
M. Soltanialvar
Ph.D Student, Science and Research Branch, Isamic Azad University (IAU), Tehran, Iran
H. Shoushtari
Division of Avian Diseases and Research, Razi Vaccine and Serum Research Institute, Karaj, Tehran, Iran
M. Bozorgmehrifard
Division of Avian Diseases, Tehran University, Iran
S. Charkhkar
Division of Avian Diseases, Science and Research Branch, Isamic Azad University, Tehran, Iran
F. Eshratabadi
Division of Avian Diseases and Research, Razi Vaccine and Serum Research Institute, Karaj, Tehran, Iran
Journal of Biological Sciences
Year: 2010 | Volume: 10 | Issue: 2 | Page No.: 145-150







ABSTRACT
A total of 512 tissue samples collected from 30 farms located in various states of Iran during 2008-2009 as part of a program to monitor AIV infection in Iran poultry population. Avian influenza virus was isolated from poultry farms with history of respiratory illness and increased mortality. Nucleotide sequence analysis of five representative isolates confirmed that all isolates possessed one type of amino acid motif (R-S-S-R/GL) at cleavage site of HA, Deduced amino acid sequences showed the presence of L226 (234 in H9 numbering) in all five Iranian isolates which indicates a preference to binding of α (2-6) sialic acid receptors. The viruses' surface glycoproteins genes were >90% similar to those of A/Quail/Hong Kong/G1/97 (H9N2) lineage. The neuraminidase stalks regions in these viruses had no deletion as compared to that A/Dk/HK/ Y280/97 lineage (Ck/Bei-like viruses) and the 2 human isolates A/HK/1073/99, A/HK/1074/99. The hemadsorbing site of neuraminidase had up to 3 amino acid substitutions and is different from those of earlier Iranian viruses. Phylogenetic analysis HA and NA genes showed that they share a common ancestor Qa/HK/G1/97 isolate which had contributed internal genes of H5N1 virus.
PDF Abstract XML References Citation
How to cite this article
M. Soltanialvar, H. Shoushtari, M. Bozorgmehrifard, S. Charkhkar and F. Eshratabadi, 2010. Molecular Characterization of Hemagglutinin and Neuraminidase Genes of H9N2 Avian Influenza Viruses Isolated From Commercial Broiler Chicken in Iran. Journal of Biological Sciences, 10: 145-150.
DOI: 10.3923/jbs.2010.145.150
URL: https://scialert.net/abstract/?doi=jbs.2010.145.150
DOI: 10.3923/jbs.2010.145.150
URL: https://scialert.net/abstract/?doi=jbs.2010.145.150
INTRODUCTION
Influenza viruses belong to the orthomyxoviridae family of RNA viruses and are divided into five genera: Influenza A, B and C, Thogtovirus and Isavirus. Avain Influenza (AI) is a highly contagious disease caused by type A influenza viruses which have negative-sense, single-stranded genomes that encode at least ten proteins including two surface glycoproteins [haemagglutinin (HA) and neuraminidase (NA)], nucleoprotein (NP), three polymerase proteins [polymerase basic (PB1), (PB2) and polymerase acidic (PA)], two matrix (M1 and M2) proteins and two non-structural (NS1 and NS2) proteins. Genetically and antigenically, AIV exist as multiple subtypes based on the two glycoproteins (HA and NA) on the virion surface. To date, 16 HA (HA1-16) and 9 NA (NA1-9) subtypes have been identified in aquatic birds (Fouchier et al., 2005). The influenza A virion is studded with glycoprotein spikes of HA and NA, in a ratio of approximately four to one. During replication, influenza A viruses require the activities of two viral surface glycoproteins, hemagglutinin (HA), responsible for binding to terminal sialic acid on cell surface glycoconjugates and neuraminidase (NA), with an associated enzymatic activity that removes sialic acid from host cell glycoconjugates as well as newly synthesized viral proteins to facilitate the budding of progeny virions from cells (Kobasa et al., 2001).
In the last decade, H9N2 viruses had caused disease outbreaks in chicken in many parts of the world including Germany, Italy, Ireland, Pakistan, Saudi Arabia, South Korea, UAE, Indian, Israel, Jordan, China, South Africa and USA (Alexander, 2007; Nagarajan et al., 2009). Avian H9N2 viruses were also recovered from pigs in Hong Kong, China and Indonesia (Ninomiya et al., 2002). Some avian H9 viruses have acquired receptor binding characteristics typical of human strains, increasing the potential for reassortment in both human and pig respiratory tracts (Suzuki, 2005). In 1999 and 2003, human cases of H9N2 virus were recorded in Hong Kong although, no death was reported (Peiris et al., 1999; Riedel, 2006). Crossing the species barrier to mammals highlights the pandemic potential of H9N2 virus. With the emergence of non-human H9N2 isolates with avian characteristics, it is important to study the H9N2 isolates from avian hosts in addition to those obtained from humans for pandemic influenza understanding and preparedness.
H9N2 viruses circulate widely in the Middle East and are associated with serious disease in poultry. In this study, we characterized HA and NA genes and proteins of 5 Iranian isolate which have been isolated from Commercial broiler chicken in the Iran between 2008 and 2009. These isolates resulted in rapid mortality due to tracheitis and respiratory congestion. We delineated the genomes, receptor binding profile and HB sites of NA gene of these field isolates and we also established their phylogenetic relationship to the other Asian H9N2 lineages.
MATERIALS AND METHODS
Sampling and virus isolation: Samples collected from 30 farms were received between April 2008 and February 2009, from various parts of the country. The tissue samples usually included trachea and lung. The samples were collected from flocks with the history of acute respiratory illness.
Virus isolation was carried out in 9-11 days old SPF embryonated chicken eggs following the standard protocol (OIE, 2005). Briefly, triturated tissue samples treated with 2 antibiotic and antimycotic solution (pecillin 10000 unit mL-1, streptomycin 10000 unit mL-1 and nystatin 20000 unit mL-1) for 30 min at 37°C and clarified by centrifugation at 1500 RPM for 10 min were inoculated through allantoic routes. The allantoic fluid was harvested after incubation at 37°C for 72-96 h and clarified by centrifugation at 1500 g for 15 min at 4°C. The HA test negative samples were given two more passages and tested again before being declared negative for AIV isolation.
The identification of viruses subtype was determined by a standard Hemagglutination Inhibition (HI) and Neuraminidase Inhibition (NI) tests (Shortridge et al., 1997). The 5 virus isolates obtained in this study were named as follows: (A/chicken/Iran/RZ28/2008,A/chicken/Iran/RZ36/2008,A/chicken/Iran/RZ37/2008, A/chicken/Iran/RZ42/2008 and A/chicken/Iran/RZ53/2008).
RT-PCR and sequence analysis: The viral RNA was extracted directly from the allantoic fluid by means of the high pure viral Nucleic Acid Kit (Roche Germany). Purified genomic RNA was used to generate cDNA clones by (RT-PCR) according to the standard procedure. The RT-PCR was performed using specific primers for 940 (nucleotides 1 to 940) and 560 (nucleotides 920 to 1470) bp fragment of NA and 1740 bp for HA.
Primers used for HA amplication were:
| • | Forward primer (1720 bp): 5'- AGC AAA AGC AGG GG-3' |
| • | Reverse primer (1720 bp): 5'- AGT AGA AAC AAG GGT GTT-3' |
Primers used for NA amplication were:
| • | Forward primer (980 bp): 5'- GCAAAAGCAGGA GTGAAATGA-3' |
| • | Reverse primer (980 bp): 5'- AGTCCTGAGCAC AAATAATGC-3' |
| • | Forward primer (560 pb): 5'-TTAGATGTGTTTGCA GGGCAC-3' |
| • | Reverse primer (560 bp): 5'- GGTTCTAAAATTGCG AAAGCC-3' |
The PCR products were purified using high pure product purification kit (Roche Germany). The PCR products were applied to Low Melting Point (LMP) agarose and the distinct bands were purified from gel for sequencing (MWG Co., Germany).
Nucleotide and deduced amino acid sequences of the HA and NA genes were edited with the Editseq (DNASTER Software package Version 5.2 (Nucleotide and deduced amino acid sequences were aligned by ClustalW, Version 1.4.
Nucleotide sequences of the HA and NA genes were used for phylogenetic tree construction. The phylogenetic analysis was performed with the MegAlign program 2.8.
Nucleotide sequence accession No.: The sequences determined in this study are available in the GenBank under accession No.: GUO71964-GUO7173.
RESULTS AND DISCUSSION
Disease signs in the field: The symptoms included anorexia, reduced water consumption, gasping and conjunctivitis with facial edema. The major post mortem findings were tracheitis, respiratory congestion and hemorrhages on proventriculus. The mortality rose sharply (1 to 10%), resulting in considerable economic losses.
HA gene of the Iran viruses: The sequenced part of the HA gene contains 1720 nucleotides coding 566 amino acid residues. This protein region includes a complete receptor-binding pocket and cleavage site. The Iranian isolates did not exhibit insertions or deletions within this region, but rather numerous point mutations were registered. These viruses differed from other viruses by 10 amino acid substitutions at the following positions: 7 (V to I) 40 (A to T) 273 (M to L) 278 (R to k) 283 (N to S) 285 (D to N) 501) R to k) 535 (H to L) 555) K to R) 556 (S to C).
Nucleotide sequence analysis of the HA gene showed that these isolates are very closely related (96.7-99.6%) and shared a homology of (92/0-96%) with H9N2 isolates of A/ Qa/HK/G1/97 lineage and the 2 human isolates A/HK/1073/99 and A/ HK/1074/99. The deduced amino acid sequences revealed that the HA cleavage site motif sequence was (RSSR/GLF) which is similar to the other Asian H9N2 viruses including those from, Pakistan, Saudi Arabia, Israel and Hong Kong (Table 1). All isolates carry amino acid substitution Q226L (H3 numbering throughout this text) in the receptor-binding site, a mutation that correlates with a shift in affinity of the HA from the avian type sialic receptors to the human type and from a preference for α2'-3' link to a preference for α2'-6' link between the sialic acid residues and galactose (Suzuki, 2005). Analysis of HA protein sequences showed that these H9N2 viruses have seven glycosylation sites with the N-X-T/S motif in which X may be any amino acid except proline. Five of these sites were located in the HA1 region (positions 21, 97,133,290,279) and two in HA 2 region (positions 484, 543).
The phylogenetic relationships between the HA genes of this Iranian isolates and those of selected H9N2 viruses isolated in several other countries showed that all the Iranian isolates belonged to a single lineage which is closer to viruses from the so-called G1 lineage (Fig. 1).
The Iranian isolates (these isolates and earlier isolates) can be divided into three subgroups. The first subgroup includes viruses, isolated in 1998 to 2002 together with the viruses A/HK/1073/99 and A/HK/1074/99, isolated from human and the viruses of Pakistan, Saudi Arabia and Hong Kong which isolated from 1998 until the end of 2002. The Iranian strains isolated during 2003 to 2005 fall into the second subgroup .The strains isolated after 2005 and until the end of the study (February 2009) fall into the third subgroup. All of this new isolates fall into the third subgroup (Fig. 1).
NA gene of the Iranian viruses: In this study, 1447 base pair of the NA genes were sequenced and amino acid sequences 469 of the NA genes 5 isolates were deduced from the nucleotide sequence. The amino acid sequences of the isolates showed (90/6-99/6) identity to each other. Amino acids of HB_site at position 366-373 are IRKDSRAG except A/chicken/Iran/RZ53/2008 isolate which is IRKDSREG, amino acids of at position 399-404 have two type DSDNWS or DSDNLS (Table 2).
The neuraminidase stalk regions in these viruses had no deletion as compared to that A/Dk/HK/ Y280/97 lineage(ck/Bei-like viruses) and the 2 human isolates A/HK/1073/99 , A/HK/1074/99. Phylogenetic analysis of the N2 gene showed that all the NA genes of the Iranian H9N2 viruses fall into a single group within a G1-like lineage (Fig. 2). These findings further confirm the Asian origins of the Iranian viruses. In turn, this group subdivides into two main subgroups: the first of these includes viruses isolated from 1998 to 2004 and surprisingly the isolate A/chicken/Iran/RZ53/2008, the second subgroup includes strains isolated after 2004 (Fig. 2).
| Table 1: | Receptor-binding sites and cleavage site of H9N2 viruses |
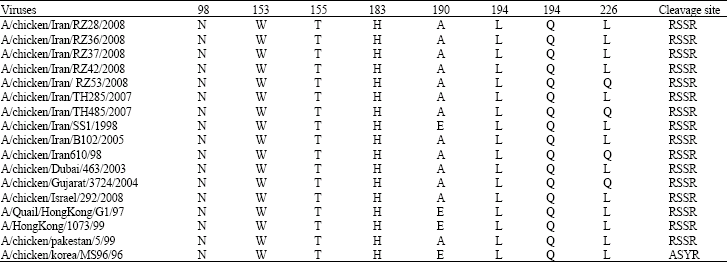 | |
| Table 2: | Amino acid substitutions in the hemadsorbing sites and deletion in stalk |
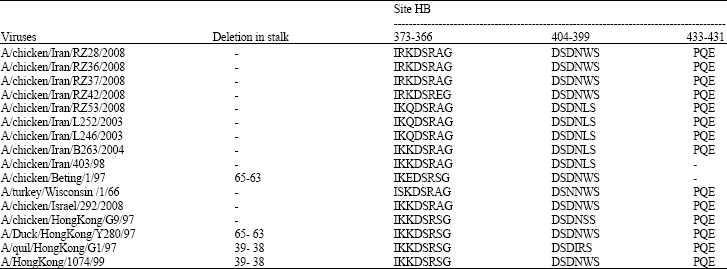 | |
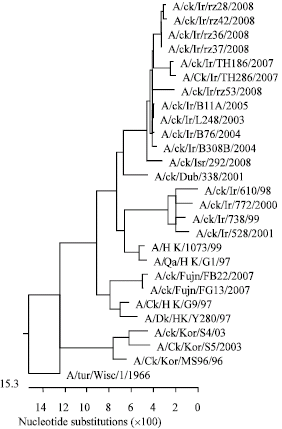 | |
| Fig. 1: | Phylogenetic relationships of hemagglutinin genes from H9N2 viruses isolated in Iran during 2008-2009 and other representative AIVs of this subtype. Ck: Chicken, Dk: Duck, Qa: Quail, Tur: Turkey, HK: Hong Kong, Kor: Korea, Wis: Wisconsin, Ir: Iran, Dub: Dubai, fujn: Fujian |
The H9N2 AIVs have been circulating in the domestic poultry in Iran for 12 years and were still isolated from chickens, even from some vaccinated flocks. Although, the H9N2 viruses are characterized as low pathogenic avian influenza (LPAI) viruses, they may cause high morbidity and mortality (Aamir et al., 2007; Kim et al., 2006; Nili and Asasi, 2002).
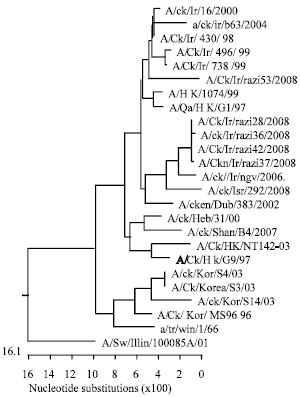 | |
| Fig. 2: | Phylogenetic relationships of neuraminidase genes from H9N2 viruses isolated in Iran during 2008-2009 and other representative AIVs of this subtype Ck: Chicken, Dk: Duck, Qa: Quail, Tur: Turkey, HK: Hong Kong, Kor: Korea, Wis: Wisconsin, Ir: Iran, Dub: Dubai, fujn: Fujian |
Guo et al. (2000) have reported that Ck/Beijing/1/94 is lethal for chickens in the field and also under experimental condition. In this study, overt clinical signs in chickens observed at affected farms included acute respiratory signs and 1-10% mortality rates during the outbreak. The clinical signs and mortality observed under field conditions could be due to the concurrent infections or environmental factors. Bano et al. (2003) and Nili and Asasi (2002) reported that H9N2 viruses with an IVP index of 0.00/3.00 could cause high mortality (20-65%) in the field due to environmental stress and secondary bacterial (E. coli) and Mycoplasma gallisepticum infections. Kishida et al. (2004) has reported that co-infection of Staphylococcus aureus or Haemophilus paragallinarum exacerbates H9N2 influenza A virus infection in chicken. The deduced amino acid sequences revealed that the HA cleavage site motif sequence was(RSSR/GLF ) which is similar to the other Asian H9N2 viruses including those from, Indian, Pakistan, Saudi Arabia, Israel and Hong Kong (Table 1). The absence of multiple basic amino acids confirms the low pathogenic nature of the Iranian H9N2 isolates. Nucleotide sequences of the HA cleavage sites of the Iranian H9N2 viruses showed that only one nucleotide substitution is required at the _2 position to change serine (S) to arginine (R) to produce the basic motif required for highly pathogenic viruses(R/K-X-R-R/GLF) (Perdue et al., 1997). Hence, continuous monitoring of evolution of HA gene cleavage motif of Iranian H9N2 viruses is essential. According to the phylogenetic analysis, all the Iranian isolates fall into a special group, related to the G1 lineage, a finding that may indicate a common origin of all the Iranian isolates from a comprises single progenitor. No Ck/Bei-like viruses and Korean- like viruses was found in chicken, population since the outbreak of H9N2 influenza in Iran in 1997. This evidence indicates that (HA genes) and (NA gene) of H9N2 influenza virus circulating in Iran during the past years were well conserved and the earliest Iranian isolates or neighboring countries may be considered to represent such a progenitor. Nevertheless, according to the phylogenetic analysis, the Iranian group was not homogeneous, so that the viruses may be distributed among two (NA gene) or three (HA gene) subgroups, associated with the time of their isolation. It seems that these differences are the result of accumulation of mutations among the viruses circulating within the poultry reservoir. One of the factors driving the evolution of low-pathogenic influenza viruses is the immunological pressure, which increases in the case of the ongoing vaccination. The postvaccinal specific immunity of the population could be decisive as a selective factor in ensuring prevalence of a viral strain which differed from the vaccinal strain and earliest Iranian isolates; however, there are great numbers of report that indicated the group viruses in Eurasian sublineage confer an immunity to each other (Aamir et al., 2007; Xu et al., 2007).
The deduced amino acid sequence at the receptor binding sites showed the presence of L226 (234 in H9 numbering) which is similar to A/Qa/ HK/G1/97 (Guan et al., 2000) and the 2 human isolates A/HK/1073/99, A/HK/1074/99. Amino acid differences in the receptor binding sites of HAs have been shown to be associated with differences in receptor binding specificity, (Suzuki, 2005) so these Iranian H9N2 isolates can bind to α(2, 6) glycosidic bond and have the potential to infect human beings. The hemadsorbing (HB) site of NA is highly conserved in equine and aquatic bird influenza viruses. However, avian viruses co-circulating in mammals such as pigs and humans accumulate various substitutions in the HB site, thus decreasing their hemadsorbing capacity (Baum and Paulson, 1991). Earlier studies have shown the HB site in the NA of Asian H9N2 viruses to be under positive selection pressure for mutations, which results in compatible combinations of HA and NA (Matrosovich et al., 2001). The mutations in the HB site were previously seen in viruses that bind to α-2,6-linked receptors. The NA of these new isolates carried substitutions (in spite of earlier Iranian isolates) in the HB site similar to those of other avian H9N2 viruses from Asia and human H9N2 viruses (Table 2). Together, human virus-like receptor specificity and similar substitutions in the HB region of the NA in these recent isolates suggest the possible role of poultry species such as chickens in the zoonotic transmission of influenza viruses from aquatic birds to humans (Gambarian et al., 2002).
REFERENCES
- Aamir, U.B., U. Wernery, N. Ilyushina and R.G. Webster, 2007. Characterization of avian H9N2 influenza viruses from United Arab Emirates 2000 to 2003. Virology, 361: 45-55.
CrossRefPubMedDirect Link - Alexander, D.J., 2007. An overview of the epidemiology of avian influenza. Vaccine, 25: 5637-5644.
CrossRef - Bano, S., K. Naeem and S.A. Malik, 2003. Evaluation of pathogenic potential of avian influenza virus serotype H9N2 in chickens. Avian Dis., 47: 817-822.
PubMedDirect Link - Baum, L.G. and J.C. Paulson, 1991. The N2 neuraminidase of human influenza virus has acquired a substrate specificity complementary to the hemagglutinin receptor specificity. Virology, 180: 10-15.
CrossRef - Fouchier, R.A.M., V. Munster, A. Wallensten, T.M. Bestebroer and S. Herfst et al., 2005. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J. Virol., 79: 2814-2822.
CrossRefDirect Link - Gambarian, A.S., S.S. Iamnikova, D.K. L'vov, J.S. Robertson, R.G. Webster and M.N. Matrosovich, 2002. Differences in receptor specificity between the influenza A viruses isolated from the duck, chicken and human. Mol. Biol. (Mosk.), 36: 542-549.
PubMedDirect Link - Guan, Y., K.F. Shortridge, S. Krauss, P.S. Chin and K.C. Dyrting et al., 2000. H9N2 influenza viruses possessing H5N1-like internal genomes continue to circulate in poultry in Southeastern China. J. Virol., 74: 9372-9380.
CrossRefDirect Link - Guo, Y.J., S. Krauss, D.A. Senne, I.P. Mo and K.S. Lo et al., 2000. Characterization of the pathogenicity of members of the newly established H9N2 influenza virus lineages in Asia. Virology, 267: 279-288.
CrossRefDirect Link - Kim, J.A., S.H. Cho, H.S. Kim and S.H. Seo, 2006. H9N2 influenza viruses isolated from poultry in Korean live bird markets continuously evolve and cause the severe clinical signs in layers. Vet. Microbiol., 118: 169-176.
CrossRefDirect Link - Kishida, N., Y. Sakoda, M. Eto and H. Kida, 2004. Co-infection of Staphylococcus aureus or Haemophilus paragallinarum exacerbates H9N2 influenza A virus infection in chicken. Arch. Virol., 149: 2095-2104.
CrossRefDirect Link - Kobasa, D., K. Wells and Y. Kawaoka, 2001. Amino acides responsible for the absolute sialidase activity of the influenza A virus neuraminidase: Relationship to growth in intestine. J. Virol., 75: 11773-11780.
CrossRefDirect Link - Nagarajan, S., K. Rajukumar, C. Tosh, V. Ramaswamy and K. Purohit et al., 2009. Isolation and pathotyping of H9N2 avian influenza viruses in Indian poultry. Vet. Microbiol., 133: 154-163.
CrossRef - Nili, H. and K. Asasi, 2002. Natural cases and an experimental study of H9N2 avian influenza in commercial broiler chickens of Iran. Avian Pathol., 31: 247-252.
PubMedDirect Link - Ninomiya, A., A. Takda, K. Okazaki, K.F. Shortridge and H. Kida, 2002. Seroepidemiological evidence of avian H4, H5 and H9 influenza viruses transmission to pigs to South Eastern China. Vet. Microbiol., 88: 107-114.
CrossRef - Peiris, M., K.Y. Yuen, C.W. Leung, K.H. Chan and P.L.S. Ip et al., 1999. Human infection with influenza H9N2. Lancet, 354: 916-917.
CrossRefDirect Link - Perdue, M.L., M. Garcia, D. Senne and M. Fraire, 1997. Virulence associated sequence duplication at the hemagglutinin cleavage site of avian influenza viruses. Virus Res., 49: 173-186.
PubMedDirect Link - Riedel, S., 2006. Crossing the species barrier: The threat of an avian influenza pandemic. Proc. Bayl. Univ. Med. Cent., 19: 16-20.
Direct Link - Shortridge, K.F., W.K. Butterfield, R.G. Webster and C.H. Campbell, 1977. Isolation and characterization of influenza A viruses from avian species in Hong Kong. Bull. WHO., 55: 15-20.
Direct Link - Suzuki, Y., 2005. Sialobiology of influenza: Molecular mechanism of host range variation of influenza viruses. Biol. Pharm. Bull., 28: 399-408.
CrossRefDirect Link - Xu, K.M., G.J.D. Smith, J. Bahl, L. Duan and H. Tai et al., 2007. The genesis and evolution of H9N2 influenza viruses in poultry from Southern China, 2000 to 2005. J. Virol., 81: 10389-10401.
CrossRefPubMedDirect Link