
Ali Vaziri Gohar
Department of Biochemistry, Afzalipour School of Medicine, Kerman University of Medical Sciences, Kerman, Iran
Abbas Mohammadi
Department of Biochemistry, Afzalipour School of Medicine, Kerman University of Medical Sciences, Kerman, Iran
Journal of Biological Sciences
Year: 2010 | Volume: 10 | Issue: 3 | Page No.: 200-208







ABSTRACT
The strategies for reduction of carcinogenic factors exposure can reduce the risk of most cancer types. Millions of people around the world get exposed to high levels of heavy metals in the drinking-water. Therefore, quality control in drinking-water and detection of its heavy metals is extremely critical issue. The carcinogenicity of aluminum, arsenic, chromium, nickel and selenium has been documented previously, but in a scattered fashion. Trace amount of these elements can induce genetic and epigenetic alterations in different cancer-related genes of somatic and stem cells, thus involved in cancer stem cell formation. Through epigenetic effects of these elements have more prominent role than their genetic effects, these elements are able to change the pattern of cancer-related genes expression profiles, too. Therefore, understanding the underlying epigenetically mechanisms of these trace elements can reduce their toxicities duo to individual and social preventive behaviors. It seems that the toxic effects of these elements in many regions are predictable, but antioxidant supplements eliminate the reactive oxygen species as leading effects of these elements.
PDF Abstract XML References Citation
Received: October 30, 2009;
Accepted: February 08, 2010;
Published: March 17, 2010
How to cite this article
Ali Vaziri Gohar and Abbas Mohammadi, 2010. Epigenetic Effects of Carcinogens. Journal of Biological Sciences, 10: 200-208.
DOI: 10.3923/jbs.2010.200.208
URL: https://scialert.net/abstract/?doi=jbs.2010.200.208
DOI: 10.3923/jbs.2010.200.208
URL: https://scialert.net/abstract/?doi=jbs.2010.200.208
INTRODUCTION
The importance of cancer prevention has led to detection and elimination of environmental carcinogenic agents. Many carcinogens have already been identified and the relevant information with regard to these agents is available. However, humans use many food and beverage items, assuming that they are safe. One example is the potentially harmful presence of heavy metals that can cause serious health problems. People may be exposed to heavy metals over the course of their lifetime. The heavy metals in drinking-water pose the greatest threat to public health in this regard. This necessitates setting suitable quality control procedures.
The main source of heavy metals in drinking-water is contamination of surface and ground waters by industrial sewage and agricultural run-off (Karavoltsos et al., 2008).
In the areas that the water distribution network is made of alloys containing heavy metals, some people may not afford bottled or mineral water with controlled heavy metal concentrations and they consume tap water, therefore the possibility of contamination of drinking-water with heavy metals greatly increases (Leivadara et al., 2008).
According to some WHO reports, the concentration of these elements in ground water is high in several countries including Bangladesh, India and Argentina (WHO, 2003).
Heavy metals in drinking-water are toxic and can easily enter the body. The genetic and epigenetic effects of these elements are associated with an increased risk of different cancer types (Bower et al., 2005).
Epigenetic mechanisms play an equally important if not a more prominent role than genetic events in carcinogenesis. These effects occur most frequently during the early stages of tumor development. Epigenetic events include reversible modifications of histone proteins and CpG islands of gene promoters that affect not only gene expression of germ and somatic cells, but also cause indirect gene-sequence changes (Jones and Baylin, 2002; Vaziri Gohar et al., 2007).
CpG islands (5'-CG-3' sequence) exist in about 40% of mammalian genes. Hypermethylation or hypomethylation of C5 position of the cytosine base are involved in the inhibition of expression of tumor suppressor genes or the increase of the oncogene expression, both of which contribute to cancer development and progression (Kinzler and Vogelstein, 1997).
Gene silencing can also take place through methylation of lysine 9 in histone-H3 (H3-K9), which results in a cascade of clustering of several proteins including HP1 protein, SUV39H1 histone methyltransferase, histone deacetylases, DNA Methyl transferases and finally methyl-C binding proteins (MBD) (Tamaru and Selker, 2001; Jackson et al., 2002; Vogelstein and Kinzler, 2004).
Methylated cytosine may be spontaneously deaminated to produce a thymine, resulting in a specific transition mutation in CpG islands, for example in the TP53 tumor suppressor gene as a guard of genome. In addition, hypermethylation of histone proteins causes changes in the chromatin configuration, predisposing cells to allelic loss at a specific locus in the chromosomes (Egger et al., 2004). Such genetic and epigenetic alterations in growth-control genes such as DNA-repair genes, tumor suppressor genes, oncogenes, apoptotic genes, combine to determine the cellular phenotype and differentiation (Vaziri Gohar et al., 2007; Mohammadi et al., 2008).
In ranking the carcinogens, heavy metals have been classified by the International Agency for Research on Cancer (IARC) and Environmental Protection Agency (EPA) as the first group, except for selenium that has been listed within group 3 (not carcinogen to humans) of the IARC classification (IARC, 1989).
The goal of this review was the introduction and comparison of epigenetic effects caused by the following elements in cancer-related genes in biological systems and show that the incidence of cancer can be reduced by adopting prevention behavior especially in terms of drinking-water.
Aluminum: The compounds of this element have a wide range of applications in different industries, including cosmetics and food additives (Laden and Felger, 1988).
Aluminum-induced carcinogenesis is related to its ability to bind to the estrogen receptor and mimic estrogen functions, therefore its named metalloestrogen (Darbre, 2005). Metalloestrogen triggers expression in genes that contain Estrogen Responsive-Element (ERE) on their promoters. In mammary gland cells, this gives rise to an increase in the number of divisions of breast cells, thus increasing replication errors in cancer-related genes (Sun et al., 2007).
It has been shown that if antiperspirants containing aluminum applied on the skin around the underarm and breast areas are not effectively washed, some aluminum salts remains in the area. This gives rise to continuous exposure and enhances the risk of breast cancer (Stellman et al., 2000).
There are two different groups of Estrogen Receptors (ER). The first group exist in cytosol/nucleus (ER-α and ER-β) and act as transcription factors by directly binding to ERE. The second group exists in plasma membrane as transmembrane G-protein coupled receptors. This kind of ERs can also regulate gene expression through interaction with other transcription factors, without directly binding to ERE. Plasma membrane located ER46 is involved in endothelial Nitric Oxide Synthase (eNOS) phosphorylation and rapid Nitric Oxide (NO) release via phosphatidyl inositol 3-kinase in endothelial cells. Another plasma membrane located ER family called ER66 regulates reporter gene expression (Darbre, 2005).
Aluminum can bind to both nuclear and membrane ERs and ERE; as a result, it can activate both ER signal transductions. Therefore, as expected, Al3+ treatment results in intracellular NO generation (Satoh et al., 2007).
In addition to breast carcinogenesis, estrogen can activate telomerase gene expression as a gene containing ERE, in ER-α positive cells, but not in ER negative cells, this results in endometrial cancer (Harley, 2008).
Epigenetic effects of aluminum take place through the binding of trivalent (Al3+) to the phosphate groups of double stranded DNA under physiologic pH, thus changing DNA topology from B to Z in (CCG)12 repeat regions (Zhang et al., 2002).
The expansion of the triplet repeats is named dynamic mutation and may be localized both in coding and non-coding regions. A minimum of 5-10 triplet-repeats increases the probability of hairpin formations, mainly in the lagging strand. Movement of DNA polymerase along the hairpin structure leads to replication slippage and genomic instability, causing deletion mutations. Expansion of more than 200 copies of these repeats leads to excessive methylation of cytosines in the promoter of FMR1 gene which results in fragile X syndrome (Lukusa and Fryns, 2008).
Arsenic: Arsenic is mostly known as an epigenetic carcinogen metalloid when in the form of an inorganic compound. Trivalent arsenite (As3+) has more carcinogenic properties than the pentavalent arsenate (As5+) (Patterson et al., 2003; Alkahtani, 2009). Trivalent arsenic can bind with high affinity to thiol groups of proteins and reduced glutathione (GSH) (Suzuki et al., 2004).
Long time uptake of drinking-water containing low levels of arsenite, induces carcinogenesis in skin, lung, bladder and kidney tissues, resulting from alteration in multiple signaling pathways (Jensen et al., 2009). The risk of bladder cancer in people drinking water with an arsenic level above 100 ppb increases over 15 times compared with people living in areas with less than 10 ppb (Vahter, 2002).
Arsenic is methylated for detoxification and excretion from the body. It is exactly this reaction that gives rise to the carcinogenic properties of arsenic through epigenetic transformations. This is contrary to the general belief that considers the methylation as a way for detoxification. The toxicity of monomethyl arsenic (MMAs) and dimethyl arsenic (DMAs) is more than arsenite (Patterson et al., 2003; Suzuki et al., 2004).
Arsenic methylation occurs by Glutathione S-Transferase (GST), arsenic III methyltransferase (AS3MT) and S-Adenosyl Methionine (SAM). These enzymes compete with DNA methyltransferase (DNMT) for DNA methylation, hence, inhibiting DNA methyltransferase indirectly and inducing the reactivation of silenced tumor suppressor genes (Huang, 2002).
Exposure to arsenic induces the ROS formation (through its reduction) as an unavoidable reaction of normal cell metabolism (Galaris et al., 2008).
ROS, acting as a second messenger, are involved in the activation of PI3K/Akt pathway and the subsequent induction of transcription factor hypoxia inducible factor-1 (HIF-1α) but not HIF-1β and Vascular Endothelial Growth Factor (VEGF) induction (Gao et al., 2004; Galanis et al., 2008).
Another important mechanism of arsenic induced carcinogenesis is through enhancing the genotoxicity of other carcinogens, including ultraviolet radiation (UVR), ionizing radiation, alkylating agents, or oxidants. UVR induces non-melanoma skin cancer. Strands of DNA exposed to photons of UVA and UVB break and Cyclobutane Pyrimidine Dimers (CPDs) are formed (Ravanat et al., 2001; Melnikova and Ananthaswamy, 2005).
UVRs can activate a zinc-finger protein family poly (ADP-ribose) polymerase (PARP), particularly one member of this family named PARP-1, has an important role in the regulation of Nucleotide Excision Repair (NER). CPDs have been identified in p53 and PTCH tumor suppressor genes and ras oncogenes (Rossman et al., 2001, 2002).
Arsenite stimulates inducible Nitric Oxide Synthase (iNOS) expression and NO production through mammalian mitogen-activated protein kinases p38 and activation of nuclear transcription factor-kappa B (NF-κB) (Ding et al., 2008).
Between 40-60% of arsenic intake is excreted into the urine (Fujihara et al., 2009). A major proportion of urinary arsenic is composed of dimethylated arsenic (Vahter, 2000).
Chromium: Trivalent chromium is an epigenetic carcinogen factor since, it can form stable compounds with macromolecules such as DNA and cysteine residue of proteins and glutathione (Zhitkovich et al., 1995).
The trivalent form of chromium cannot pass the cell membrane; however, the hexavalent salts are able to enter the cell and are converted to the trivalent form (Alexander and Aaseth, 1995).
Thus, depending on the situation, reducing agents can affect carcinogenic properties of chromium and inside the cell, chromium (VI) can be converted to a carcinogen. During Cr (VI) reduction, many compounds such as oxygen radicals, DNA Inter-strand Cross Links (ICLs) and Single-Strand Breaks (SSBs) may form.Inter-strand Cross Links act as physical barriers to DNA replication and transcription events, thus inducing apoptosis (Schnekenburger et al., 2007).
The chromium carcinogenicity, particularly in lung epithelial cells and fibroblasts, is imposed through hypermethylation of CYP1a1 promoter. Chromium recruits histone deacetylase 1 (HDAC1) and DNMT1, especially to CYP1a1 promoter and this assembly recruits BP1 and inhibits CYP1a1 gene expression (Wei et al., 2004).
The CYP1A1 is important in the metabolism of carcinogens such as Polycyclic Aromatic Hydrocarbons (PAHs) and heterocyclic amines that are widely distributed widely in our environment through automobile exhausts, cigarette smoke, charcoal-broiled cooking and industrial waste. In contrast to other cytochrome P450 enzymes such as epoxide hydrolase and dihydrodiol dehydrogenase that are involved in PAH and Benzo(a)pyrene-induced carcinogenesis, CYP1A1 inhibits PAH carcinogenesis. Thus, inhibition of CYP1A1 by chromium leads to the production of a PAH (Wu et al., 2008).
PAHs have an important role in the activation of cytosolic ligand-activated transcription factor named Aromatic hydrocarbon Receptor (AhR) (Nebert et al., 2000). After formation, the PAH-AhR complex transfer into the nucleus. In the nucleus, PAH is detached from the complex and AhR binds to its nuclear partner, Arnt. This new complex acts as a transcription factor and interacts with DRE of CYP1A1 gene, leading to the activation of CYP1A1 gene expression, thus causing bioactivation of exogenous procarcinogens of both hepatocellular and lung carcinomas (Li et al., 2009).
It is interesting that PAH through binding to transcription factor AhR, activates CYP1a1 gene expression and CYP1A1 inhibits PAH carcinogenesis, but in the presence of Cr, the promoter of CYP1a1 is inactivated and PAH can act as carcinogens (Wei et al., 2004).
Benzo(α)pyrene is also a member of Polycyclic Aromatic Hydrocarbon (PAHs) family that is metabolically transformed from its pro-carcinogenic status to the carcinogenic metabolite (BP-7,8-dihydrodiol-9,10-epoxide (BPDE)), that can binds covalently to DNA and forms BPDE-DNA adduct and ROS. The BPDE activates apoptosis through p53 independent and dependent manner (Drukteinis et al., 2005).
P53 dependent Cr-induced apoptosis takes place by increasing p53 phosphorylation at Ser392, as well as up-regulation of pro-apoptotic gene bcl-XS and caspase-7 and down-regulation of several anti-apoptotic genes from Bcl2-family (bcl-W and bcl-XL) and bax. These apoptotic events result in the destruction of the mitochondria and release of cytochrome c (Blankenship et al., 1994; Carlisle et al., 2000; Ceryak et al., 2004). Moreover, Cr induces the ATM protein production, which phosphorylates and activates Chk2 protein. The phosphorylated Chk2 in turn phosphorylates and activates p53. The phosphorylated p53 does not bind to MDM2 protein (Ha et al., 2003; Vaziri Gohar et al., 2007).
Cr exposure at very high concentrations activates all subclasses of MAPK through phosphorylation; therefore, Cr acts as a MAPK kinase and increases survival/proliferation in a dose-dependent manner. This function is associated with its ability in ROS generation (Thomas, 1992; Gao et al., 2002).
Nickel: Water-insoluble nickel compounds including nickel sulfides, disulfides and oxides readily enter the cell and are very potent carcinogens (Dunnick et al., 1995). In contrast, water-soluble nickel compounds including acetate, chloride, nitrate and sulfate do not enter the cells as readily as water-insoluble nickel compounds (Abbracchio et al., 1982).
The increase in the usage of nickel compounds and the spread of nickel due to its dissolution from nickel ore-bearing rocks are the main causes of nickel presence in the environment. The primary source of nickel in drinking-water is the leaching of metals in water network (WHO, 2005). However, food is the major source of nickel exposure in the non-smoking, non-occupationally exposed population, but nickel absorption from water was significantly higher than absorption of nickel from beverages like tea, coffee, or orange juice and milk (Nielsen et al., 1999).
Ni2+ induces carcinogenesis through several processes including DNA hypermethylation, DNMT inhibition, DNA mutation, ROS generation, inhibiting histone H2A, H2B, H3 and H4 acetylation, converting the tumor suppressor genes to the heterochromatin and substantial increases of the ubiquitination of H2A and H2B (Ke et al., 2008). Therefore, nickel plays an important role in silencing of genes (Lee et al., 1998).
Nickel binds to DNA in different positions. It binds to phosphate backbone of DNA in place of Mg and promotes the conversion of suppressor genes to the heterochromatin (Cangul et al., 2002). Moreover, it’s binding to histone H4 leads to the inhibition of lysine acetylation and subsequently DNA hypermethylation (Broday et al., 2000).
These events play an important role in silencing of tumor suppressor genes and the other genes that are involved in carcinogenesis pathways.
Selenium: Selenium is an essential trace element with a narrow range between toxic and therapeutic doses; its activity is therefore highly dose dependent. Enzymes containing selenium such as glutathione peroxidase like other antioxidant elements can protect body from oxidative damage and reduce the risk of cancer incidence and mortality through several pathways such as apoptosis and alteration of some collagen types (Rayman, 2000).
Since selenium, like arsenic is detoxified by methylation through S-adenosylmethionine pathway, competition between arsenic, selenium and DNMT1 for methyl donated by S-adenosylmethionine leads to DNA hypomethylation and an increase in arsenic retention in tissues (Xiang et al., 2008).
Organic selenium compounds such as selenomethionine, Se-methyl-selenocysteine (Se-MSC) and particularly Selenocystine (SeC), have shown more anticarcinogenic activity than inorganic compounds in lung cancer model systems. However, in contrast to selenomethionine, selenocystine decreases cellular reduced thiol agents like N-acetylcysteine (NAC) and GSH, thus increasing the ROS formation (Zou et al., 2008). Selenium-containing proteins can induce apoptosis pathway through caspase activation. But, selenite, SeC and selenomethionine mostly activate apoptosis by caspase-independent pathways through p53 activation and anti-apoptotic inactivation and release of cytochrome c from mitochondria as follows. First, these compounds increase production of reactive oxygen species. ROS-mediated modified products such as DSBs, are detected by ATM and ATR proteins, which in turn can activate p53 in MCF-7 human breast cancer cells and human prostate cancer. These DSBs can even synergistically increase the intracellular ROS production. Second, they induce p53 phosphorylation at Ser15, Ser20 and Ser392 residues, thus decreasing p53-MDM2 protein interaction and p53 stability (Chen and Wong, 2008).
The Se-MSC shows its anticarcinogenic activity through down regulation of some extracellular matrix proteins such as collagen type I alpha 1 (COL1A1), COL1A2 and COL7A1 and up-regulation of COL6A1 and COL4A5 genes in human prostate cell line (Hurst et al., 2008).
DISCUSSION
Heavy metal-induced carcinogenesis is the cause of death in some countries with concentrations greater than global standard levels. Also, analysis of heavy metals in archival Formalin-Fixed Paraffin-Embedded (FFPE) tissue specimens showed the possible roles of these elements in cancer development, so detecting and removal of heavy metal from tap and bottled drinking-waters needs special attention (Asadikaram et al., 2008; Sarafanov et al., 2008; Vaziri Gohar and Mohammadi, 2009).
In most populations, the main source of heavy metal entrance to the body may not be drinking water, but this is an important route (WHO, 2003).
In spite of the important role of heavy metals in the deviation of cell growth toward carcinogenesis, some of these elements such as selenium, arsenite and chromium have anticarcinogenic effects.
Selenium containing compounds that serve as antioxidants have an important role in prevention and attenuation of toxic effects of other heavy metals and their deficiency leads to Keshan and Kaschin-Beck diseases (Rayman, 2000). Using 15 mg kg-1 sodium selenite fortified-salt for 6 years resulted in decrease of hepatocellular carcinoma (HCC) incidence in humans (Yu et al., 1999).
Arsenite is involved in the treatment of Acute Promyelocytic Leukaemia (APL) and hematological malignancies (Bachleitner-Hofmann et al., 2002). Cr-exposure causes reduction of the proliferative ability of human lymphoblast cell lines (Zhang et al., 2002). Chromium also has an application in individuals with impaired glucose tolerance. SeC and chromium are effective against lung tumors induced by a compound named 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) (Kasprzak and Buzard, 2000).
Figure 1 shows a summary of cellular events which are attached be heavy metals. The major DNA lesions associated with heavy metal exposure result from production of ROS, DNA base damage, strand breaks, apurinic sites and cross-links (Kasprzak and Buzard, 2000). The ROS are products of heavy metal reduction. Thus, the reducing conditions like acidic condition in the stomach, extracellular ascorbic acid and GST activity can be involved in heavy metal-induced carcinogenesis.
ROS induce a heterodimeric helix-loop-helix transcription factor HIF-1 (Imamura et al., 2009). HIF-1 expression gives rise to resistance to chemotherapy and radiotherapy. The expression of HIF occurs in several human cancers, such as prostate and Sporadic Clear Cell Renal Cell Carcinoma (SCRCC). Also, HIF production leads to up-regulation of VEGF and thus, tumor angiogenesis (Gordan et al., 2008).
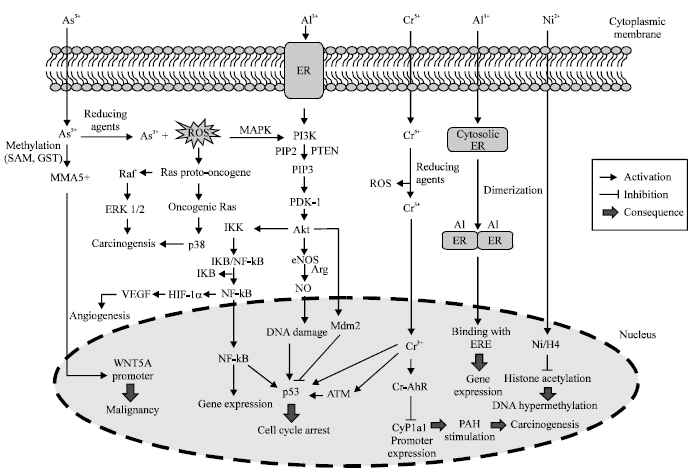 | |
| Fig. 1: | Epigenetic pathway of heavy metals in human. Schematic representation of heavy metal signaling pathways which lead to carcinogenesis. Individual pathways are discussed in the text (Cowell, 2001; Vaziri Gohar et al., 2007) |
The Von Hippel-Lindau tumor suppressor protein (VHL) inhibits VEGF formation by inhibition of hypoxia-inducible factors. Therefore, loss of VHL tumor suppressor gene function promotes tumor cell invasion, which has an important role in familial and most sporadic clear cell renal cell carcinoma and prostate tumorigenesis (Monsef et al., 2007).
Besides ROS, heavy metals can induce NO production. NO exposure can alter and damage DNA molecule, thus promoting p53 post-translation and accumulation. Therefore, as expected, inhibition of iNOS gene activity is involved in heavy metal-induced carcinogenesis (Hussain et al., 2003).
ROS also can act as mediators of NF-kB activation (Monsef et al., 2007). This transcription factor is involved in carcinogenesis by inhibition of the apoptosis pathway and also can induce cyclin D1 gene (Guttridge et al., 1999). Cyclin-D1 is one of the important targets that has underlying role in the estradiol and metalloestrogen-induced proliferations. Binding of Aluminum as a metalloestrogen to ERs has different effects on cyclin-D1 expression, e.g. binding to ER-α increase and binding to ER-β represses the expression level. Activation of cyclin-D1 as a protein of G1 phase of cell cycle followed by cyclin-E leads to the inactivation of pRB1 via phosphorylation, thus permits the cell to enter the S phase of the cell cycle (Guttridge et al., 1999; Leader et al., 2006).
Exposure to low level of MMAs3+ induces malignancy through irreversible epigenetic activation of a member of the non-canonical WNT signaling pathway, named WNT5A tumor suppressor gene. Therefore, this arsenic form mediates the activation of PKC, hence alteration of the intracellular calcium levels. WNT5A gene has a tumor suppressive function, but up-regulation of this gene is involved in stomach, pancreas, bladder, melanoma and glioblastoma malignancies. The activation of WNT5A transcription correlates with the enrichment of permissive histone modifications and the reduction of repressive modifications in the WNT5A promoter region (Jensen et al., 2009).
One enzyme family involved in heavy metal metabolism is cysteine-rich enzymes called metallothioneins (MTs). This enzyme family as an antioxidant, act as a scavenger of heavy metals and ROS. It is interesting that besides zinc, cadmium can also induce expression of this enzyme (Majumder et al., 2006). Another enzyme family with dual-functional is GST that in one side, can methylate and detoxify heavy metals through faster excretion, on the other side they may accelerate the reduction of these elements and increase their carcinogenicities (Adams et al., 2002).
Another mechanism that these elements affect the body is through their effects on stem cells. Stem cells with long lifetime such as embryonal, germinal and progenitor cells have a high opportunity for epigenetic and genetic accumulations. Epigenetic alterations of growth-control genes in the stem cells change the cellular differentiation. Therefore, heavy metal exposures in pregnancy, especially through drinking-water can transfer the metals to the fetus and make instable stem cells that have been defined as cancer stem cells which can result in abnormal cell expansion in the precancerous stage of tumor development (Sell, 2004). These cancer stem cells have first been detected in transmissible mice leukemia and have been reported in several types of malignancies such as brain, breast, colon and melanoma (Zhang and Rosen, 2006).
CONCLUSION
The heavy metals ability in ROS and NF-kB production and also human genetic differences through polymorphisms in glutathione S-transferase, metallothioneins and methyltransferase genes induce carcinogenesis. Many of these results may eliminate by daily antioxidant usage (Evans, 2008). The recommended dose of intake for adults by Recommended Dietary Allowance (RDA) for zinc, chromium and selenium is 11 mg, 25 and 55 μg.
This review shows that these heavy metals are carcinogen, particularly epigenetic carcinogen and might be solely responsible for tumors presentation and progression. The importance of these elements in cancer stem cell development give rise to the understanding other roles of these carcinogens and detection of possible removal drinking-water heavy metals by future research.
REFERENCES
- Abbracchio, M.P., J.D. Heck and M. Costa, 1982. The phagocytosis and transforming activity of crystalline metal sulfide particles are related to their negative surface charge. Carcinogenesis, 3: 175-180.
Direct Link - Adams, T.K., N. Saydam, F. Steiner, W. Schaffner and J.H. Freedman, 2002. Activation of gene expression by metal-responsive signal transduction pathways. Environ. Health Perspect., 110: 813-817.
Direct Link - Alexander, J. and J. Aaseth, 1995. Uptake of chromate in human red blood cells and isolated rat liver cells: The role of the anion carrier. Analyst, 120: 931-933.
Direct Link - Alkahtani, S., 2009. Antioxidation and hypomethylation effects on genotoxicity and programmed cell death induced in mice somatic cells by arsenic trioxide. J. Biol. Sci., 9: 721-729.
CrossRefDirect Link - Bachleitner-Hofmann, T., M. Kees and H. Gisslinger, 2002. Arsenic trioxide: Acute promyelocytic leukemia and beyond. Leuk. Lymphoma, 43: 1535-1540.
PubMed - Blankenship, L.J., F.C. Manning, J.M. Orenstein and S.R. Patierno, 1994. Apoptosis is the mode of cell death caused by carcinogenic chromium. Toxicol. Appl. Pharmacol., 126: 75-83.
PubMed - Bower, J., S. Leonard and X. Shi, 2005. Conference overview: Molecular mechanisms of metal toxicity and carcinogenesis. Mol. Cell Biochem., 279: 3-15.
CrossRef - Broday, L., W. Peng, M.H. Kuo, K. Salnikow and M. Zoroddu et al., 2000. Nickel compounds are novel inhibitors of histone H4 acetylation. Cancer Res., 60: 238-241.
Direct Link - Cangul, H., L. Broday, K. Salnikow, J. Sutherland and W. Peng et al., 2002. Molecular mechanisms of nickel carcinogenesis. Toxicol. Lett., 127: 69-75.
CrossRef - Carlisle, D.L., D.E. Pritchard, J. Singh, B.M. Owens, L.J. Blankenship, J.M. Orenstei and S.R. Patierno, 2000. Apoptosis and P53 induction in human lung fibroblasts exposed to chromium(VI): Effect of ascorbate and tocopherol. Toxicol. Sci., 55: 60-68.
Direct Link - Chen, T. and Y.S. Wong, 2008. Selenocystine induces caspase-independent apoptosis in MCF-7 human breast carcinoma cells with involvement of p53 phosphorylation and reactive oxygen species generation. Int. J. Biochem. Cell Biol., 41: 666-676.
PubMed - Darbre, P.D., 2005. Aluminium, antiperspirants and breast cancer. J. Inorganic Biochem., 99: 1912-1919.
PubMed - Ding, W., L.G. Hudso, X. Sun, C. Feng and K.J. Liu, 2008. As(III) inhibits ultraviolet radiation-induced cyclobutane pyrimidine dimer repair via generation of nitric oxide in human keratinocytes. Free Radiat. Biol. Med., 45: 1065-1072.
CrossRef - Drukteinis, J.S., T. Medrano, E.A. Ablordeppey, J.M. Kitzman and K.T. Shiverick, 2005. Benzo[a]pyrene, but Not 2,3,7,8-TCDD, Induces G2/M Cell Cycle Arrest, p21CIP1 and p53 Phosphorylation in human choriocarcinoma JEG-3 Cells: A distinct signaling pathway. Placenta, 26: S87-S95.
Direct Link - Dunnick, J.K., M.R. Elwell, A.E. Radovsky, J.M. Benson and F.F. Hahn et al., 1995. Comparative carcinogenic effects of nickel subsulfide, nickel oxide, or nickel sulfate hexahydrate chronic exposures in the lung. Cancer Res., 55: 5251-5256.
Direct Link - Egger, G., G. Liang, A. Aparicio and P.A. Jones, 2004. Epigenetics in human disease and prospects for epigenetic therapy. Nature, 429: 457-463.
PubMed - Evans, J., 2008. Antioxidant supplements to prevent or slow down the progression of AMD: A systematic review and meta-analysis. Eye, 22: 751-760.
PubMed - Fujihara, J., Y. Fujii, T. Agusa, T. Kunito, T. Yasuda, T. Moritani and H. Takeshita, 2009. Ethnic differences in five intronic polymorphisms associated with arsenic metabolism within human arsenic (+3 oxidation state) methyltransferase (AS3MT) gene. Toxicol. Applied Pharmacol., 234: 41-46.
CrossRef - Galanis, A., A. Karapetsas and R. Sandaltzopoulos, 2008. Metal induced carcinogenesis, oxidative stress and hypoxia signaling. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis, 674: 31-35.
CrossRef - Galaris, D., V. Skiada and A. Barbouti, 2008. Redox signaling and cancer: The role of labile iron. Cancer Lett., 266: 21-29.
Direct Link - Gao, N., L. Shen, Z. Zhang, S.S. Leonard and H. He et al., 2004. Arsenite induces HIF-1α and VEGF through PI3K, Akt and reactive oxygen species in DU145 human prostate carcinoma cells. Mol. Cell Biochem., 255: 33-45.
Direct Link - Gao, N., B.H. Jiang, S.S. Leonard, L. Corum and Z. Zhang et al., 2002. p38 signaling-mediated hypoxia-inducible factor 1α and vascular endothelial growth factor induction by Cr(VI) in DU145 human prostate carcinoma cells. J. Biol. Chem., 277: 45041-45048.
Direct Link - Gordan, J.D., P. Lal, V.R. Dondeti, R. Letrero and K.N. Parekh et al., 2008. HIF-alpha effects on c-Myc distinguish two subtypes of sporadic VHL-deficient clear cell renal carcinoma. Cancer Cell, 14: 435-446.
PubMed - Guttridge, D.C., C. Albanese, J.Y. Reuther, R.G. Pestell and A.S. Baldwin Jr., 1999. NF-kappaB controls cell growth and differentiation through transcriptional regulation of cyclin D1. Mol. Cell Biol., 19: 5785-5799.
PubMed - Ha, L., S. Ceryak and S.R. Patierno, 2003. Chromium (VI) activates ATM: Requirement of ATM for both apoptosis and recovery from terminal growth arrest. J. Biol. Chem., 278: 17885-17894.
Direct Link - Huang, S., 2002. Histone methyltransferases, diet nutrients and tumour suppressors. Nat. Rev. Cancer, 2: 469-476.
PubMed - Hurst, R., R.M. Elliott, A.J. Goldson and S.J. Fairweather-Tait, 2008. Se-methylselenocysteine alters collagen gene and protein expression in human prostate cells. Cancer Lett., 269: 117-126.
Direct Link - Hussain, S.P., L.J. Hofseth and C.C. Harris, 2003. Radical causes of cancer. Nat. Rev. Cancer, 3: 276-285.
CrossRefDirect Link - Imamura, T., H. Kikuchi, M.T. Herraiz, D.Y. Park and Y. Mizukami et al., 2009. HIF-1alpha and HIF-2alpha have divergent roles in colon cancer. Int. J. Cancer, 124: 763-771.
PubMed - Jackson, J.P., A.M. Lindroth, X. Cao and S.E. Jacobsen, 2002. Control of CpNpG DNA methylation by the kryptonite histone H3 methyltransferase. Nature, 416: 556-560.
PubMed - Jensen, T.J., R.J. Wozniak, K.E. Eblin, S.M. Wnek, A.J. Gandolfi and B.W. Futscher, 2009. Epigenetic mediated transcriptional activation of WNT5A participates in arsenical-associated malignant transformation. Toxicol. Applied Pharmacol, 235: 39-46.
CrossRef - Jones, P.A. and S.B. Baylin, 2002. The fundamental role of epigenetic events in cancer. Nat. Rev. Genet., 3: 415-428.
CrossRefPubMedDirect Link - Karavoltsos, S., A. Sakellari, N. Mihopoulos, M. Dassenakis and M.J. Scoullos, 2008. Evaluation of the quality of drinking-water in regions of Greece. Desalination, 224: 317-329.
Direct Link - Ke, Q., T.P. Ellen and M. Costa, 2008. Nickel compounds induce histone ubiquitination by inhibiting histone deubiquitinating enzyme activity. Toxicol. Applied Pharmacol., 228: 190-199.
CrossRef - Leader, J.E., C. Wang, M. Fu and R.G. Pestell, 2006. Epigenetic regulation of nuclear steroid receptors. Biochem. Pharmacol., 72: 1589-1596.
CrossRef - Leivadara, S.V., A.D. Nikolaou and T.D. Lekkas, 2008. Determination of organic compounds in bottled waters. Food Chem., 108: 277-286.
CrossRef - Li, R., Y.Y. Shugart, W. Zhou, Y. An and Y. Yang et al., 2009. Common genetic variations of the cytochrome P450 1A1 gene and risk of hepatocellular carcinoma in a Chinese population. Eur. J. Cancer, 45: 1239-1247.
Direct Link - Lukusa, T. and J.P. Fryns, 2008. Human chromosome fragility. Biochim. Biophys. Acta, 1779: 3-16.
PubMed - Majumder, S., H. Kutay, J. Datta, D. Summers, S.T. Jacob and K. Ghoshal, 2006. Epigenetic regulation of metallothionein-i gene expression: Differential regulation of methylated and unmethylated promoters by DNA methyltransferases and methyl CpG binding proteins. J. Cell. Biochem., 97: 1300-1316.
PubMed - Mohammadi, A., A. Vaziri Gohar and M.R. Shakibaie, 2008. Mutations in tumor suppressor TP53 gene in formalin- fixed, paraffin embedded tissues of Squamous Cell Carcinoma (SCC) of lung cancer. Am. J. Biochem. Biotehcnol., 4: 1-6.
CrossRefDirect Link - Melnikova, V.O. and H.N. Ananthaswamy, 2005. Cellular and molecular events leading to the development of skin cancer. Mutat. Res. Fundam. Mol. Mechanisms Mutagenesis, 571: 91-106.
CrossRefDirect Link - Monsef, N., L. Helczynski, A. Lundwall, S. Pahlman and A. Bjartell, 2007. Localization of immunoreactive HIF-1alpha and HIF-2alpha in neuroendocrine cells of both benign and malignant prostate glands. Prostate, 67: 1219-1229.
PubMed - Nebert, D.W., A.L. Roe, M.Z. Dieter, W.A. Solis, Y. Yang and T.P. Dalton, 2000. Role of the aromatic hydrocarbon receptor and [Ah] gene battery in the oxidative stress response, cell cycle control and apoptosis. Biochem. Pharmacol., 59: 65-85.
PubMed - Nielsen, G.D., U. Soderberg, P.J. Jorgensen, D.M. Templeton, S.N. Rasmussen, K.E. Andersen and P. Grandjean, 1999. Absorption and retention of nickel from drinking-water in relation to food intake and nickel sensitivity. Toxicol. Applied Pharmacol., 154: 67-75.
Direct Link - Patterson, T.J., M. Ngo, P.A. Aronov, T.V. Reznikova, P.G. Green and R.H. Rice, 2003. Biological activity of inorganic arsenic and antimony reflects oxidation state in cultured human keratinocytes. Chem. Res. Toxicol., 16: 1624-1631.
Direct Link - Ravanat, J.L., T. Douki and J. Cadet, 2001. Direct and indirect effects of UV radiation on DNA and its components. J. Photochem. Photobiol. Biol., 63: 88-102.
CrossRef - Rayman, M.P., 2000. The importance of selenium to human health. Lancet, 356: 233-241.
CrossRefPubMedDirect Link - Asadikaram, G., M. Mirzai, N. Naimi and R. Dargahi, 2008. Removal of arsenic using Acetobacter Xylinum cellulose. J. Biol. Sci., 8: 209-212.
CrossRef - Rossman, T.G., A.N. Uddin and F.J. Burns, 2002. Arsenite cocarcinogenesis: An animal model derived from genetic toxicology studies. Environ. Health. Perspect., 110: 749-752.
PubMedDirect Link - Rossman, T.G., A.N. Uddin, F.J. Burns and M.C. Bosland, 2001. Arsenite is a cocarcinogen with solar ultraviolet radiation for mouse skin: An animal model for arsenic carcinogenesis. Toxicol. Applied Pharmacol., 176: 64-71.
CrossRef - Sarafanov, A.G., T.I. Todorov, A. Kajdacsy-Balla, M.A. Gray, V. Macias and J.A. Centeno, 2008. Analysis of iron, zinc, selenium and cadmium in paraffin-embedded prostate tissue specimens using inductively coupled plasma mass-spectrometry. J. Trace Element Med. Biol., 22: 305-314.
CrossRef - Satoh, E., I. Yasuda, T. Yamada Y. Suzuki and T. Ohyashiki, 2007. Involvement of NO generation in aluminum-induced cell death. Biol. Pharm. Bull., 30: 1390-1394.
Direct Link - Schnekenburger, M., L. Peng and A. Puga, 2007. HDAC1 bound to the Cyp1a1 promoter blocks histone acetylation associated with Ah receptor-mediated trans-activation. Biochim. Biophys. Acta Gene Struct. Expression, 1769: 569-578.
CrossRef - Sell, S., 2004. Stem cell origin of cancer and differentiation therapy. Crit. Rev. Oncol./Hematol., 51: 1-28.
CrossRefDirect Link - Stellman, S.D., M.V. Djordjevic, J.A. Britton, J.E. Muscat and M.L. Citron et al., 2000. Breast cancer risk in relation to adipose concentrations of organochlorine pesticides and polychlorinated biphenyls in Long Island, New York. Cancer Epidemiol. Biomarks Prev., 9: 1241-1249.
Direct Link - Sun, X., J.M. Fontaine, I. Bartl, B. Behnam, M.J. Welsh and R. Benndorf, 2007. Induction of Hsp22 (HspB8) by estrogen and the metalloestrogen cadmium in estrogen receptor-positive breast cancer cells. Cell Stress Chaperones, 2: 307-319.
PubMed - Suzuki, K.T., A. Katagiri, Y. Sakuma, Y. Ogra and M. Ohmichi, 2004. Distributions and chemical forms of arsenic after intravenous administration of dimethylarsinic and monomethylarsonic acids to rats. Toxicol. Applied Pharmacol., 198: 336-344.
PubMed - Tamaru, H. and E.U. Selker, 2001. A histone H3 methyltransferase controls DNA methylation in Neurospora crassa. Nature, 414: 277-283.
CrossRef - Vahter, M., 2000. Genetic polymorphism in the biotransformation of inorganic and its role in toxicity. Toxicol. Lett., 112-113: 209-217.
CrossRef - Vogelstein, B. and K.W. Kinzler, 2004. Cancer genes and the pathways they control. Nat. Med., 10: 789-799.
PubMed - Wei, Y.D., K. Tepperman, M.Y. Huang, M.A. Sartor and A. Puga, 2004. Chromium inhibits transcription from polycyclic aromatic hydrocarbon-inducible promoters by blocking the release of histone deacetylase and preventing the binding of p300 to chromatin. J. Biol. Chem., 279: 4110-4119.
Direct Link - Wu, J.P., L.W. Chang, H.T. Yao, H. Chang and H.T. Tsai et al., 2008. Involvement of oxidative stress and activation of aryl hydrocarbon receptor in elevation of CYP1A1 expression and activity in lung cells and tissues by arsenic: An in vitro and in vivo study. Toxicol. Sci., 107: 385-393.
CrossRefDirect Link - Xiang, N., R. Zhao, G. Song and W. Zhong, 2008. Selenite reactivates silenced genes by modifying DNA methylation and histones in prostate cancer cells. Carcinogenesis, 29: 2175-2181.
CrossRef - Zhang, Q., T. Kluz, K. Salnikow and M. Costa, 2002. Comparison of the cytotoxicity, cellular uptake and DNA-protein crosslinks induced by potassium chromate in lymphoblast cell lines derived from three individuals. Biol. Trace Element Res., 86: 11-22.
PubMed - Zhang, R.Y., Y. Lui, D.W. Pang, R.X. Cai and Y.P. Qi, 2002. Spectroscopic and voltammetric study on the binding of aluminium (III) to DNA. Anal. Sci., 18: 761-766.
Direct Link - Zou, Y., P. Niu, J. Yang, J. Yuan, T. Wu and X. Chen, 2008. The JNK signaling pathway is involved in sodium-selenite-induced apoptosis mediated by reactive oxygen in HepG2 cells. Cancer Biol. Ther., 7: 689-696.
PubMed - Kinzler, K.W. and B. Vogelstein, 1997. Cancer-susceptibility genes: Gatekeepers and caretakers. Nature, 386: 761-763.
Direct Link - Yu, M.W., I.S. Horng, K.H. Hsu, Y.C. Chiang, Y.F. Liaw and C.J. Chen, 1999. Plasma selenium levels and the risk of hepatocellular carcinoma among men with chronic hepatitis virus infection. Am. J. Epidemiol., 150: 367-374.
Direct Link