
J.W. Muthomi
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
P.E. Otieno
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
G.N. Chemining wa
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
J.H. Nderitu
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
J.M. Wagacha
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 7 | Page No.: 1163-1170







ABSTRACT
Greenhouse experiments were conducted over two cropping cycles to investigate the effect of fungicide seed treatment and fungal root rot pathogens on nodulation and dry matter accumulation of selected food legumes. The legumes were common bean (Phaseolus vulgaris L. variety GLP 2), green gram (Vigna radiata L. variety M66) and lablab (Lablab purpureus L.) while the pathogens were Fusarium oxysporum f. sp. phaseoli, Macrophomina phaseolina, Sclerotinia sclerotiorum and Rhizoctonia solani. Treatments consisted of inoculation of legume seeds with appropriate rhizobia alone, rhizobia together with fungicide, rhizobia together with pathogen and a combination of rhizobia, fungicide and pathogen. Fungicide copper oxychloride was used as a seed dresser. Rhizoctonia solani and S. sclerotiorum were more pathogenic and showed significantly increased seedling mortality and greater reduction in seedling emergence, number of nodules and root dry matter. Fungicide seed dressing significantly increased seedling emergence and reduced seedling mortality. However, fungicide seed dressing alone and in combination with pathogen depressed nodulation in all the legumes. Inoculation with F. oxysporum and M. phaseolina had no significant effect on nodulation in common bean. All the treatments had little or no significant effect on shoot dry matter. The results suggest that fungicide seed treatment in combination with rhizobia inoculation is beneficial in management of root rot and enhancement of nodulation in food legumes.
PDF Abstract XML References Citation
How to cite this article
J.W. Muthomi, P.E. Otieno, G.N. Chemining wa, J.H. Nderitu and J.M. Wagacha, 2007. Effect of Legume Root Rot Pathogens and Fungicide Seed Treatment on Nodulation and Biomass Accumulation. Journal of Biological Sciences, 7: 1163-1170.
DOI: 10.3923/jbs.2007.1163.1170
URL: https://scialert.net/abstract/?doi=jbs.2007.1163.1170
DOI: 10.3923/jbs.2007.1163.1170
URL: https://scialert.net/abstract/?doi=jbs.2007.1163.1170
INTRODUCTION
Grain legumes constitute next to maize the most important group of food crops in the diet in Kenya. They form a major source of proteins of high biological value, energy, minerals and vitamins for many people in tropical Africa where main diets consist mostly of starchy staples and minimal animal protein (Taylor et al., 2005). The protein content of edible species of legumes, with exception of soybeans and groundnuts, varies between 18 and 32% (FAO, 2006). However, dry seed yields in most African countries have been declining due to diseases, low soil fertility, insect pests and low, erratic rainfall (Blackshaw, 2007; Belle, 2006; Miklas et al., 2006; Tefera, 2005; Heard, 2004; Guene et al., 2003; Singh et al., 2003; Araujo et al., 2000; Otsyula et al., 1998; Nderitu et al., 1997; Allen, 1990). Commercial fertilizers are expensive and out of reach for most small-scale farmers. Therefore, cheap sources of nitrogen like biological nitrogen fixation, combined with better pest and disease management options need to be optimized if yields are to be sustained and food security attained. The fact that legume crops are generally cultivated in poor environments re-emphasizes the need for biological nitrogen fixation.
All agriculturally important legume species have the ability to symbiose with Rhizobium and fix free atmospheric nitrogen. World wide, some 44 to 66 million tonnes of nitrogen are fixed annually, providing nearly half of all the nitrogen used in agriculture (Giller, 2001). The amount of nitrogen fixed by legumes varies with species and may supply a major part or the entire nitrogen needed by a crop (Russo, 2006; Chui et al., 2003). Reduced dependence on nitrogen fertilizers and attention to farming practices that favour the more economically viable and environmentally friendly nitrogen fixation through legume- Rhizobium symbiosis has a special relevance to the developing countries (Bünemann et al., 2006; Giller, 2001; Obara et al., 2000). Symbiotically-fixed nitrogen has been considered as a useful source of nitrogen to non-fixing plants in intercropping systems (Shaharoona et al., 2006; Guene et al., 2003; Carsky et al., 1997; Hudgens, 1996; Sanginga et al., 1994; Fujita et al., 1992).
Root rot diseases are a major limiting factor in legume production. The diseases depress seedling germination and cause post emergence damping off, resulting in poor crop stand and low yields. The disease causal agents are seed borne but most farmers often use seeds saved from previous harvest, a practice that negates the principle of sanitary practices (Buruchara, 1990). Effective control strategies against root rot fungal pathogens have not been fully developed. Sanitation and use of clean planting material is the primary way of preventing damping-off and other root diseases. Chemical seed treatment before planting is a common practice to prevent seed and seedling rots, damping off and other fungal diseases. However, problems arise when the chemical seed treatments are to be used in conjunction with rhizobia inoculants. In some cases the applied seed fungicide may fail to protect against the intended pathogen or suppresses the effectiveness of the rhizobia bacteria. For the last three decades, studies on compatibility of rhizobial strains with fungicides have been controversial. Application of Captan, Pentachloronitrobenzene (Curley and Burton, 1975) and Apron (Revellin et al., 1993), on soybean (Glycine max) seeds reduced the viability of Bradyrhizobium japonicum by 18, 75 and 61%, respectively, after one hour exposure. Graham et al. (1980) observed that less than 10% of R. phaseoli strains survived on Thiram-treated seeds of common bean. By contrast, no detrimental effect was found on the compatibility of Apron with R. japonicum applied to soybean seeds (Diatloff, 1986) or with R. meliloti on alfalfa seeds (Edmisten et al., 1988). Hashem et al. (1997) observed differences in compatibility with fungicides between peanut (Arachis hypogaea) and Bradyrhizobium inoculants. Geune et al. (2003) reported varying effects on nodulation and nitrogen fixation of common bean by treatment with Dichlorofenothion-thiram based on the Rhizobium strain used.
Therefore, this study was carried out to investigate the interaction of fungicide seed treatment and rhizobia inoculation in the management of fungal root rots and their effect on nodulation and dry matter accumulation of selected food grain legumes.
MATERIALS AND METHODS
Pathogen isolation and inoculum production: Root or hypocotyl portions of legume plants showing symptoms of root rot were collected from the University of Nairobi’s Field Station, Kabete. Tissue portions were surface sterilized and aseptically plated on Potato Dextrose Agar (PDA) medium at room temperature (22±3°C) for 7-14 days. Pure cultures were obtained by hyphal tip transfer of each of the colonies onto fresh PDA media. Identification of the pathogens was done using cultural (colour of aerial mycelium, pigmentation) and morphological characteristics (septation of hyphae, conidia shape and size, septation of conidia). Inoculum was prepared by growing each pathogen separately on PDA for 7-14 days at room temperature (22±3°C). Whole cultures of each pathogen were macerated with sterile distilled water in a blender to make a slurry using sterile distilled water.
Experimental design and treatments: Greenhouse experiments were conducted over two cropping cycles in 2006 at the University of Nairobi’s Faculty of Agriculture, Kenya. The grain legumes used were common bean (Phaseolus vulgaris L.var GLP 2), green gram (Vigna radiata L.) and lablab (Lablab purpureus L.). Treatments included, inoculation with pathogens (Fusarium oxysporum f. sp. phaseoli, Macrophomina phaseolina, Sclerotinia sclerotiorum, Rhizoctonia solani), or with appropriate rhizobia or application of fungicide (copper oxychloride) and their combinations. Prior to inoculation and fungicide treatment, the seeds were surface-sterilized in 10% sodium hypochlorite and rinsed twice in sterile distilled water. Common bean was inoculated with bean rhizobia while green gram and lablab were inoculated with cowpea cross-nodulating rhizobia strain. Slightly wetted seeds were thoroughly mixed with the appropriate rhizobia inoculant. Sucrose was added to the mixture to enable the rhizobia stick to the seed surface and also to offer initial nutrients for the bacteria. Fungicide treatment was done by mixing seeds with the copper oxychloride powder. Pathogen slurry (15 mL) containing 5x105 propagules per milliliter for each of the pathogens was mixed into steam-sterilized soil in plastic pots and five appropriately treated seeds were then placed on the surface of the soil and covered with 1 cm layer of soil. Watering was done regularly as required using distilled water. The experiment was arranged in a randomized complete block design and each treatment was replicated four times.
Assessment of root rot, nodulation and dry matter accumulation: Root rot was assessed on the basis of percent seedling emergence, seedling mortality and plant dry weight. Percent seedlings emergence was recorded after germination while percent seedling mortality was recorded on the second, fourth and sixth week after emergence. The experiment was terminated on the sixth week after emergence. The potting medium was carefully washed off the roots and the number of nodules counted for each plant. The plant were then separated into roots and shoots and dried in an oven at 110°C for 48 h for dry weight determination. Data was subjected to analysis of variance (ANOVA) using Genstat software and the treatment means separated using tukey LSD test at 5% probability level.
RESULTS
Seedling emergence and mortality: The interaction between the treatments and legume species on seedling emergence was not significant (p = 0.05) (Table 1). However, significant differences were observed among the treatments and legume species. Rhizoctonia and Sclerotinia spp. significantly reduced seedling emergence but Fusarium and Macrophomina spp. showed no significant effect. Fungicide application significantly increased seedling emergence on Rhizoctonia and Sclerotinia inoculated seeds. Green gram showed significantly lower emergence compared to common bean and lablab. Treatments, legume species and their interaction had a significant (p = 0.05) effect on the percent seedling mortality (Table 2). Fungicide application significantly reduced seedling mortality in Rhizoctonia and Sclerotinia treated seeds by as much as 57% but fungicide seed treatment had little effect on Fusarium and Macrophomina treated seeds except in common bean. However, inoculation with Rhizoctonia and rhizobia combined with fungicide significantly reduced seedling mortality on all the three legumes.
| Table 1: | Mean percentage emergence of legumes treated with fungicide and inoculated with different root rot fungi and rhizobia inoculant |
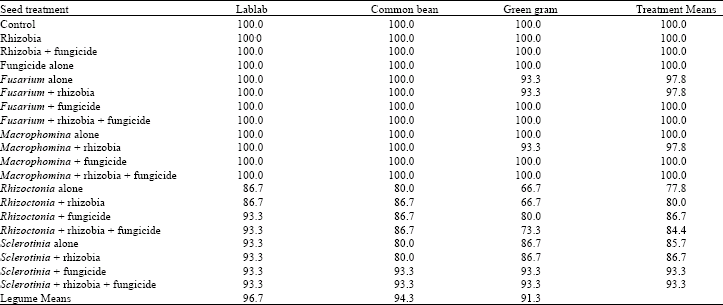 | |
| LSD (p≤0.05) Legumes = 2.7; Treatments = 7.0; Treatments x Legumes = Not significant | |
| Table 2: | Mean percentage mortality at the 6th week after emergence for three legume species treated with fungicide and inoculated with different root rot fungi and rhizobia inoculant |
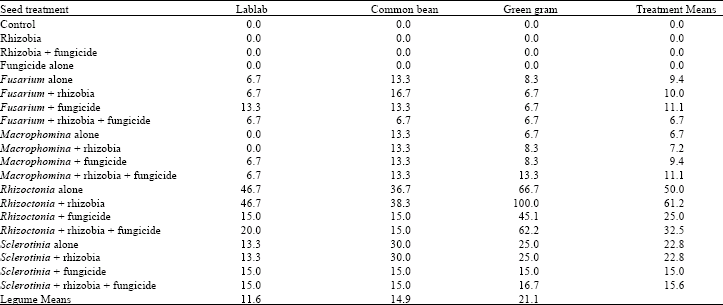 | |
| LSD (p≤0.05) Legumes = 4.9; Treatments =12.6; Treatments x Legumes = 21.8 | |
Number of nodules per plant: The interaction between the treatments and the legume species on number of nodules per plant was significant (p = 0.05). Rhizobia inoculation significantly increased the number of nodules per plant in all the legume species except green gram, which showed no effect (Table 3). However, combination of Fusarium and rhizobia with fungicide and rhizobia had no significant effect on nodulation in lablab but it significantly reduced the number of nodules per plant in the other treatments. In addition, application of fungicide on rhizobia inoculated seeds significantly reduced the number of nodules per plant in common bean but had no effect in lablab and green gram.
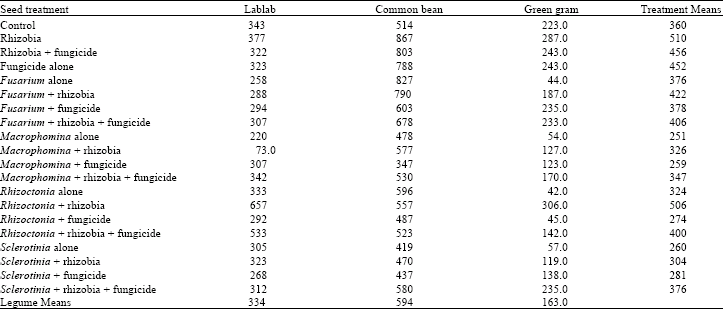
Dry matter accumulation: The interaction between treatments and legume species on shoot and root dry matter was not significant (Table 4 and 5). However, significant (p = 0.05) differences were observed among the treatments and legume species for both shoot and root dry matter. All the pathogens had no effect on shoot dry matter of the legume species. Rhizobia inoculation alone significantly increased shoot dry matter but not when applied in conjunction with the pathogens and fungicide. All the pathogens had no significant effect on root dry matter although Macrophomina, Sclerotinia and Rhizoctonia spp. showed reduced root dry matter. Fungicicide application and its combination with pathogens or with rhizobia had no effect on root dry matter.
| Table 3: | Mean number of nodules per plant of legumes treated with fungicide and inoculated with different root rot fungi and rhizobia inoculant at 6 weeks after emergence |
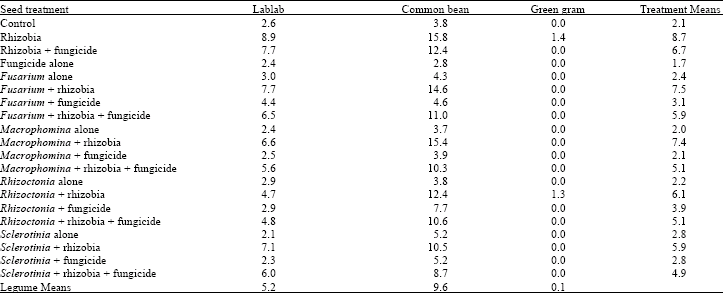 | |
| LSD (p≤0.05) Legumes = 0.7; Treatments = 1.8; Treatments x Legumes = 3.1 | |
| Table 4: | Mean shoot dry weight per plant (g) of three legume species treated with fungicide and inoculated with different root rot fungi and rhizobia inoculant at 6 weeks after emergence |
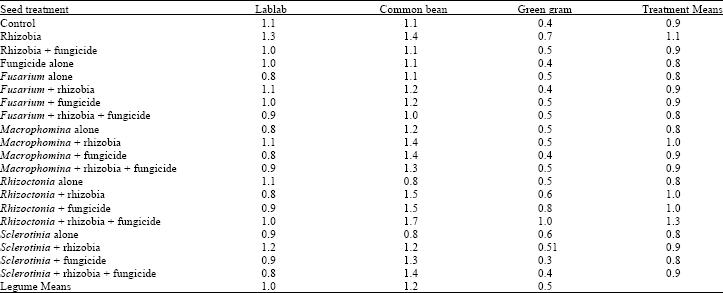 | |
| LSD (p = 0.05): Legumes 0.1; Treatments 0.2; Treatments x legumes – Not significant | |
| Table 5: | Mean root dry weight per plant (mg) of three legume species treated with fungicide and inoculated with different root rot fungi and rhizobia inoculant at 6 weeks after emergence |
 | |
| LSD (p = 0.05): Legumes 64.8; Treatment 167.0; Treatment x Legume = Not significant | |
DISCUSSION
Inoculation of the legumes with Rhizoctonia and Sclerotinia spp. significantly reduced seedling emergence and increased mortality whereas Fusarium and Macrophomina spp. had little or no effect. This indicated that Rhizoctonia and Sclerotinia spp. were more pathogenic at the conditions provided during the experiment. Similar findings were reported by Wong et al. (2003). The insignificant effect of Fusarium and Macrophomina spp. may be due to the unfavourable conditions characterized by moderate temperature and high moisture content provided during the experiment. Fusarium and Macrophomina spp. thrive well under moisture stress and high temperatures but disease development is reduced in flooded soils (Kirkpatrick et al., 2006; Nischwitz et al., 2004; Afouda, 1999; Ratnoo et al., 1997).
Inoculation with rhizobia improved nodulation and shoot dry matter. However, fungicide application significantly reduced disease incidence but negatively impacted on nodulation. Valenciano et al. (2004) working with Phaseolus vulgaris made a general observation that pesticides accelerated bean emergence by allowing seed protection as well as improving germination. Various researchers have reported reduction of nodules or the number of viable rhizobia on the seed with fungicide application. Kyei-Boahen et al. (2001) reported significant reduction in nodulation, percent nitrogen derived from the atmosphere and shoot dry matter by seed dressing fungicides Apron®, Arrest 75W®, Crown® and Captan® in chick pea. Arrest® and Captan® decreased nodule dry matter and percent nitrogen derived from the atmosphere while Arrest® reduced dry matter yield. In a recent study by Stovold and Evans (2006), seed dressers Thiram®, Captan® and Captafol® provided consistent protection of soybean against seedling diseases at rates of 2000 ppm a.i w/wor 3000 ppm a.i. w/w but had inhibited nodulation. Other fungicide seed dressers with negative effect on nodulation and growth of Rhizobia include carbendazim, carboxin, mancozeb, chloraniliprodione, metalaxyl and thirum (Rennie and Dubetz, 1984; Revellin et al., 1993; Castro et al., 1997). Niewladomska and Klama (2005) reported reduction of nitrogenase activity of Rhizobium leguminosarum bv. trifolii, Sinorhizobium melilotii and Bradyrhizobium sp. ornithopus bacteria on clover, lucerne and siratro, respectively, due to the effects of seed-dressing with the fungicide Furaben®. Dichlorofenthion-thiram® completely inhibited nodulation of non-nodulating soybean treated with Rhizobium etli and significantly decreased nodulation (but not nitrogen fixation) when seeds were treated with R. tropici (Guene et al., 2003).
A seed dresser may also act as a temporary barrier for gaseous exchange between seeds and environment resulting in phytotoxicity (Valenciano et al., 2004). These reports by different researchers on the effect of seed dressers on nodulation and shot dry matter weight indicate a pattern of specificity in interaction among seed dressers, rhizobia strain and the test legume plant but not a generalized interaction. The influence of a particular seed dresser on nodulation and performance of a Rhizobium strain seem to be dependent on their interaction, which determines their compatibility.
When fungicide was used in combination with rhizobia, the disease incidence was significantly reduced and nodulation increased compared to when applied alone. This indicates that simultaneous use of rhizobia and fungicides for root rot management and nodulation enhancement is beneficial. However, the significantly higher nodulation of the legumes treated with rhizobia alone than a combination of rhizobia and fungicide raises concern on the effect of the seed dresser on nitrogen fixation. Studies to determine fungicides that are less toxic to rhizobia and their optimum concentration ought to be done. For instance, concentration of Thiram beyond 500 μg mL-1 has been reported to be highly toxic with respect to plant growth factors and rhizobial infection to Glycine max (Bikrol et al., 2005). However, nodulation, nodule dry weight, nitrogenese activity were observed to be maximum at 100 μg mL-1 of Thiram.
The negative effect of copper oxychloride on nodulation in this study indicates that it had some bactericidal effects on the rhizobia. This concurs with the findings of other workers. Heweidy et al. (2005) reported that copper oxychloride was the most inhibiting to bradyrhizobial strains, even though it significantly decreased the infection percentage with Macrophomina phaseolina, Fusarium oxysporum and Sclerotium rolfisii compared to other tested fungicides. Higher nodulation and nitrogen fixation emanating from rhizobia have been suggested based on the method of application of the rhizobium strain. Whereas Hawthorne et al. (2006) generally suggested that fungicides and rhizobium should never be mixed together before application to seed, other workers have suggested specific methods. Stovold and Evans (2006) recommended use of water suspension of peat-based rhizobial inoculant or sand particles coated with peat inoculant being introduced to the furrow together with the seed as effective methods of inoculation of soybean and pea. Bogino et al. (2006) reported in-furrow inoculation with Bradyrhizobium as producing increased nodule number relative to seed inoculation on peanut.
The results suggest that copper oxychloride seed treatment is beneficial in the management of root rots of legumes caused by Rhizoctonia solani and Sclerotinia sclerotiorum but it is not effective on Fusarium oxysporum f.sp. phaseoli and Macrophomina phaseoli. The fungicides can be applied together with rhizobia to enhance nodule formation and therefore biological nitrogen fixation in food legumes. The study also found out that seed dressing plays an important role in enhancing seedling emergence while reducing seedling mortality. However, there is need for more studies on the optimal use of fungicides in combination with rhizobia in relation to nodulation and root rot management under field conditions. Optimum fungicide concentration and most effective method of fungicide-rhizobia application in relation to various legumes ought to be studied further. Developing fungicide resistant rhizobial strains remains one approach to overcome the current constraint of delivering inoculants.
ACKNOWLEDGMENT
This research was carried out through the financial support from the Deans Committee research grant of the University of Nairobi, Kenya.
REFERENCES
- Araujo, A.P., M.G. Teixeira and D.L. Almeida, 2000. Growth and yield of common bean cultivars at two soil phosphorous levels under biological nitrogen fixation. Pesq. Agropec. Bras., 35: 809-817.
Direct Link - Bikrol, A., N. Saxena and K. Singh, 2005. Response of Glycine max in relation to nitrogen fixation as influenced by fungicide seed treatment. Afr. J. Biotechnol., 4: 667-671.
Direct Link - Blackshaw, R.F., L.J. Molnar, G.W. Clayton, K.N. Harker and T. Entz, 2007. Dry bean production in zero and conventional tillage. Agron. J., 99: 122-126.
Direct Link - Carsky, R.J., R. Abaidoo, K.E. Dashiell and N. Sanginga, 1997. Effect of soybean on subsequent maize grain yield in the Guinea savanna zone of West Africa. Afr. Crop Sci. J., 5: 31-38.
Direct Link - Castro, S., M. Vinocur, M. Permigiani, C. Halle, T. Taurian and A. Fabra, 1997. Interaction of the fungicide in culture and under field conditions. Biol. Fertil. Soils, 25: 147-151.
Direct Link - Fujita, K., K.G. Otsu and S. Ogta, 1992. Biological nitrogen fixation in mixed legume-cereal cropping systems. Plant Soil, 141: 155-175.
Direct Link - Guene, N.F.D., A. Diouf and M. Gueye, 2003. Nodulation and nitrogen fixation of field grown common bean (Phaseolus vulgaris) as influenced by fungicide seed treatment. Afr. J. Biotechnol., 2: 198-201.
Direct Link - Hashem, F.M., S.A. Saleh, P. Van Berkum and M. Voll, 1997. Survival of Bradyrhizobium sp. (Arachis) on fungicide treated peanut seed in relationship to plant growth and yield. World J. Microbiol. Biotechnol., 13: 335-340.
Direct Link - Kirkpatrick, M.T., J.C. Rupe and C.S. Rothrock, 2006. Soybean response to flooded soil conditions and the association with soilborne plant pathogenic genera. Plant Dis., 90: 592-596.
Direct Link - Kyei-Boahen, S., A.E. Slinkard and F.L. Walley, 2001. Rhizobial survival and nodulation of chickpea as influenced by fungicide seed treatment. Can. J. Microbiol., 47: 585-589.
Direct Link - Miklas, P.N., J.D. Kelly, S.E. Beebe and M.W. Blair, 2006. Common bean breeding for resistance against biotic and abiotic stresses: From classical to MAS breeding. Emphytica, 147: 105-131.
Direct Link - Niewladomska, A. and J. Klama, 2005. Pesticide side effect on the symbiotic efficiency and nitrogenase activity of Rhizobiaceae bacteria family. Poland J. Microbiol., 54: 43-48.
Direct Link - Nischwitz, C., M. Olsen and S. Rasmussen, 2004. Effect of irrigation type on inoculum density of Macrophomina in melon fields in Arizona. J. Phytopathol., 152: 133-137.
Direct Link - Otsyula, R.M., S.I. Ajanga, R.A. Buruchara and C.S. Wortmann, 1998. Development of an integrated bean root rot control strategy for Western Kenya. Afr. Crop Sci. J., 6: 61-68.
Direct Link - Ratnoo, R.S., K.L. Jain and M.K. Bhatnagar, 1997. Effect of atmospheric temperature on the development of ashy stem blight of cowpea. J. Mycol. Plant Pathol., 27: 90-91.
Direct Link - Revellin, C., P. Leterme and G. Catroux, 1993. Effect of some fungicide seed treatments on the survival of Bradyrhizobium japonicum and on the nodulation and yield of soybean (Glycine max L. Merr). Biol. Fertil. Soils, 16: 211-214.
Direct Link - Russo, V.M., 2006. Mineral nutrient and protein contents in tissues and yield of navy bean, in response to nitrogen fertilization and row spacing. J. Food Agric. Environ., 4: 168-171.
Direct Link - Sanginga, N., S.K.A. Danso, K. Mulongoy and A.A. Ojeifo, 1994. Persistence and recovery of introduced Rhizobium ten years after inoculation on Leucaena leucocephala grown on an Alfisol in Southwestern Nigeria. Plant Soil, 159: 199-204.
CrossRefDirect Link - Shaharoona, B., M. Arshad and Z.A. Zahir, 2006. Effect of plant growth promoting rhizobacteria containing ACC-deaminase on maize (Zea mays L.) growth under axenic conditions and on nodulation in mung bean (Vigna radiata L.). Lett. Applied Microbiol., 42: 155-159.
Direct Link - Singh, S.P., H. Teran, C.G. Munoz, J.M. Osorno, J.C. Takegami and M.D.T. Thung, 2003. Low soil fertility tolerance in landraces and improved common bean genotypes. Crop Sci., 43: 110-119.
Direct Link - Stovold, G.E. and J. Evans, 2006. Fungicide seed dressings: Their effects on emergence of soybean and nodulation of pea and soybean. Aust. J. Exp. Agric., 20: 497-503.
Direct Link - Taylor, S.R., B.D. Weaver, W.C. Wood and S.V. Edzard, 2005. Nitrogen application increases yield and early dry matter accumulation in late-planted soybean. Crop Sci. J., 45: 854-858.
Direct Link - Tefera, T., 2006. Effect of common bacterial blight severity on common bean yield. Trop. Sci., 46: 41-44.
Direct Link - Wong, D.H., N.J. Barbetti and K. Sivasithamparam, 2003. Effect of soil temperature and moisture in the pathogenecity of fungi associated with roots of subterranean clovers. Aust. J. Agric. Res., 35: 675-684.
Direct Link - Valenciano, J.B., P.A. Casquero and J.A. Boto, 2004. Influence of sowing techniques and pesticide application on the emergence and the establishment of bean plants (Phaseolus vulgaris L.). Agronomie, 24: 113-118.
Direct Link