
Nurhaida Kamaruddin
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Farah-Diba A. Bakar
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Nor M. Mahadi
Malaysia Genome Institute, Heliks Emas Block, UKM-MTDC Smart Technology Center, 43600 UKM, Bangi, Selangor, Malaysia
Abdul M.A. Murad
Malaysia Genome Institute, Heliks Emas Block, UKM-MTDC Smart Technology Center, 43600 UKM, Bangi, Selangor, Malaysia
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 4 | Page No.: 730-737







ABSTRACT
The isolation and characterization of a gene encoding the regulatory subunit of cAMP-dependent protein kinase (CgPKAR) of the fungal plant pathogen Colletotrichum gloeosporioides is described in this research. The CgPKAR gene and its corresponding cDNA have been cloned and sequenced. Sequence analysis revealed that CgPKAR contains one intron located at position -475 to -539 from the start codon (ATG). The gene encodes a protein comprising of 391 amino acids and showing 90% identity to Colletotrichum lagenarium protein kinase A regulatory subunit. The deduced CgPKAR contains predicted features of type II regulatory subunits which include a serine in the phosphorylation site, a kinase inhibitor domain and two cAMP-binding domains. Southern blot analyses indicated that CgPKAR is a single-copy gene in C. gloeosporioides. Real time RT-PCR analyses indicated that CgPKAR is highly expressed in conidia and appressoria as compared to germinating conidia and mycelia.
PDF Abstract XML References Citation
How to cite this article
Nurhaida Kamaruddin, Farah-Diba A. Bakar, Nor M. Mahadi and Abdul M.A. Murad, 2008. Isolation and Characterisation of a Gene Encoding the Colletotrichum gloeosporioides Regulatory Subunit of Protein Kinase A. Journal of Biological Sciences, 8: 730-737.
DOI: 10.3923/jbs.2008.730.737
URL: https://scialert.net/abstract/?doi=jbs.2008.730.737
DOI: 10.3923/jbs.2008.730.737
URL: https://scialert.net/abstract/?doi=jbs.2008.730.737
INTRODUCTION
Colletotrichum gloeosporioides is a plant pathogenic fungus of a wide variety of crops in the subtropical and tropical regions. It is known to be the causal organism of anthracnose diseases that bring damages to plants of economic importance such as avocado, mango, citrus, strawberry and papaya (Timmer et al., 1998; Peres et al., 2002; Sanders and Korsten, 2003; MacKenzie et al., 2007). These diseases cause significant post harvest crop loss and limit export quality of the fruits produced. In response to host specific signals, the conidium of C. gloeosporioides germinates to form a germ tube followed by differentiation into a highly specialized infection cell, the appressorium. The appressorium then produces an infection peg which penetrates the plant cuticle and the pectin layer beneath (Deising et al., 2000). Upon entering the host, it will produce a network of internal hyphae to further penetrate and degrade plant cells. Development of appressoria is a complex morphogenetic process involving multiple signals, both physical and chemical (Estrada et al., 2000; Kim et al., 2000).
Signal transduction pathways are crucial in regulating fungal morphogenetic events and the cAMP-dependent protein kinase A (PKA) pathway is known as one of the major pathways mediating most of the morphological effects in pathogenic fungi (Lengeler et al., 2000). Components of this cascade are conserved in a variety of fungi. cAMP is produced from ATP by adenylate cyclase and acts as a secondary messenger to regulate the activity of the cAMP-dependent protein kinase A (PKA) (Borges-Walmsley and Walmsley, 2000). In Saccharomyces cerevisiae, at low levels of cAMP, the PKA holoenzyme is an inactive tetramer which comprises of two regulatory and two catalytic subunits. When cAMP levels increase, cAMP binds to the regulatory subunit and induces conformational change that causes the dissociation of the tetramer to regulatory subunit dimer and two active catalytic subunits. The released catalytic subunits become active and phosphorylate the target substrates which compose of metabolic enzyme and transcriptional factor (D`Souza and Heitman, 2001).
Genes encoding the regulatory subunits of PKA have been isolated and cloned from various filamentous fungi such as Ustilago maydis, Neurospora crassa and C. lagenarium (Gold et al., 1994; Bruno et al., 1996; Takano et al., 2001). Each of them has been identified to regulate fungal development. Mutation in the regulatory subunit of PKA in U. maydis affects morphogenesis, resulting in a constitutive budding growth pattern (Gold et al., 1994). In N. crassa, a temperature-sensitive mutant of the PKA regulatory subunit showed a complete loss of growth polarity and mislocalization of septa when grown at the restrictive temperature (Bruno et al., 1996). A mutant of the gene encoding the PKA regulatory subunit (rpk1) of C. lagenarium showed significant reduction in vegetative growth and conidiation and unable to generate penetration hyphae that leads to decrease in virulence (Takano et al., 2001).
To understand the role of the cAMP-dependent protein kinase A in C. gloeosporioides, in this research, we report on the isolation and the characterization of the CgPKAR encoding the regulatory subunit of PKA from C. gloeosporioides. The nucleotide sequence representing the gene and its corresponding cDNA were isolated and characterized. We analyzed the expression of CgPKAR transcripts in different C. gloeosporioides morphological forms and showed that this gene is differentially expressed in different growth morphologies.
MATERIALS AND METHODS
Fungal strains and culture conditions: C. gloeosporioides strain PeuB (Farah Diba, 2003) was used throughout this study. C. gloeosporioides cultures were grown by subculturing on Potato Dextrose Agar (PDA; Oxoid, United Kingdom) at 30°C for 7 days. To obtain fungal mycelia, the fungus was grown in Potato Dextrose Yeast Extract (PDYE) broth at 30°C for three days with shaking at 180 rpm. Escherichia coli strain DH5α was used for plasmid propagation. E. coli was grown in LB broth at 37°C for overnight.
Isolation of C. gloeosporioides CgPKAR: The Cg-PKAR was isolated using a Polymerase Chain Reaction (PCR)-based strategy. The primers were designed based on the conserved regions of several fungal genes encoding the Protein kinase A regulatory subunit. These primers are the RSF (forward) 5` CGT CGA ACC TCG GTC TCT GCC GAG 3` (RRTSVSAE) and the RSR2 (reverse) 5` GCT AGC GGC GCG AGG CGC ATC ATT 3` (NDAPRAAS). The PCR was performed on C. gloeosporioides genomic DNA. The PCR reaction was performed with one cycle at 94°C for 5 min, followed by 30 cycles at 94°C for 1 min, 56.1°C for 1 min and 72°C for 2 min. These cycles were followed by an extension cycle of 20 min at 72°C. Subsequently, the purified PCR products were cloned into cloning vector, pGEM-T Easy vector (Promega, USA) and sequenced. Following sequence information, 5` and 3` Rapid Amplification of cDNA Ends (RACE) PCR were performed using SMARTTM RACE cDNA amplification kit (Clontech, USA) following the protocol supplied by the manufacturer. Sequences of the primers used for the amplification, REGF and REGR, are given in Table 1. The amplified product were cloned into pGEM-T Easy vector and sequenced. To isolate the 5` regulatory region of the gene, DNA walking strategy using the DNA Walking SpeedUpTM kit (Seegene, Korea) was used. Three sequence specific primers designated as TSP1, TSP2 and TSP3 (Table 1) were used in the amplification reaction according to manufacturer instructions. The amplicons obtained were cloned and sequenced.
Genomic DNA blot hybridization: Total DNA of C. gloeosporioides was isolated from mycelia using polyvinylpyrrolidone (PVP) as described by Pich and Schubert (1993). DNA digestion, agarose gel fractionation, labeling of probes and hybridization were performed according to the manufacturer`s instructions and standard methods (Sambrook and Russel, 2001). DNA probes were labeled with [α-32P] dCTP with the kit Ready To GoTM DNA Labeling Beads (-dCTP) (Amersham, USA). Hybridization was carried out with hybridization buffer [1 M Na2HPO4.2H2O, 1 M NaH2PO4, 0.5 M EDTA, 0.1% (w/v) SDS] at 65°C for 4 h for pre-hybridization and hybridized overnight after the labeled-probes were added. The membrane was washed at 65°C with 2X SSC for 10 min followed by 2X SSC and 0.1% SDS; 1X SSC and 0.1% SDS; 0.5X SSC and 0.1% SDS until the radioactivity signal was low.
| Table 1: | Oligonucleotide primers used in this study |
 | |
Sample preparation for RNA extraction: Conidia of C. gloeosporioides strain PeuB were taken from 7 day old culture grown on PDA. To isolate the conidia, fungal mycelia were first scrapped off the plate. Subsequently 1 mL of sterile water was added onto the plate and the conidia were scrapped gently from the agar into the water. The conidia were pelleted from the suspension by centrifugation at 4000 rpm at room temperature for 5 min. To obtain germinated conidia, 1x106 spores mL-1 of freshly washed conidia were suspended in sterile water and induced for germination in 50% papaya extract at 30°C with shaking at 190 rpm. After more than 70% of the conidia germinated, usually 7 h after the inoculation, conidia were collected by centrifugation at 4000 rpm for 5 min at room temperature. Both the conidia and germinating conidia were immediately frozen in liquid nitrogen and if not used immediately, they were stored at -80°C until further usage. To obtain C. gloeosporioides appressoria, 70 mL of conidia suspension (1x106 spores mL-1) were poured onto glass plates (150 mm) that were coated with an appressoria inducer, the plant wax, extracted from rubber leaves (Nurhaida et al., 2007). After 6 h of induction, when more than 80% of the conidia germinated into appressoria, the suspension covering the glass plate was poured off and the surface of the plates were briefly rinsed with deionised water to remove some of the spores that failed to adhere and differentiate.
RNA extraction and RT-PCR optimization: Total RNA of conidia, germinating conidia and mycelia were extracted using TRI REAGENT® solution (Molecular Research Center, USA) while RNA from the appressoria was extracted using guanidine isothiocyanate in combination with mechanical cell disruption by glass beads (Nurhaida et al., 2007). Integrity and yield of the RNA was tested by agarose gel electrophoresis. Prior to cDNA synthesis, RNA samples were purified with Clean Up RNeasy (Qiagen, Germany) dan treated with RNase-free DNase (Qiagen). Subsequently, the purified RNA was used as template in reverse transcriptase reaction (RT-PCR) using Access® RT-PCR kit (Promega) following the manufacturer`s protocol.
SYBR green real-time RT-PCR assays: SYBR green real-time RT-PCR assays was conducted by iCycler iQ™ Real-Time PCR Detection System (Bio-Rad, USA). Detection of real-time RT-PCR products was by binding of the flourescent DNA dye, SYBR green. The iScriptTM One-Step RT-PCR kit with SYBR® Green (Bio-Rad) was used in which the cDNA synthesis and PCR amplification were carried out in the same tube. All assays were carried out in triplicates and appropriate controls were included (PCR reactions without template). All tubes were heated for 10 min at 50°C for cDNA synthesis followed by 5 min at 95°C for iScript Reverse transcriptase inactivation. Subsequently, each tube was subjected to a PCR amplification protocol of 94°C for 2 min followed by 44 cycles of 10 sec at 94°C and 30 sec at 60°C (data collection). Melt curve analysis was performed immediately after the amplification protocol under the following conditions: 1 min denaturation at 95°C, 1 min annealing at 55°C, 80 cycles of 0.5°C increments (10 sec each) beginning at 55°C (data collection). All PCR products were electrophoresed on agarose gel (1%) to verify amplifications. Assays were repeated with independently isolated total RNA and subsequently synthesized cDNA samples. PCR fragments were cloned and sequenced to confirm that only the target sequence was amplified. Melt curve profile was also analysed. Melt curve is one of the methods to measure the melting temperature of double stranded DNA (dsDNA) and its major application is to determine the number of amplified product by looking at the peak produced by the curve. Expression of the CgPKAR in different cDNA samples was compared to the level of its expression in the reference sample, which is the cDNA from mycelia. Thus, expression of the regulatory subunit gene in mycelial cDNA sample was assigned the value of 1.0. Amplification efficiency of CgPKAR and 18S rDNA were shown to be equivalent, allowing the use of comparative Ct method for relative quantification as described by Livak and Schmittgen (2001). Relative gene expression of CgPKAR was analysed using 2-ΔΔCT method as described by Livak and Schmittgen (2001).
RESULTS
A partial fragment of the gene encoding C. gloeosporioides regulatory subunit of protein kinase A was obtained by PCR amplification using primers designed based on amino acids in conserved regions of the protein. Partial sequence of 830 bp of the PKA regulatory gene was cloned and sequenced (data not shown). Based on this partial sequence, specific primers were designed to isolate the full length gene and cDNA of C. gloeosporioides regulatory subunit of PKA, designated as CgPKAR, by amplification of 5` and 3` flanking regions using DNA walking and RACE PCR strategies.
The full length CgPKAR and its cDNA were amplified and sequenced. The full-length sequence encompassing the 5` and 3` flanking regions of CgPKAR is 2186 bp. CgPKAR consists of a 1241 bp open reading frame and by comparing the cDNA sequence with the sequence of the corresponding gene, a single intron of 65 bp was identified (Fig. 1). The cDNA encodes for a 391 amino acid protein with a putative molecular mass of 42 kDa. The size of the CgPKAR ORF was similar to PKA regulatory genes of S. cerevisiae, 1251 bp (Toda et al., 1987), N. crassa, 1158 bp (Bruno et al., 1996), Magnaporthe grisea, 1173 bp (Adachi and Hamer, 1998), Colletotrichum trifolii, 1215 bp (Yang and Dickman, 1999) and Candida albicans, 1380 bp (Cassola et al., 2004). The deduced amino acid sequence of CgPKAR also shares significant homology with those of the PKA regulatory genes: C. lagenarium, RPK1 (90% identity), M. grisea, SUM1 (71%), N. crassa, mcb (69%) dan C. trifolii, Ct-PKAR (61%) (Fig. 2). A putative TATA and fungal CAAT boxes were found upstream from the start codon at positions -120 and -288, respectively. The CgPKAR gene sequence has been submitted to the GenBank with an accession number DQ673616.
 | |
| Fig. 1: | Nucleotide and deduced amino acid sequences of CgPKAR. GenBank accession No. for CgPKAR is DQ673616. The predicted TATAA and CAAT sequences are colored in grey boxes. The start (ATG) and stop codons (TAA) are underlined (__). Intron sequences are indicated in lowercase |
 | |
| Fig. 2: | Amino acid alignment of CgPKAR with other fungal PKA regulatory subunits and indication of conserved domains found in the protein. Kinase inhibitor domain, a highly conserved six residue sequence (RRTSVSAE) containing an autophosphorylation site (Ser-91) is underlined in full. Two cAMP-binding domains, CBD-A (-206 to -222) and CBD-B (-327 to -344) are represented by dotted lines. Accession number: C. gloeosporioides (C. gloeos) (ABG66306), C. lagenarium (C. lagena) (AAK31209); C. trifolii (C. trifol) (AAC04356); M. grisea (AAC34140); N. crassa (AAB00121) |
 | |
| Fig. 3: | Southern blot analysis of C. gloeosporioides genomic DNA. DNA (8 μg/lane) was digested with Pst1 (P), Kpn1 (K), HindIII (H), EcoR1 (E) and BamH1 (B). The blot was probed with a 830 bp partial fragment of CgPKAR |
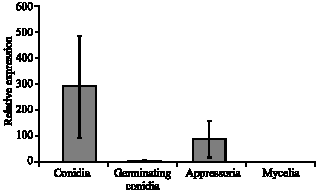 | |
| Fig. 4: | Expression level of CgPKAR in different morphological cells; conidia, germinating conidia, appressoria and mycelia. 18SrDNA was used as a reference gene and the expression of CgPKAR in different morphological cells were compared to the level of its expression in the reference sample (mycelia) |
Southern blot analysis with genomic DNAs digested with BamH1, EcoR1, HindIII, KpnI and PstI indicates that CgPKAR is a single-copy gene in the genome of C. gloeosporioides (Fig. 3).
In RT-PCR optimisation, single PCR products for both CgPKAR (78 bp) and 18S rDNA (101 bp) were amplified using different morphological cells (data not shown). The optimal annealing temperatures for CgPKAR and 18S rDNA were 56 and 61°C, respectively. The positive control (amplification using primers and template that were supplied with the kit) showed the expected result while negative control (using specific primers without cDNA template) showed no amplification. Hence, all these optimized conditions were used in real-time RT-PCR.
The melt curve profile was generated by iCycler iQ™ System. The total fluorescence produced by the binding of SYBR Green I to dsDNA during temperature changes was recorded. The -dF/dT plot showed the rate of significant fluorescence and melting temperature of the PCR product. The melt curve graph of real-time RT-PCR assay involving CgPKAR and 18S rDNA showed one curve with only one peak, respectively (data not shown). The specificity of the primers was proven to amplify a single PCR product of CgPKAR and 18S rDNA at melting temperatures of 80 and 85°C, respectively.
Relative expression of CgPKAR was found highest in conidia with 234-fold, appressoria with 72-fold and germinating conidia with 4-fold as compared to mycelia (reference sample) (Fig. 4). Expression level of CgPKAR in different morphological cells was normalized with the expression level of 18S rDNA prior to relative comparison to mycelia (reference sample). Reading from the mycelia was chosen as reference sample because of its lowest expression between all different morphological forms. The real-time RT-PCR was repeated three times, with three technical replicates for each run and all results showed consistency in the expression data of CgPKAR.
DISCUSSION
The subunit regulatory PKA gene from C. gloeosporioides (CgPKAR) was cloned and characterised. It is present as a single copy in C. gloeosporioides genome and contains one intron, similar to RPK1 of C. lagenarium (Takano et al., 2001) and SUM1 of M. grisea (Adachi and Hamer, 1998). Two out of three putative general features of regulatory subunit PKAs were found on CgPKAR (Fig. 2). First is the presence of the kinase inhibitor domain, which is a highly conserved six residue sequence, RRTSVSAE (88-95) containing an autophosphorylation site (Ser-91). This domain acts as an inhibitor of catalytic subunit kinase activity and also mediates interaction between the regulatory and catalytic subunits (Bruno et al., 1996). Second is the presence of two cAMP-binding domains, which are CBD-A (-206 to -222) and CBD-B (-327 to -344). Each CBD has the conserved amino acid sequence required for the formation of helix subdomains and eight α-barrel chain for the binding of cAMP. An important feature of the β-barrel is a conserved Phosphate-Binding Cassette (PBC) (Canaves and Taylor, 2002), which is also conserved in CgPKAR (Fig. 2). The deduced amino acid sequence also suggests that CgPKAR of C. gloeosporioides is most closely related to type II regulatory subunit, showing an overall identity to type II PKA-R genes: 71% with M. grisea (Adachi and Hamer, 1998), 69% with N. crassa (Bruno et al., 1996), 61% with C. trifolii (Yang and Dickman, 1999) and 59% with A. niger (Staudohar et al., 2002). Given the similarity to other regulatory subunit proteins and the structural features of the protein, C. gloeosporioides CgPKAR most likely belongs to type II PKA regulatory subunit. However, we found that CgPKAR lacks a dimerisation domain which is normally located in the N-terminal one-third of the protein. Dimerisation domain has been shown to mediate dimer formation between two regulatory subunits and interaction with other cellular proteins (Bruno et al., 1996). Thus, the structure of the C. gloeosporioides PKA holoenzyme is most likely heterodimeric that composes of only one catalytic and one regulatory subunit. From the sequence similarity and conservation of domains found in previously studied regulatory subunits of PKA, we conclude that CgPKAR encodes C. gloeosporioides regulatory subunit of PKA.
Real-time RT-PCR assay was performed to determine the relative quantification of regulatory subunit gene expression in different morphological cells using SYBR green as a measurement of PCR product formation. How CgPKAR is regulated during conidia-appressoria morphogenesis is still unknown. Results of this work indicate that the expression of CgPKAR is developmentally regulated at least at the level of transcription. CgPKAR expression levels oscillated during different growth stages, with conidia and appressoria accumulating high levels of CgPKAR transcripts while the germinating conidia and mycelia showing low levels of expression. In C. gloeosporioides, conidia that germinate in rich media form mycelia and enter saprophytic life cycle (Barhoom and Sharon, 2004). However, in the absence of rich media and in the presence of plant and hard surface signals, the fungus enters the parasitic life cycle, where the conidia germinate and form appressoria surrounded by a thick melanized cell wall. Appressoria swells as a result of increase in internal turgor pressure and force the penetration peg into the plant tissues, followed by invasive growth of the fungus. Barhoom and Sharon (2004) showed that exogenous cAMP is required for conidia saprophytic germination and for appressorium formation. However the pathogenic-specific spore germination is cAMP-independent process. Thus, CgPKAR may be expressed abundantly in conidia to regulate the next developmental stages which are the formation of germ tube and appressoria. CgPKAR is also highly expressed in the appressoria as it is needed to help the fungal differentiate and form penetration peg. In M. grisea and C. lagenarium, the cAMP-PKA pathway was shown to be involved in lipid degradation and this metabolism produces glycerol that is thought to generate appressorial turgor pressure (Thines et al., 2000; Yamauchi et al., 2004; Wang et al., 2005). Based on the expression data, CgPKAR could also play a similar role in C. gloeosporioides as the gene is expressed relatively high in appressoria. The reason why CgPKAR is expressed lowly in germ tube and mycelia could be because cAMP-dependent PKA pathway is not actively involved in the cellular development during saprophytic growth such as hyphal elongation. The fungus may employ other signal transduction pathways to regulate genes required for hyphal growth and elongation. These results suggest that the PKA regulatory subunit gene undergoes regulation at the transcriptional level involving developmental changes of the C. gloeosporioides conidia and appressoria.
ACKNOWLEDGMENTS
This research project is funded by the Ministry of Science, Technology and Innovation, Malaysia under the IRPA grant 09-02-02-0105-EA257. Nurhaida Kamaruddin is supported by Universiti Pendidikan Sultan Idris, Perak, Malaysia.
REFERENCES
- Adachi, K. and J.E. Hamer, 1998. Divergent cAMP signaling pathways regulate growth and pathogenesis in the rice blast fungus Magnaporthe grisea. Plant Cell, 10: 1361-1374.
Direct Link - Barhoom, S. and A. Sharon, 2004. cAMP regulation of pathogenic and saprophytic fungal spore germination. Fungal Genet. Biol., 41: 317-326.
Direct Link - Borges-Walmsley, M.I. and A.R. Walmsley, 2000. cAMP signaling in pathogenic fungi: Control of dimorphic switching and pathogenicity. Trends Microbiol., 8: 133-141.
Direct Link - Bruno, K.S., R. Aramayo, P.F. Minke, R.L. Metzenberg and M. Plamann, 1996. Loss of growth polarity and mislocalization of septa in a Neurospora mutant altered in the regulatory subunit of cAMP-dependent protein kinase. EMBO J., 15: 5772-5782.
Direct Link - Canaves, J.M. and S.S. Taylor, 2002. Classification and phylogenetic analysis of the cAMP-dependent protein kinase regulatory subunit family. J. Mol. Evol., 54: 17-29.
Direct Link - Cassola, A., M. Parrot, S. Silberstein, B.B. Magee, S. Passeron, L. Giasson and M.L. Cantore, 2004. Candida albicans lacking the gene encoding the regulatory subunit of protein kinase A displays a defect in hyphal formation and an altered localization of the catalytic subunit. Eukaryot. Cell, 3: 190-199.
CrossRefPubMedDirect Link - D'Souza, C.A. and J. Heitman, 2001. Conserved cAMP signaling cascades regulate fungal development and virulence. FEMS. Microbiol. Rev., 25: 349-364.
Direct Link - Deising, H.B., S. Werner and M. Wernitz, 2000. The role of fungal appressoriain plant infection. Microb. Infect., 216: 1631-1641.
Direct Link - Estrada, A.B., J.C. Dodd and P. Jeffries, 2000. Effect of humidity and temperature on conidial germination and appressorium development of two Philippine isolates of the mango anthracnose pathogen Colletotrichum gloeosporioides. Plant Pathol., 49: 608-618.
CrossRefDirect Link - Gold, S.E., G. Duncan, K. Barrett and J.W. Kronstad, 1994. cAMP regulates morphogenesis in the fungal pathogen Ustilago maydis. Genes Dev., 8: 2805-2816.
CrossRef - Kim, Y.K., Z.M. Liu, D. Li and P.E. Kolattukudy, 2000. Two novel genes induced by hard-surface contact of Colletotrichum gloeosporioides conidia. J. Bacteriol., 182: 4688-4695.
Direct Link - Lengeler, K.B., R.C. Davidson, C. D'Souza, T. Harashima, W. Shen, P. Wang, X. Pan, M. Waugh and J. Heitman, 2000. Signal transduction cascades regulating fungal development and virulence. Microbiol. Mol. Biol. Rev., 64: 746-785.
Direct Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - MacKenzie, S.J., T.E. Seijo, D.E. Legard, L.W. Timmer and N.A. Peres, 2007. Selection for pathogenicity to strawberry in populations of Colletotrichum gloeosporioides from native plants. Phytopathology, 97: 1130-1140.
CrossRef - Nurhaida, K., A.B. Farah Diba, A.R. Rohaiza, N.M. Mahadi and A.M.A. Murad, 2007. Rapid isolation of total RNA from conidia, germinating conidia and appressoria of the fungal plant pathogen Colletotrichum gloeosporioides. Sains Malaysiana, 36: 91-95.
Direct Link - Peres, N.A.R., E.E. Kuramae, M.S.C. Dias and N.L. de Souza, 2002. Identification and characterization of Colletotrichum spp. affecting fruit after harvest in Brazil. J. Phytopathol., 150: 128-134.
CrossRefDirect Link - Pich, U. and I. Schubert, 1993. Midiprep method for isolation of DNA from plants with a high content of polyphenolics. Nucleic Acids Res., 21: 3328-3328.
Direct Link - Sanders, G.M. and L. Korsten, 2003. Comparison of cross inoculation potential of South African avocado and mango isolates of Colletotrichum gloeosporioides. Microbiol. Res., 158: 143-150.
CrossRefDirect Link - Staudohar, M., M. Bencina, P.J.I. Van de Vondervoort, H. Panneman, M. Legisa, J. Visser and G.J.G. Ruijter, 2002. Cyclic AMP-dependent protein kinase is involved in morphogenesis of Aspergillus niger. Microbiology, 148: 2635-2645.
Direct Link - Takano, Y., K. Komeda, K. Kojima and T. Okuno, 2001. Proper regulation of cyclic AMP-dependent protein kinase is required for growth, conidiation and appressorium function in the anthracnose fungus Colletotrichum lagenarium. Am. Phytopathol. Soc., 14: 1149-1157.
CrossRefDirect Link - Thines, E., R.W. Weber and N.J. Talbot, 2000. MAP kinase and protein kinase A-dependent mobilization of triacylglycerol and glycogen during appressorium turgor generation by Magnaporthe grisea. Plant Cell, 12: 1703-1718.
Direct Link - Timmer, L.W., G.E. Brown and S.E. Zitko, 1998. The role of Colletotrichum spp. in postharvest anthracnose of citrus and survival of C. acutatum on fruit. Plant Dis., 82: 415-418.
CrossRefDirect Link - Toda, T., S. Cameron, P. Sass, M. Zoller, J.D. Scott, B. McMullen, M. Hurwitz, E.G. Krebs and M. Wigler, 1987. Cloning and characterization of BCY1, a locus encoding a regulatory subunit of the cyclic AMP-dependent protein kinase in Saccharomyces cerevisiae. Mol. Cell. Biol., 7: 1371-1377.
CrossRefDirect Link - Wang, Z.Y., J.M. Jekinson, L.J. Holcombe, D.M. Soanes, C. Veneault-Fourrey, G.K. Bhambra and N.J. Talbot, 2005. The molecular biology of appressorium turgor generation by the rice blast fungus Magnaporthe grisea. Biochem. Soc. Trans., 33: 384-388.
Direct Link - Yamauchi, J., N. Takayanagi, K. Komeda, Y. Takano and T. Okuno, 2004. cAMP-PKA signaling regulates multiple steps of fungal infection cooperatively with Cmk1 MAP kinase in Colletotrichum lagenarium. Mol. Plant Microbe. Interact., 17: 1355-1365.
Direct Link - Yang, Z. and M.B. Dickman, 1999. Molecular cloning and characterization of Ct-PKAR, a gene encoding the regulatory subunit of cAMP-dependent protein kinase in Colletotrichum trifolii. Arch. Microbiol., 171: 249-256.
CrossRefDirect Link