
S.K. Sharma
Regional Medical Research Centre, North East Region, Indian Council of Medical Research, P.O. Box 105, Dibrugarh-786 001, Assam, India
J. Mahanta
Regional Medical Research Centre, North East Region, Indian Council of Medical Research, P.O. Box 105, Dibrugarh-786 001, Assam, India
Journal of Biological Sciences
Year: 2009 | Volume: 9 | Issue: 3 | Page No.: 288-291







ABSTRACT
The present study is an attempt to evaluate the relationship of haemoglobinopathies, particularly Hb E and Plasmodium falciparum malaria in northeastern region of India. The diverse autochthonous inhabitant of this part of India exhibits variable gene frequency for βE-globin gene. The geo-climatic condition of the region supports transmission of Plasmodium falciparum malaria in northeastern parts of India. The study revealed that HbE is predominant with a variable gene frequencies in ethnic groups affiliated to Tibeto-Burman linguistic families. Prevalence of Hb E is also associated with the linguistic affiliation of various Tibeto-Burman linguistic families inhabiting in malaria endemic northeast India. We have also observed a positive correlation (R2 = 0.703) of βE-globin gene frequency and mean incidence of Plasmodium falciparum infection (Pf %) in malaria endemic zones.
PDF Abstract XML References Citation
How to cite this article
S.K. Sharma and J. Mahanta, 2009. Prevalence of Haemoglobin Variants in Malaria Endemic Northeast India. Journal of Biological Sciences, 9: 288-291.
DOI: 10.3923/jbs.2009.288.291
URL: https://scialert.net/abstract/?doi=jbs.2009.288.291
DOI: 10.3923/jbs.2009.288.291
URL: https://scialert.net/abstract/?doi=jbs.2009.288.291
INTRODUCTION
Haemoglobinopathies are the worldwide prevalent monogenic genetic disorders with variable geographic distribution (Krafft and Breymann, 2004). In Southeast Asia and the Indian subcontinent, this has been considered as common disorders of blood posing a major genetic and public health problem (Fucharoen and Winichagoon, 1997). Amongst variant haemoglobins, haemoglobin E (Hb E, β26 GAG→AAG) is the most common β-thalassaemic haemobinopathy in Asian population affecting about 30 million inhabitants of Southeast Asian (Lukens, 1999). High gene frequency for Hb E is prevalent in autochthonous inhabitant, having linguistic and cultural affiliation with the population of Southeast Asian countries, of the northeastern part of Indian (Deka et al., 1987). Sickle cell haemoglobin (Hb S, β6 GAG→GTG) in this part of India is restricted to the Tea garden labour communities, a group of population brought to Assam by the British colonial tea planters as indentured labourer from central, eastern and southern India during mid 19th century (Balgir and Sharma, 1988). Besides, Haemoglobin D (HbD, β121 GAA→CAA) is reported in an Ahom family, affiliated Tai-Kadai linguistic group (Sharma et al., 2003).
The unique geo-climatic conditions of the northeastern part of India facilitate transmission of malaria in this part of the country (Mohapatra and others, 1998). Malaria in this region is predominantly contributed by Plasmodium falciparum (P. falciparum) with widespread distribution of Chloroquine resistant strains (Mohapatra et al., 1998; Satyanarayna et al., 1991). Hence, overlapping of haemoglobinopathies and P. falciparum malaria persists in northeast India.
MATERIALS AND METHODS
In the present study, seven homogenously distributed ethnic groups concentrated in different geographical locations of northeast India with variable Plasmodium falciparum infection (Pf %) were included (Fig. 1). Linguistic affiliations of the ethnic groups, excluding Tea garden labour community, are represented in Fig. 2. The Tea garden labour community is predominantly localized in Assam. Incidence of Plasmodium falciparum malaria (pf%) of three consecutive years, of the respective study districts, were acquired from the district health authorities of the respective states of Northeast India (Table 1).
Six hundred fifty two blood samples were obtained randomly from the volunteers, in EDTA vials, affiliated to seven ethnic groups of the region after obtaining written consent from each individual (Table 1). The seven ethnic groups represents in the study are Tangsa and Yobin (Lisu) from the state of Arunachal Pradesh, Mishing, Deori and Tea garden Labour community of Assam, Mizo’s from Mizoram and Tripuri from Tripura State (Table 1).
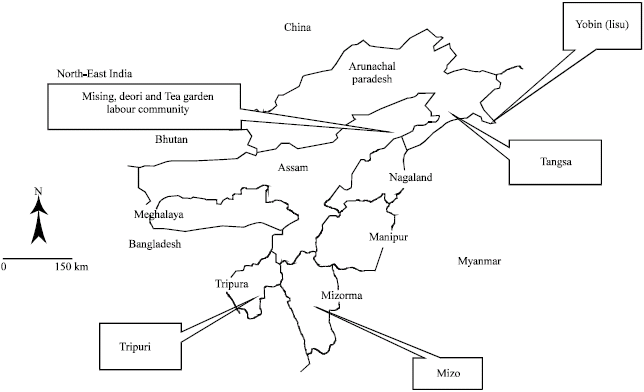 | |
| Fig. 1: | Distribution of the study population in Northeast India |
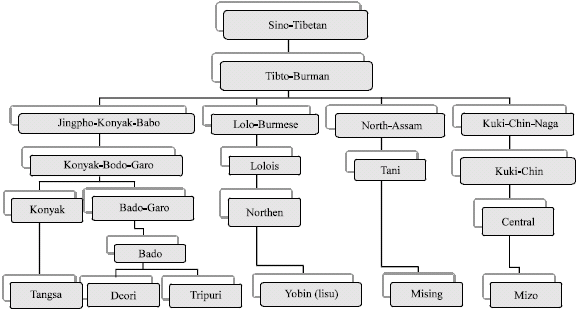 | |
| Fig. 2: | Linguistic affiliation of the study population |
| Table 1: | Detail about the study population and malaria situation of the study area |
 | |
Haemolysate of individual samples were prepared and presence of haemoglobin variants was identified initially by electrophoresis of haemolysate on cellulose acetate using Tris-EDTA-Borate buffer pH 8.6 (Lewis et al., 2001). Sickleing in whole blood, in presence of buffered Sodium dithionite, was performed simultaneously for all samples to confirm the presence of Sickle cell haemoglobin (Lewis et al., 2001). Further, an aliquot of 5 μL of blood was transferred to 1 mL of Hemolysis Reagent, a component of BioRad -thalassaemia Short Program kit and stored at 4°C. The samples were transported to the Central laboratory maintaining the cold chain and analysed in the VariantTM Hemoglobin Testing System (BioRad) using β-thalassaemia Short Program within ten days of sample collection adopting standard protocol.
RESULTS AND DISCUSSION
Hb E is the most prevalent variant haemoglobin in ethnic groups affiliated to Tibeto-Burman linguistic family. Gene frequency for βE-globin gene in these groups ranged from 0.006-0.569 with an overall prevalence of 0.266. In addition to malaria status of the area, βE-globin gene frequency is also associated with the population groups affiliated to Tibeto-Burman linguistic family tree.
The Deori and Tripuri tribes affiliated to Bodo subgroup, expressed high gene frequency (0.569 and 0.525) for βE-globin gene. Substantially high gene frequency (0.403) was also observed in Mishing tribe, affiliated to Tani subgroup. The Tangsas, affiliated to Konyak subgroup of Tibeto-Burman linguistic family, indicates considerably low (0.084) gene frequency for βE-globin gene. Significantly lower βE-globin gene frequency was depicted in Yobin (0.006) and Mizo (0.008), affiliated to Lolo-Burmese and Kuki-chin-Naga subgroups of Tibeto-Burman linguistic family tree respectively. The gene frequency for sickle cell haemoglobin in Tea garden labour community is 0.090.
Further, the study also indicated a positive correlation (R2 = 0.703) of βE-globin gene frequency and mean incidence of Plasmodium falciparum infection (Pf%) (Fig. 3). βE-globin gene frequency is considerably high in Deori (0.569), Tripuri (0.525) and Mishing (0.403) tribes in areas with high pf%. In contrast, substantially low gene frequency for βE-globin gene is observed in Mizo (0.008) population inhabited in a highly malaria endemic belt (Health and Family Welfare Department, Government of Mizoram, http://healthmizoram.nic.in/malaria_programme.htm). Similarly, in a meso to hyper endemic malarial zone, Tangsa also demonstrated low gene frequency (0.084) for βE-globin gene. However, in a relatively malaria free area of Vijaynagar circle of Changlang district, the Yobins exhibits lowest gene frequency (0.006) for βE- globin gene.
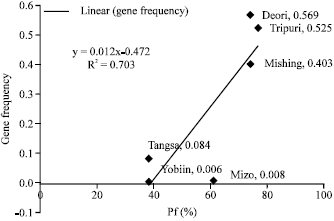 | |
| Fig. 3: | Prevalence of βE-globin gene in the different ethnic groups of northeastern region and P. falciparum status of the study area |
Variable gene frequency for βE-globin gene in autochthonous inhabitants of malaria endemic northeast India though partially support the malaria hypothesis, still earlier postulation on association of number of other evolutionary factors at microgeographic levels is also indicated in the present study (Livingstone, 1983, 1985). Occurrence of significantly low gene frequency (0.008) for βE-globin gene in Mizo population of a highly malaria endemic district of northeast India is probably associated with genetic diversity of the population. Linguistically, Mizo’s are affiliated to Kuki-Chin-Naga sub group of Tibeto-Burman linguistic family (Gordon, 2005). Likewise, in hyperendemic malarial background lower mean Hb E frequency is prevalent in Thai populations, originating from Upper Myanmar and southwestern Yunnan, is reported (Poolsuwan, 2003). The prevalence of lower gene frequency in Tangsa population of a meso to hyper endemic malarial zone may have similar explanation. Neighbour-joining tree constructed on DA distance matrix depicting the genetic relationship between the twenty three populations of Arunachal Pradesh, based on two classical genetic markers revealed, genetic difference of Tangsa people from other tribes (Krithika et al., 2007). Occurrence of low gene frequency for βE-globin gene in Yobins (Lisu) having ancestral origin from Myanmar and Yunnan also indicates linguistic or affiliation of the population in prevalence of βE-globin gene. Hence, in addition to a positive correlation of the βE-globin gene with Pf% as malaria endemicity (Fig. 3), genetic affiliation of the population, it reflects that other genetic polymorphism including thalassemia and G-6PD deficiency may also probably associated with the distribution of HbE in malaria endemic zones of northeast India.
ACKNOWLEDGMENTS
Researchers are thankful to Dr. K. Narain, Dr. H.K.D as and Dr. H.K. Chaturvadi for their kind help and support during the fieldwork at Vijoynagar circle of Arunachal Pradesh. Thanks are also due to Mr. R. Dutta and Mr. S. Gogoi for technical assistance.
REFERENCES
- Fucharoen, S. and P. Winichagoon, 1997. Hemoglobinopathies in Southeast Asia: Molecular biology and clinical medicine. Hemoglobin, 21: 299-319.
PubMed - Gordon, R.G. Jr., 2005. Ethnologue: Languages of the World 15th Edn., Tex.: SIL International, Dallas,.
Direct Link - Krafft, A. and C. Breymann, 2004. Haemoglobinopathy in pregnancy: Diagnosis and treatment. Curr. Med. Chem., 11: 2903-2909.
Direct Link - Krithika, S., S. Maji and T.S. Vasulu, 2007. Geographic contiguity, patterns of gene flow and genetic affinity among the tribes of Arunachal Pradesh. India Int. J. Hum. Genet., 7: 267-276.
Direct Link - Poolsuwan, S., 2003. Testing the malaria hypothesis for the case of Thailand: A genetic appraisal. Hum. Biol., 75: 585-605.
PubMed - Satyanarayna, S., S.K. Sharma, P.K. Chelleng, P. Dutta, L.P. Dutta and R.N.S. Yadav, 1991. Chloroquine resistant P. falciparum malaria in Arunachal Pradesh. Indian J. Malariol., 28: 137-140.
PubMed - Lewis, S.M., B.J. Bain and I. Bates, 2001. Dacie and Lewis Practical Haematology. 9th Edn., Churchill Livingstone, London, ISBN-13: 9780443063770, Pages: 633.
Direct Link