
Dongwei Li
School of Biological and Food Engineering, Fuyang Normal University, Fuyang 236037 Anhui, China
LiveDNA: 86.35819
Meixia Cao
School of Biological and Food Engineering, Fuyang Normal University, Fuyang 236037 Anhui, China
Yongmin Li
School of Biological and Food Engineering, Fuyang Normal University, Fuyang 236037 Anhui, China
Changqing Qu
School of Biological and Food Engineering, Fuyang Normal University, Fuyang 236037 Anhui, China
International Journal of Pharmacology
Year: 2021 | Volume: 17 | Issue: 3 | Page No.: 103-112







ABSTRACT
Background and Objective: Berberine is well known as a traditional anti-bacterial and anti-inflammatory Chinese medicine. Recently, it was reported to have interesting effects on adipogenic differentiation, glucose uptake, insulin resistance, etc. However, the molecular mechanisms of these underlying pharmacological effects are not sufficiently elucidated. Here, the mechanism of berberine on adipogenic differentiation in porcine intramuscular preadipocytes was investigated. Materials and Methods: Preadipocytes were obtained for primary culture from the healthy piglets under sterile conditions. Preadipocytes viability was detected by MTT assay. Intracellular lipid droplets were observed under microscopy by oil-red O staining and the extraction quantified by colorimetry at room temperature. The key adipogenic genes were detected by real-time quantitative PCR. Luciferase assays were used to validate the targeting relationship between microRNA and the MAPK signalling pathway. Results: When porcine preadipocytes were induced into adipogenic differentiation, cell proliferation and accumulation of intracellular lipid droplets were decreased in a time- and dose-dependent manner by berberine. Berberine attenuated adipogenic differentiation by impairing some key adipogenic genes expression, such as PPARγ and adiponectin and even provoking comprehensive effects through the interference with the interaction of a microRNA (miR-143) and certain a target gene (MAP2K5). Conclusion: The inhibiting effects of berberine on adipogenic differentiation in porcine intramuscular preadipocytes are through the multi-gene pattern, which provides novel evidence for treatments of berberine to gain healthy animal-based foods against excessive fatty deposits, even human obesity, hyperlipidemia, diabetes, etc.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Dongwei Li, Meixia Cao, Yongmin Li and Changqing Qu, 2021. Regulatory Effects of Berberine on Adipogenic Differentiation in Porcine Intramuscular Preadipocytes Through the Multi-Gene Pattern. International Journal of Pharmacology, 17: 103-112.
DOI: 10.3923/ijp.2021.103.112
URL: https://scialert.net/abstract/?doi=ijp.2021.103.112
DOI: 10.3923/ijp.2021.103.112
URL: https://scialert.net/abstract/?doi=ijp.2021.103.112
INTRODUCTION
High-fat diets obtain from domestic animals in modern society, especially for porcine productions, which easily induce human obesity. It is reported that obesity gives rise to slight chronic systemic inflammation, which is treated acceptably as a complex disorder of energy metabolism, associated with some serious public health problems involving hyperlipidemia, hypercholesterolemia, atherosclerosis, diabetes, hypertension, endocrine maladjustment and other multi factorial chronic diseases1,2. Obesity results considerably from the pathological development of abnormal adipocytes group, involving excessive recruit from precursor cells, increasing of adipocyte size and lipid droplets3. The process of adipogenesis, in which there are multi stage responses to several kinds of hormonal signals such as glucocorticoids and insulin, is undoubtedly characterized by the regulation of symbolic adipogenic genes such as peroxisome proliferator-activated receptors (PPARs), CCAAT/enhancer-binding proteins (C/EBPs). Moreover, adipose tissue is an active endocrine and paracrine organ that secreted some adipocytokines and bioactive mediators, including adiponectin which is secreted only from adipose tissue and inversely correlated with obesity and associated complications4. Details of adipogenesis differentiation are obscure, which is preconditioned to understand the regulatory mechanism of fatty deposit of human and animals. Today, some artificial-synthesized drugs and therapeutic strategies for obesity and correlated diseases are extensively applied, but subsistent defects are difficult to be ignored, so the study of effective treatment modalities is still essential.
Berberine is an isoquinoline alkaloid mostly from Coptidisrhizoma, which is usually used as a traditional Chinese medicine without major side effects for the treatment of bacterial diarrhea, hyperlipidemia, inflammation, atherosclerosis, diabetes, obesity and other diseases5-9. In addition, previous studies demonstrate that berberine could reduce triglyceride accumulation and block adipogenesis of 3T3-L1 cells through regulating some symbolic genes of lipid metabolism, such as PPARs, C/EBPs10-14. The expression of adiponectin is inhibited by berberine, which is abolished by knockdown of AMP-activated protein kinase α1 (AMPKα1)15. Furthermore, it was reported that the anti-adipogenic effect of berberine has been associated with some non-coding RNAs (ncRNAs). Among the long ncRNAs that were acquired from the hepatic expression profiles in the diet-induced steatotic model with berberine treatment by micro array analysis, MRAK052212 and MRAK080926 were strongly linked to triglyceride synthesis in lipid metabolism16 and in 3T3-L1 cells treated with berberine, microRNA (miRNA) sequence analyses revealed a decrease of miR-92a expression and that increased RNA-binding motif protein 4a (RBM4a) expression, which expedited the beige adipogenesis17.
It is well known that miRNAs are involved in the regulatory network of numerous biological processes, such as cell proliferation, cell differentiation and energy homeostasis18-20. Some circulating miRNAs have been identified as candidate biomarkers of physiologic status and abnormal expressions of miRNAs should be implicated in pathological conditions21. Usually, one miRNA could target diverse mRNAs, which coordinate or antagonize functions of each other through the post-transcriptional gene regulatory network. It has been reported that some of the miRNAs may interact with berberine by multiple regulation mechanism, including multiples signal pathways22. Mitogen-activated protein kinase (MAPK) signaling pathways are important and conservative in diversified essential biochemical process, including cells proliferation, differentiation and development, which conventionally comprise extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK) and p38 MAPK. There are miRNAs regulating adipogenesis through MAPK signaling pathways23-25. Berberine could exert pharmacological effects by MAPK signaling pathways26,27. For instance, berberine could activate ERK and JNK28,29.
To sum up, berberine plays an interesting role in lipid metabolism through multiple gene modulations and their interactions. While fat deposits, as a quantitative trait, is regulated multiplying a genetic regulatory network. In this study, the effect of berberine on adipogenic differentiation in porcine intramuscular preadipocytes by the multi-gene pattern was investigated.
MATERIALS AND METHODS
Study duration and location: This study project was performed from October, 2019 to January, 2021 in the School of Biological and Food Engineering, Fuyang Normal University, Fuyang, China.
Materials: Dulbecco's modified Eagle's medium/hamF12 (DMEM/F12), fetal bovine serum (FBS), phosphate-buffered saline (PBS) and HEPES were purchased from Gibco Life Technology (Grand Island, NY, USA). Bovine serum albumin (BSA), type I collagenase, 3-isobutyl-1-methylxanthine, dexamethasone, Rosiglitazone, insulin, berberine, TRIzol reagent, oil-red O, MTT, DMSO, penicillin and streptomycin were purchased from Sigma-Aldrich Ltd. (Saint Louis, MO, USA). A PrimeScript™ reverse transcription (RT) reagent kit was purchased from Takara Biotechnology Corporation (Dalian, China) and other real-time quantitative PCR reagents, miR-143 mimics, miR-143 inhibitor and negative control oligonucleotide were purchased from GenePharma Co. Ltd. (Shanghai, China). The Dual-Glo luciferase assay system was purchased from Promega (Madison, WI, USA). Bradford protein assay reagent and protease inhibitor were purchased from Sino Gene (Beijing, Chima). Primary antibodies, including anti-TGFβ, anti-MAP2K5, anti-β-actin and goat anti-rabbit antibody were purchased from Abcam (Wales, England).
Preadipocyte culture and adipogenic differentiation: The porcine subcutaneous pre adipocytes were obtained for primary in vitro culture from the healthy piglets under sterile conditions in the local livestock farm. All treatments of animals were carried out according to the Guide for the Care and Use of Laboratory Animals. Primary in vitro culture of preadipocytes and differentiation was carried out according to the methods previously described30. In brief, adipose tissue was sliced from the back of piglets and rinsed immediately with the basal medium of DMEM/F12 containing 100 mmol L–1 HEPES and 50 U mL–1 penicillin-streptomycin. Then, the tissue was cut into pieces and digested with 1 mg mL–1 type I collagenase in the basal medium complemented with 20 mg mL–1 BSA for about 1 hr under the condition of 37 shaking water bath, followed by filtration through 70 nylon sieve mesh and centrifugation at 1000 rpm for 10 min. Subsequently, the Stromal Vascular Fraction (SVF) pellets were collected and suspended in the complete medium, which consisted of the basal medium and 10% FBS. Preadipocytes were isolated from SVF pellets and seeded in culture dishes at a density of 5×104 per square centimeter in a 5% CO2 damp atmosphere at 37. Under normal conditions, these fibroblast-like preadipocytes were cultured in the complete medium. When preadipocytes reached approximately 90% confluence, these cells were passaged or induced into adipogenic differentiation by the treatment with the complete medium complemented with 0.1 μmol L–1 3-isobutyl-1-methylxanthine, 1.0 μmol L–1 dexamethasone, 0.1 μmol L–1 Rosiglitazone, 0.2 μmol L–1 insulin for 48 hrs. Then, the medium was converted to the complete medium supplemented with1.0 μmol L–1 dexamethasone and 0.2 μmol L–1 insulin to maintain adipogenic differentiation. In special experiments, the cells were treated with different concentrations of berberine (5, 10 and 20 μmol L–1).
Preadipocytes transfected with oligonucleotides: Preadipocytes were resuspended in a serum-free medium before transfection. The cell suspension (104 per μ, 300 μL volume) was incubated with 100 nM miR-143 mimic, miR-143 inhibitor or control oligonucleotide for 5 min on ice and then twice electroporated for 9 msec at 260 V by Electro Square Porator (BTX, ECM2001, SanDiego, CA). The cells were resuspended in 10 mL of pre warmed medium and then seeded on culture plates in a 37 incubator for subsequent assays.
MTT assay: Cell viability was detected by MTT assay. Preadipocytes were cultured at 1×104 per well in the complete medium. When the cells reached approximately 90% confluence, the cells treated with different concentration of berberine (0, 5, 10 and 20 μmol L–1) for 48 hrs. The cells treated with 0.1% DMSO were used for controls. PBS containing 5 mg mL–1 MTT was then added into each well and incubated for 3 hrs. The result ant formazan product was dissolved by about 0.1 mL DMSO and the Optical Density (OD) was measured at 540 nm by a microplate reader.
Oil-red O staining extraction assay: Intracellular lipid droplets, accumulated at mature period, were observed under the microscopy by oil-red O’staining and the extraction quantified by colorimetry at room temperature. Firstly, cells were washed three times in PBS and then fixed in 4% formaldehyde for 30 min. Secondly, the fixed cells were quickly rinsed in PBS and then stained with filtered 1% oil-red O solution for 15 min. After the oil-red O solution was removed, the cells were gently rinsed with distilled water and then instantly destained with 100% isopropanol solution for 15 min. The OD was detected at 490 nm by ultraviolet spectrophotometry.
Real-time quantitative PCR (RT-qPCR): The total RNA was extracted with a TRIzol reagent and the qualified mRNA was reversely transcribed with random primers to synthesize the first-strand cDNA by the PrimeScriptTM Reverse Transcription (RT) reagent kit. The real-time qPCR reactions were conducted in a final volume of 20 μLin triplicate using SYBR-Green with an ABI 7500 system (Applied Biosystems). The reactions were performed as follows: The initial incubation at 95 for 30 sec, 40 cycles at 95 for 5 sec and 60 for 31 sec. The coding genes were normalized to the level of β-actin mRNA and the small nuclear U6 was amplified as an endogenous control for the miRNA. The relative expression level of each gene of interest was calculated by the method of the 2–Ct. The primers and probes were as follows: PPARγ, 5 -GTGAAGGATGCAAGGGTT-3 (forward) and 5-TGATGGCGTTATGAGACA-3 (reverse); adiponectin, 5-TAGGAGGTAAGGTTGGAGAT-3 (forward) and 5-GTGATGTGGAAGGAGAAGTA-3 (reverse); MAP2K5, 5-TTCTC TTCACCACCTTCAC-3 (forward) and 5-CATCTACTAGCAG CACTACA-3 (reverse); β-actin, 5-GCCAACCGTGAGAAGATG-3 (forward) and 5-CAGAGGCGTACAGGGACA-3 (reverse); miR-143, 5-GTCGTATCCAGTGCGTGTCGTGGAGTCGGCAATTGCA CTGGATACGACTGAGCT-3 (RT primer), 5-GGCTGAGATG AAGCACTGT-3 (gene-specific stem-loop primer) and 5-CAG TGCGTGTCGTGGAG-3 (probe); U6, 5-TTCTCATCTCA GCGTTCAG-3 (forward) and 5-TGCCAGTCCACAGTAAGA-3 (reverse).
Dual-glo luciferase reporter assay: MAP2K5 was predicted as a target gene of miR-143 by the MicroCosm and miR and a target prediction tool, which then was experimentally validated by the Dual-Glo Luciferase assay system. The 3-untranslated region (3 -UTR) sequence of MAP2K5 mRNA was from the National Centre for Biotechnology Information and the sequence of miR-143 was from miRBase, which were used for the construction of EGFP/RFP reporter vectors. The miR-143 mimics or negative control was co-transfected into 293T cells with the reporter vector (pMIR-MAP2K5 3-UTR or pMIR-MAP2K5-knockout3-UTR). The EGFP/RFP reaction activities were examined after 2 days of the transfection by the F4500 fluorescence spectrophotometer.
Western blot: Preadipocytes were washed with PBS and then transferred into the total protein extract containing below 1 mL protease inhibitor. The protein concentration in the lysate was examined by the Bradford protein assay reagent. The total proteins were separated by SDS-PAGE and transferred to PVDF membranes, which were incubated with primary antibodies (anti-β-actin, anti-TGFβ or anti-MAP2K5). Then, the membranes were processed with goat anti-rabbit antibody for overnight immunoblot. The immunoreactivity was visualized by enhanced chemiluminescence (ECL, Engreen).
Statistical analysis: The data were presented as Means±SEM. One-way analysis of variance (ANOVA) was used for two groups in a single experiment and multiple comparisons. Individual comparisons were assessed by Student s t-test. The value of p<0.05 was regarded to be statistically significant.
RESULTS
Effect of berberine on intracellular triglyceride accumulation and cell viability: Numerous intracellular triglyceride droplets inmature adipocytes may be observed by oil red O staining after adipogenic differentiation. It was found that fibroblast-like preadipocytes could scarcely trigger adipogenic differentiation (Fig. 1a), which were not influenced by berberine (Fig.1b). In addition, the degree of adipogenic differentiation could be significantly changed in a dose- and time-dependent manner by berberine (Fig. 1c-f, Fig. 2a-b). When the gradient concentration of berberine was increased, intracellular triglyceride content was significantly decreased in the constant incubation time. As the cultivating time extended, intracellular triglyceride content in adipocytes was significantly decreased under the same concentration of berberine. The proliferation of preadipocytes was significantly decreased, as the concentration of berberine was increased. Even so, the downward trend of cells propagation could be partly offset before preadipocytes proliferation reached a plateau (Fig. 2c). In short, berberine may impair intracellular triglyceride accumulation and preadipocytes viability.
Effect of berberine on mRNA expressions of PPARγ and
adiponectin: To investigate the effect of berberine on adipogenesis at the gene transcription level, the mRNA expression of several key representative adipogenic genes were analyzed by RT-qPCR. PPARγ is the master regulator of adipogenesis and adiponectin is an adipocyte-specific secreted protein which is induced during preadipocyte differentiation. The mRNA expression levels of PPARγ and adiponectin were examined to confirm the further effects of berberine on the differentiation of preadipocytes (Fig. 3a-b). As a result, berberine may inhibit the gene transcription of PPARγ and adiponectin, especially in high concentrations and further impact on adipogenesis.
MiR-143 regulates adipogenic differentiation by targeting
the 3-UTR of MAP2K5: Since berberine has effects on expressions of coding genes related to adipogenesis, its effects on the interaction of non-coding genes and coding genes are interesting to dig deep into the question. As a focused RNA type of non-coding genes, miRNAs have effects on biological processes by targeting the specific seed region of objective genes. It was well reported that miR-143 is thought to be involved in adipogenic differentiation. MAP2K5 from the MAPK signaling pathway was estimated to be a target gene of miR-143 by bioinformatics algorithms, such as the DIANA microT-CDS, MicroCosmormiR and a prediction program. Furthermore, MAP2K5 was confirmed to be the target gene of miR-143 by Dual-Gloluciferase reporter assay and western blot analysis. Our results showed that MAP2K5 mRNA and protein levels were significantly decreased in the miR-143 mimics group compared with those in the untreated, empty-vector-transfected or MAP2K5 mutant groups, which bases the binding-site were knockout in the 3-UTR of MAP2K5 (Fig. 4a-b).
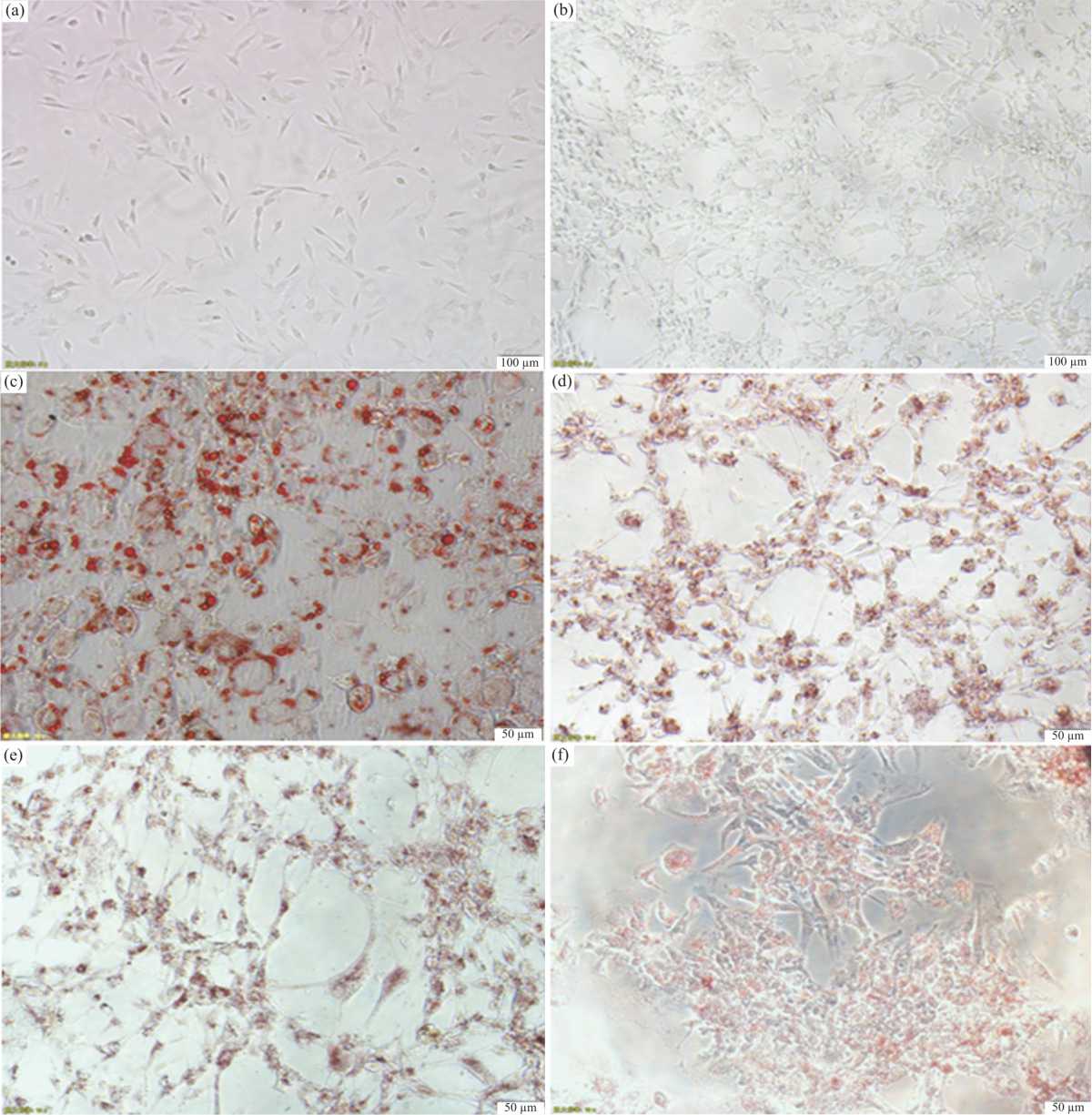 |
| Fig. 1(a-f): | Effect of berberine on adipogenic differentiation was observed by oil red O staining and cell morphology (a) Fibroblast-like preadipocytes without any treatment in the complete medium. Bar, 100 μm. (b) Fibroblast-like preadipocytes treated with 5 μmol L–1 berberine for 48 hrs after cultivating 7 days in the complete medium. Bar, 100 μm. (c) Morphological changes of preadipocytes after 7 days of adipogenic differentiation. Intracellular lipid droplets with oil-red O staining were visualized. Bar, 50 μm. (d) Morphological changes of preadipocytes treated with 5 μmol L–1 berberine for 48 hrs after 7 days of adipogenic differentiation. Bar, 50 μm. (e) Morphological changes of preadipocytes treated with 10 μmol L–1 berberine for 48 hrs after 7 days of adipogenic differentiation. Bar, 50 μm and (f) Morphological changes of preadipocytes treated with 20 μmol L–1 berberine for 48 hrs after 7 days of adipogenic differentiation. Bar, 50 μm |
These indicated that miR-143 may regulate adipogenic differentiation of preadipocytes by targeting MAP2K5 via the MAPK signalling pathway.
Effect of berberine on expression of the MiR-143 and its targeting MAP2K5: The expression of miR-143 was maintained at a low level before the early stage of adipogenic differentiation but subsequently began to significantly increase during adipogenic differentiation (Fig. 5a). Meanwhile, the expression of MAP2K5 was down-regulated from clonal expansion to terminal differentiation (Fig. 5b).
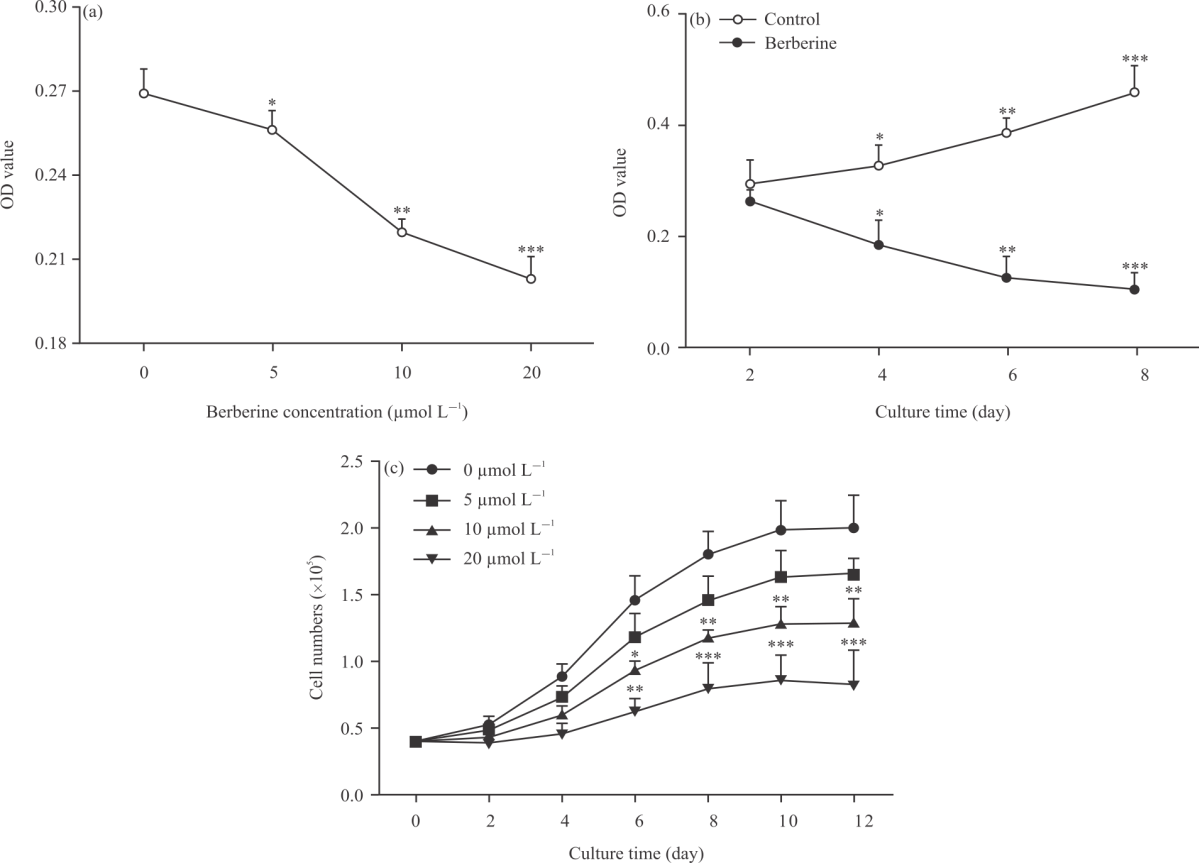 |
| Fig. 2(a-c): | Effect of berberine on intracellular triglyceride accumulation or cell viability which was measured by oil red O staining extraction or MTT assay, respectively (a) Effect of berberine on intracellular triglyceride content was determined in a dose-dependent manner after 0, 5, 10, 20 μmol L–1 berberine treatments for 48 hrs after 3 days of adipogenic differentiation, respectively. (b) Effect of berberine on intracellular triglyceride content was determined in a time-dependent manner after 10 μmol L–1berberine treatments for 2, 4, 6, 8 days, respectively and (c) Growth curves of preadipocytes treated with 0, 5, 10, 20 μmol L–1 berberine, respectively. All data are shown as Mean±SEM. *p<0.05, **p<0.01, ***p<0.001 |
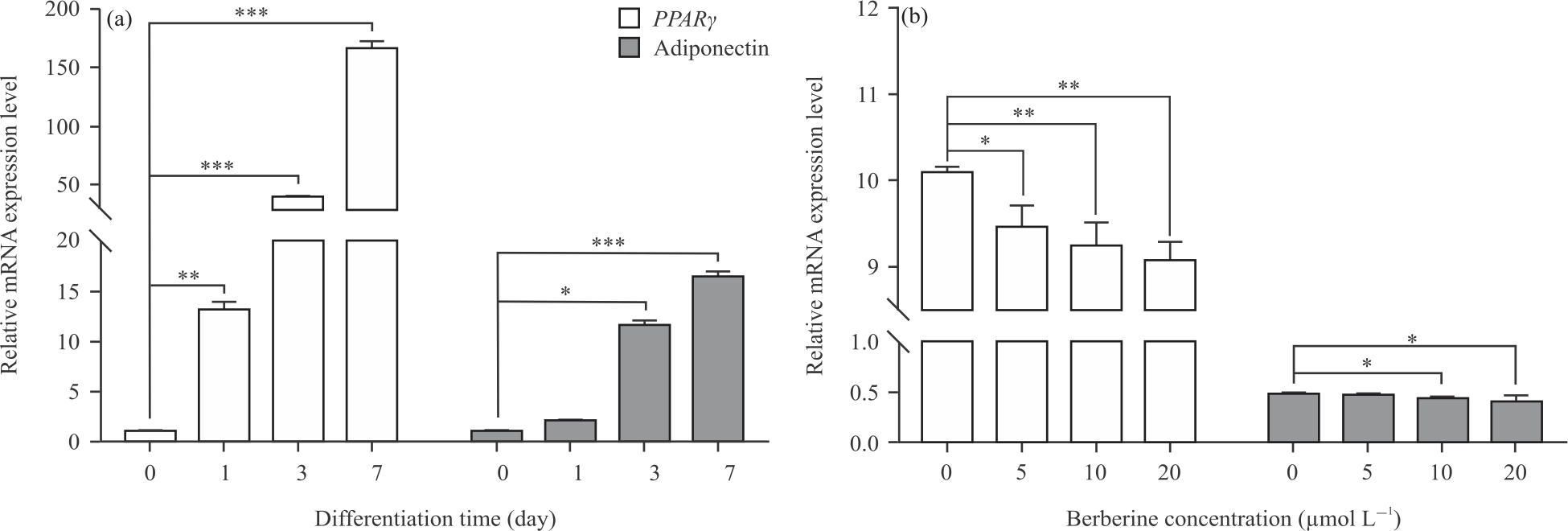 |
| Fig. 3(a-b): | Relative mRNA expressions of PPARγ and Adiponectin during adipogenic differentiation were examined by RT-qPCR (a) Relative mRNA expressions of PPARγ and Adiponectin after 0, 1, 3, 7 days of adipogenic differentiation and (b) Relative mRNA expressions of PPARγ and Adiponectin in preadipocytes treated with 0, 5, 10, 20 μmol L–1 berberine for 48 hrs after 3 days of adipogenic differentiation, respectively. All data are shown as Means±SEM. *p<0.05, **p<0.01, *p<0.001 |
 |
| Fig. 4(a-b): | MiR-143 regulates adipogenic differentiation by targeting the 3 -UTR of MAP2K5 (a) miRNA-143 was verified to target 3-UTR of MAP2K5 by dual-luciferase reporter assay. Cells were co-transfected with the pMIR empty vector, the pMIR-MAP2K5 wild type plasmid or the pMIR-MAP2K5 mutant plasmid, either alone or in combination with the miR-143 mimics. The fluorescence values were measured with an F-4500 fluorescence spectrophotometer after 48 hrs and (b) MAP2K5 protein levels evaluated by western blot analysis after 1 day of adipogenic differentiation. miR-143: miR-143 mimics, Anti-miR-143: miR-143 inhibitor, Vector alone: Empty vector-transfected |
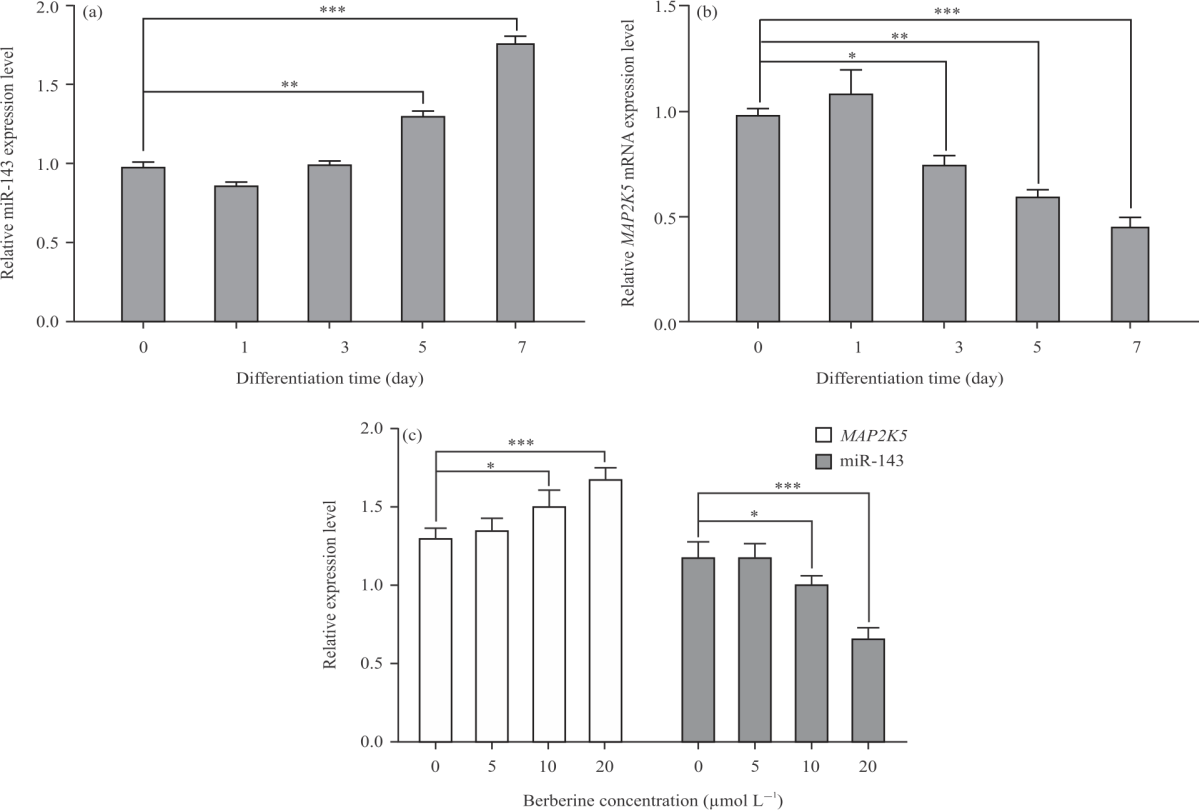 |
| Fig. 5(a-c): | Expression of miR-143 and MAP2K5 without or with the treatment of berberine (a) Expression levels of miR-143 were examined during adipocytes differentiation by RT-qPCR, (b) Expression levels of MAP2K5 were examined during adipocytes differentiation by RT-qPCR and (c) Expression levels of miR-143 and MAP2K5 mRNA in preadipocytes treated with 0, 5, 10, 20 μmol L–1 berberine for 48 hrs after3 days of adipogenic differentiation by RT-qPCR, respectively. All data are shown as Mean±SEM. *p<0.05, **p<0.01, ***p<0.001 |
When preadipocytes were treated with series of concentration gradient berberine for 48 hrs after 4 days of adipogenic differentiation, the expression level of miR-143 was decreased, but MAP2K5 mRNA expression was increased (Fig. 5c). Thus, their transcriptional actions are inversely correlated in general. Given all the above, it suggested that berberine has regulatory effects on adipogenic differentiation of preadipocytes by impacting the interaction of the miRNA and its target gene.
DISCUSSION
It is well known that reduction of animal saturated fats intake is an important step that can protect people from the risk factors of obesity and its related diseases. But pork is needed indeed nowadays, which has some fat inescapably. So decreasing porcine fat deposition is the public appeal of both consumers and producers.
Adipogenesis is indispensable in the process of fat deposition, the core of which is the proliferation and differentiation of preadipocytes. So it is always observed that numerous lipid vacuoles are accumulated in mature adipocytes. It is reported that berberine is utilized to attenuate adipogenic differentiation in various adipocytes as a traditional medicine from Chinese herbs, but its pinpoint mechanism still needs to be elucidated. Our experiment results showed that berberine could inhibit adipogenic differentiation visibly in a dose- and time-dependent manner. Triglycerides content in adipocytes was significantly decreased by berberine. These were consistent with previous findings on preadipocytes differentiation10-15. In addition, berberine impaired preadipocytes proliferation in a dose-dependent manner. This proved that berberine may have cytotoxicity, similar to responses produced in cancer cells31, suggesting that the resistance to adipogenesis of berberine might be concerned with the suppression of mitotic clonal expansion of preadipocytes, especially in fairly high concentration.
Adipogenesis is accompanied by the sequential regulation of a battery of adipogenic genes. PPARs are the key adipogenic genes of transcription factors, which could regulate the gene expression of adipocytes differentiation and gluconeogenesis32. It was reported that the lower expression of PPARγ could result in the inhibition of adipocytes differentiation in 3T3-L133. Berberine is an inhibitor of PPARγ during adipocytes differentiation12. In the current experiments, berberine inhibited the expression of PPARγ in a dose-dependent mannerin preadipocytes. So it was approved that berberine inhibits adipogenic differentiation through the PPARγ pathway. Adiponectin from adipocytes is abundantly present in serum, inversely correlated with fat deposits. It was reported that berberine inhibits the expression of adiponectin15. Similarly, it was found that berberine inhibits the expression of adiponectin in a dose-dependent manner in the current experiments. Based on the interaction of berberine and the important adipogenic genes, it is hypothesized that berberine plays a regulative role in relevant molecular signal pathways.
It is currently believed that miRNAs are involved in many biological processes and some can be secreted from fat cells or adipose tissue. So it should be expected to investigate the impact of berberine on the interaction of miRNAs and target genes in the process of adipogenic differentiation. As has been reported, miR-143 expedites adipogenic differentiation of preadipocytes in 3T3-L1, 293T or adipose-tissue-derived stromal cells and its performance is through targeting genes, such as ERK5, MAPK7, MAP2K5, pref-1/dlk-1 and pleiotrophin25,34-37. It was proved that miR-143 expression level was lower in the first 3 days, but significantly higher subsequently by RT-qPCR. The results and analysis of the Dual-Glo luciferase reporter assay confirmed that miR-143 acts upon preadipocytes differentiation by targeting MAP2K5 of the MAPK signaling pathway. The expression of MAP2K5 was suppressed at the mRNA and protein levels by miR-143 mimics since the sequence of miR-143 mimics is complementary to the seed region of 3 -UTR of MAP2K5. While the control of miR-143 mimics was abolished by specific mutations in binding sites of miR-143 and 3 -UTR of MAP2K5. It was changed that miR-143 exerts the effects on adipocytes differentiation by targeting MAP2K5 by berberine in a dose-dependent manner. There was a growing trend towards contrary on the relative mRNA expressions of miR-143 and MAP2K5 along with the increase of the consistency of berberine. It has been reported that the expression levels of miR-27a and miR-27b in 3T3-L1 cells treated with berberine were significantly increased and the berberine-mediated effect on adipogenic differentiation in 3T3-L1 was regulated with negative feedback by miR-27a and miR-27b38. Through suppression of the HNF-4α miR122 pathway, berberine reduced the development of the hepatic gluconeogenesis and lipid metabolism disorder in type 2 diabetic mice and palmitate-incubated HepG2 cells39.Unfortunately, there is insufficient evidence the effect of berberine on the interaction of miRNA and target genes at the protein level.
Now the majority of studies that berberine exerts regulatory effects on miRNAs have been focused on tumorigenesis, autophagy, glucose homeostasis and other conditions40. Effects of berberine on miRNAs in livestock adipogenesis have been scarcely illuminated. Our study provides more insight into the effects of berberine on livestock adipogenic differentiation.
CONCLUSION
This study investigated the effects of berberine on adipogenic differentiation in porcine intramuscular preadipocytes using different dominant methods. The results showed that berberine could inhibit adipogenic differentiation in porcine intramuscular preadipocytes in a time- and dose-dependent manner. By analysis, it is proved that berberine not only impairs the expressions of some key adipogenic genes, such as PPARγ, adiponectin but also interferes with the targeting of miR-143 to MAP2K5. Current study provides novel evidence for treatments of berberine to obtain healthy meat products against excessive fatty deposits, even human obesity, hyperlipidemia, diabetes, etc.
SIGNIFICANCE STATEMENT
The study described new approaches that berberine regulates adipogenic differentiation in porcine intramuscular preadipocytes through the multi-gene pattern. The information from these results could be provided to further investigate how berberine moderate’s lipid metabolism. While, these findings that berberine has a positive effect on lipid metabolism could be helpful to obtain healthy animal-based foods against excessive fatty deposits, even human obesity, hyperlipidemia, diabetes, etc.
ACKNOWLEDGMENTS
We are grateful to all kind editors and anonymous reviewers. The study was supported by Anhui Province Higher School Natural Science Key Program of China (KJ2018A0350), Scientific Research Project of Cooperation between Fuyang Municipal Government and Fuyang Normal University (XDHX201736) and Ph.D. Research Startup Fund of Fuyang Normal University (2016).
REFERENCES
- Andersen, C.J., K.E. Murphy and M.L. Fernandez, 2016. Impact of obesity and metabolic syndrome on immunity. Adv. Nutr., 7: 66-75.
CrossRefDirect Link - Park, H.S., U.I. Ju, J.W. Park, J.Y. Song and D.H. Shin et al., 2016. PPARγ neddylation essential for adipogenesis is a potential target for treating obesity. Cell Death Differ., 23: 1296-1311.
CrossRefDirect Link - Ahn, J., H. Wu and K. Lee, 2019. Integrative analysis revealing human adipose-specific genes and consolidating obesity loci. Sci. Rep., Vol. 9, No. 3087.
CrossRefDirect Link - Imenshahidi, M. and H. Hosseinzadeh, 2019. Berberine and barberry (Berberis vulgaris): A clinical review. Phytother. Res., 33: 504-523.
CrossRefDirect Link - Wang, K., X. Feng, L. Chai, S. Cao and F. Qiu, 2017. The metabolism of berberine and its contribution to the pharmacological effects. Drug Metab. Rev., 49: 139-157.
CrossRefDirect Link - Cicero, A.F.G. and A. Baggioni, 2016. Berberine and its role in chronic disease. Adv. Exp. Med. Biol., 928: 27-45.
CrossRefDirect Link - Pirillo, A. and A.L. Catapano, 2015. Berberine, a plant alkaloid with lipid- and glucose-lowering properties: From in vitro evidence to clinical studies. Atherosclerosis, 243: 449-461.
CrossRefDirect Link - Jin, Y., D.B. Khadka and W.J. Cho, 2015. Pharmacological effects of berberine and its derivatives: a patent update. Expert Opin. Ther. Pat., 26: 229-243.
CrossRefDirect Link - Hu, Y. and G.E. Davies, 2010. Berberine inhibits adipogenesis in high-fat diet-induced obesity mice. Fitoterapia, 81: 358-366.
CrossRefDirect Link - Pham, T.P.T., J. Kwon and J. Shin, 2011. Berberine exerts anti-adipogenic activity through up-regulation of C/EBP inhibitors, CHOP and DEC2. Biochem. Biophys. Res. Commun., 413: 376-382.
CrossRefDirect Link - Huang, C., Y. Zhang, Z. Gong, X. Sheng, Z. Li, W. Zhang and Y. Qin, 2006. Berberine inhibits 3T3-L1 adipocyte differentiation through the PPARγ pathway. Biochem. Biophys. Res. Commun., 348: 571-578.
CrossRefDirect Link - Zhang, J., H. Tang, R. Deng, N. Wang and Y. Zhang et al., 2015. Berberine suppresses adipocyte differentiation via decreasing creb transcriptional activity. PLOS ONE, Vol. 10.
CrossRefDirect Link - Choi, J.S., J.H. Kim, M.Y. Ali, B.S. Min, G.D. Kim and H.A. Jung, 2014. Coptis chinensis alkaloids exert anti-adipogenic activity on 3T3-L1 adipocytes by downregulating C/EBP-α and PPAR-γ. Fitoterapia, 98: 199-208.
CrossRefDirect Link - Li, Y., P. Wang, Y. Zhuang, H. Lin and Y. Li et al., 2011. Activation of AMPK by berberine promotes adiponectin multimerization in 3T3-L1 adipocytes. FEBS Lett., 585: 1735-1740.
CrossRefDirect Link - Chang, W., L. Chen and G.M. Hatch, 2016. Berberine treatment attenuates the palmitate-mediated inhibition of glucose uptake and consumption through increased 1,2,3-triacyl-sn-glycerol synthesis and accumulation in H9c2 cardiomyocytes. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids, 1861: 352-362.
CrossRefDirect Link - Lin, Y.C., Y.C. Lee, Y.J. Lin and J.C. Lin, 2019. Berberine promotes beige adipogenic signatures of 3T3-L1 cells by regulating post-transcriptional events. Cells, Vol. 8, No. 6.
CrossRefDirect Link - Bartel, D.P., 2004. MicroRNAs: Genomics, biogenesis, mechanism and function. Cell, 116: 281-297.
CrossRefDirect Link - Romao, J.M., W. Jin, M.V. Dodson, G.J. Hausman, S.S. Moore and L.L. Guan, 2011. MicroRNA regulation in mammalian adipogenesis. Exp. Biol. Med., 236: 997-1004.
CrossRefDirect Link - Chakraborty, C., C.G.P. Doss, S. Bandyopadhyay and G. Agoramoorthy, 2014. Influence of miRNA in insulin signaling pathway and insulin resistance: Micro-molecules with a major role in type-2 diabetes. Wiley Interdiscip. Rev.: RNA, 5: 697-712.
CrossRefDirect Link - Chang, W., 2017. Non-coding RNAs and berberine: A new mechanism of its anti-diabetic activities. Eur. J. Pharmacol., 795: 8-12.
CrossRefDirect Link - Esau, C., X. Kang, E. Peralta, E. Hanson and E.G. Marcusson et al., 2004. MicroRNA-143 regulates adipocyte differentiation. J. Biol. Chem., 279: 52361-52365.
CrossRefDirect Link - Ling, H.Y., G.B. Wen, S.D. Feng, Q.H. Tuo and H.S. Ou et al., 2011. MicroRNA-375 promotes 3T3-L1 adipocyte differentiation through modulation of extracellular signal-regulated kinase signalling. Clin. Exp. Pharmacol. Physiol., 38: 239-246.
CrossRefDirect Link - Jin, M., Y. Wu, J. Wang, J. Chen, Y. Huang, J. Rao and C. Feng, 2016. Microrna-24 promotes 3T3-L1 adipocyte differentiation by directly targeting the MAPK7 signaling. Biochem. Biophys. Res. Commun., 474: 76-82.
CrossRefDirect Link - Wang, J., Q. Ran, H.R. Zeng, L. Wang, C.J. Hu and Q.W. Huang, 2018. Cellular stress response mechanisms of rhizoma coptidis: A systematic review. Chin. Med., Vol. 13, No. 27.
CrossRefDirect Link - Zhang, Q., X. Xiao, K. Feng, T. Wang and W. Li et al., 2011. Berberine moderates glucose and lipid metabolism through multi pathway mechanism. Evid. Based Complement. Alternat. Med., Vol. 2011.
CrossRefDirect Link - Zhou, L., Y. Yang, X. Wang, S. Liu and W. Shang et al., 2007. Berberine stimulates glucose transport through a mechanism distinct from insulin. Metabolism, 56: 405-412.
CrossRefDirect Link - Lee, S., H.J. Lim, J.H. Park, K.S. Lee, Y. Jang and H.Y. Park, 2007. Berberine-induced LDLR up-regulation involves JNK pathway. Biochem. Biophys. Res. Commun., 362: 853-857.
CrossRefDirect Link - Yang, Y. and G. Yang, 2010. Rosiglitazone regulates IL-6-stimulated lipolysis in porcine adipocytes. Biochem. Cell Biol., 88: 853-860.
CrossRefDirect Link - Kim, J.B., J.H. Yu, E. Ko, K.W. Lee and A.K. Song et al., 2010. The alkaloid Berberine inhibits the growth of anoikis-resistant MCF-7 and MDA-MB-231 breast cancer cell lines by inducing cell cycle arrest. Phytomedicine, 17: 436-440.
CrossRef - Evans, R.M., G.D. Barish and Y.X. Wang, 2004. PPARs and the complex journey to obesity. Nat. Med., 10: 355-361.
CrossRefDirect Link - Farmer, S.R., 2006. Transcriptional control of adipocyte formation. Cell Metab., 4: 263-273.
CrossRefPubMedDirect Link - Chen, L., J. Hou, L. Ye, Y. Chen and J. Cui et al., 2014. MicroRNA-143 regulates adipogenesis by modulating the MAP2K5–ERK5 signaling. Sci. Rep., Vol. 4.
CrossRefDirect Link - Zhang, P., J. Du, L. Wang, L. Niu and Y. Zhao et al., 2018. MicroRNA-143a-3p modulates preadipocyte proliferation and differentiation by targeting MAPK7. Biomed. Pharmacother., 108: 531-539.
CrossRefDirect Link - Kim, Y.J., T.S. Min, K.S. Seo and S.H. Kim, 2015. Expression of pref-1/dlk-1 is regulated by microRNA-143 in 3T3-L1 cells. Mol. Biol. Rep., 42: 617-624.
CrossRefDirect Link - Yi, C., W.D. Xie, F. Li, Q. Lv and J. He et al., 2011. MiR-143 enhances adipogenic differentiation of 3T3-L1 cells through targeting the coding region of mouse pleiotrophin. FEBS Lett., 585: 3303-3309.
CrossRefDirect Link - Wu, Y.Y., X.M. Huang, J. Liu, Y. Cha and Z.P. Chen et al., 2016. Functional study of the upregulation of miRNA-27a and miRNA-27b in 3T3-L1 cells in response to berberine. Mol. Med. Rep., 14: 2725-2731.
CrossRefDirect Link - Wei, S., M. Zhang, Y. Yu, X. Lan and F. Yao et al., 2016. Berberine attenuates development of the hepatic gluconeogenesis and lipid metabolism disorder in type 2 diabetic mice and in palmitate-incubated Hepg2 cells through suppression of the HNF-4α miR122 pathway. PLOS ONE, Vol. 11.
CrossRefDirect Link - Ayati, S.H., B. Fazeli, A.A. Momtazi-Borojeni, A.F.G. Cicero, M. Pirro and A. Sahebkar, 2017. Regulatory effects of berberine on micrornome in cancer and other conditions. Crit. Rev. Oncol./Hematol., 116: 147-158.
CrossRefDirect Link