-328.
CrossRef - Zhang, W.W. and X.H. Mao, 2011. Toll-like receptors and their role in pathogenesis and vaccine study of cutaneous diseases. J. Zhejiang Univ., 40: 222-229.
PubMedDirect Link