
M.M. Hongmei Chen
Affiliated Hospital of Inner Mongolia University for the Nationalities, 028000 Tongliao, Inner Mongolia Autonomous of China, China
M.D. Xiulan Wang
College of Traditional Mongolian Medicine and Pharmacy of Inner Mongolia University for the Nationalities, 028000 Tongliao, Inner Mongolia Autonomous of China, China
LiveDNA: 86.16492
M.M. Enhesuren
College of Traditional Mongolian Medicine and Pharmacy of Inner Mongolia University for the Nationalities, 028000 Tongliao, Inner Mongolia Autonomous of China, China
B.M. Chang Chun
College of Traditional Mongolian Medicine and Pharmacy of Inner Mongolia University for the Nationalities, 028000 Tongliao, Inner Mongolia Autonomous of China, China
B.M. Wenjie Jin
College of Traditional Mongolian Medicine and Pharmacy of Inner Mongolia University for the Nationalities, 028000 Tongliao, Inner Mongolia Autonomous of China, China
B.M. Mengqiqige
College of Traditional Mongolian Medicine and Pharmacy of Inner Mongolia University for the Nationalities, 028000 Tongliao, Inner Mongolia Autonomous of China, China
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 3 | Page No.: 310-319







ABSTRACT
Background and Objective: Nephropathy is a chronic non-communicable disease that can result in serious consequences. As the dried fruit of Zingiberaceae plant, Amomum cardamomum can improve renal function. This study aimed to reveal the action mechanisms of Amomum cardamomum in nephropathy rats. Materials and Methods: After nephropathy rat model was established, nephropathy rats were divided into model and drug groups (treated with Amomum cardamomum). The microRNA (miRNA) expression profilings of the rats were generated and the differentially expressed miRNAs (DE-miRNAs) between control and model groups were analyzed using the limma package. The DE-miRNAs between drug and model groups were also analyzed and then their targets were predicted by the miRWalk2.0 tool. Using DAVID tool, functional enrichment analysis for target genes was performed. In addition, protein-protein interaction (PPI) network and module analyses were conducted for target genes and miRNA-miRNA co-regulatory network was also constructed. Results: The nephropathy rat model was successfully established. A total of 21 DE-miRNAs were identified in drug group compared with model group. In the PPI networks, TP53, AKT2, HDAC1 and STAT3 had higher degrees. Besides, TP53 could interact with AKT2 and HDAC1. Moreover, STAT3 was co-regulated by rno-miR-30a-3p and rno-miR-30e-5p. Additionally, functional enrichment analysis showed that rno-miR-30a-3p-rno-miR-30e-5p, rno-miR-195-3p-rno-miR-32-3p separately had synergistic effects. Conclusion: Amomum cardamomum might improve the renal function of nephropathy rats by regulating TP53, AKT2, HDAC1, STAT3, miR-30a-5p, miR-30e-5p and miR-195-3p.
PDF Abstract XML References Citation
Received: May 21, 2017;
Accepted: November 13, 2017;
Published: March 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
M.M. Hongmei Chen, M.D. Xiulan Wang, M.M. Enhesuren, B.M. Chang Chun, B.M. Wenjie Jin and B.M. Mengqiqige, 2018. MicroRNA Microarray Analysis to Investigate the Key Genes and microRNAs Regulated by Amomum cardamomum in Nephropathy Rat Model. International Journal of Pharmacology, 14: 310-319.
DOI: 10.3923/ijp.2018.310.319
URL: https://scialert.net/abstract/?doi=ijp.2018.310.319
DOI: 10.3923/ijp.2018.310.319
URL: https://scialert.net/abstract/?doi=ijp.2018.310.319
INTRODUCTION
Nephropathy (also named kidney disease or renal disease) is damage to or disease of a kidney. Nephropathy is usually caused by administration of analgesics, deposition of the IgA antibodies in the glomerulus, toxicity of chemotherapy agents and xanthine oxidase deficiency1. Nephropathy without effective controlling can result in serious consequences, such as renal insufficiency, renal failure and uremia2. Millions of people are undergoing nephropathy globally, among which thousands of people need kidney transplants3. Therefore, revealing the key mechanisms underlying nephropathy is of great significance for developing effective therapeutic strategies.
The pathogenesis of nephropathy has been explored by several researches. For example, kidney-targeting Smad7 gene transfer may improve type 2 diabetic nephropathy through inhibiting the nuclear factor κB (NF-κB) and transforming growth factor-β (TGF-β)/SMAD signaling pathways4. The increased matrix metalloproteinase-9 (MMP-9) is adverse in renal interstitial fibrogenesis, which can result in the destruction of tubular basement membrane and the promotion of epithelial to myofibroblast transition5. MicroRNA-21 (miR-21) is reported to play an important role in the pathology of fibrosis and is suggested as a target for the treatment of diabetic nephropathy6,7. The miR-135a functions in renal fibrosis via regulating transient receptor potential cation channel, subfamily C, member 1 (TRPC1) and inhibition of miR-135a is considered effective for treating diabetic nephropathy8. The expression of miR-29c in urinary exosome has impacts on both the degree of histological fibrosis and renal function, suggesting that miR-29c may serve as a promising marker for renal fibrosis9,10.
Cardamom (Amomum cardamomum) is the dried fruit of Amomum cardamomum, which is a perennial herb belonging to the Ginger family and having strong antioxidant activity11. Amomum cardamomum has been found to improve renal function of alloxan induced diabetic rats and induce the decreased uric acid, creatinine, urea and the enhanced serum protein activity12. However, the action mechanisms of Amomum cardamomum in nephropathy have not been fully revealed yet.
In this study, Wistar rats were injected with doxorubicin hydrochloride to establish nephropathy rat model. Then, Amomum cardamomum was used to treat the rats with nephropathy. The miRNA expression profiling of the rats was then generated, followed by identification of the differentially expressed miRNAs (DE-miRNAs) and prediction of their targets. Subsequently, functional enrichment analysis, protein-protein interaction (PPI) network and module analyses for target genes was performed. Besides, miRNA-miRNA co-regulatory network was also constructed. The objective of this study was to identify the key miRNAs and genes associated with the effects of Amomum cardamomum on nephropathy. The findings of this study will help to discover new drug targets for nephropathy, thus improving the clinical outcome of this disease.
MATERIALS AND METHODS
Sample treatment: Male Wistar rats weighting 200-220 g were obtained from Yisi Laboratory Animal Technology Co., Ltd. (Jilin, Changchun, China). After the rats were fed for 5 days, their 24 h urine was collected and then urine protein quantity was determined. A total of 15 rats with normal urine protein quantity were chosen, from which 5 rats were randomly included into normal control group and were injected with normal saline (6.5 mL kg–1) through caudal vein. The remaining 10 rats were injected with doxorubicin hydrochloride (6.5 mg kg–1, Shenzhen Arcandor’s Pharmaceutical Co., LTD, Guangdong, Shenzhen, China) into caudal vein to establish nephropathy rat model. From the 4th day after establishment of nephropathy rat model, 5 rats were randomly included into drug group which were treated with Amomum cardamomum (3.0 g kg–1, once a day, continuing for 21 days, Tong Kang Pharmaceutical Co., LTD, Hebei, Anguo, China) by intragastric administration, while the other 5 rats were given by gavage with the same volume of distilled water as model group. Meanwhile, the rats in control group were also given by gavage with the same volume of distilled water.
Microarray detection and DE-miRNAs screening: The miRNA expression profilings of the rats were generated by miRNA 4.0 Array (Affymetrix, Santa Clara, CA). Then, the raw data were preprocessed using the Robust Multichip Average (RMA) method of Affy package13 in R, including background correction and normalization. To confirm whether nephropathy rat model was successfully established or not, the DE-miRNAs between control and model groups were identified using the t-test method14 in limma package15. In addition, the DE-miRNAs between drug group and model group were also analyzed using the same method. The p-value<0.05 and |log2 fold change (FC)|>0.58 were taken as the thresholds.
Prediction of the targets of DE-miRNAs: Based on the information of miRWalk16, MicroT417, miRanda18, miRBridge19, miRDB20, miRMap21, miRNAMap22, PICTAR223, PITA24, RNA2225, RNAhybrid26 and TargetScan27 databases, the genes targeted by the DE-miRNAs between drug group and model group were predicted using the miRWalk2.0 tool28. The miRNA-gene pairs predicted by no less than 6 databases were selected.
Functional and pathway enrichment analysis: Gene ontology (GO) database introduces gene products from molecular function (MF), biological process (BP) and cellular component (CC) aspects29. The Kyoto Encyclopedia of Genes and Genomes (KEGG) database is developed for predicting the pathways involving genes or other molecules30. Using the Database for Annotation, Visualization and Integrated Discovery (DAVID) online tool31, GO functional and KEGG pathway enrichment analyses for target genes of the DE-miRNAs were performed, with p-value<0.05 and gene count>2 as the thresholds.
PPI network and module analyses: The PPI pairs among the target genes were predicted by the Search Tool for the Retrieval of Interacting Genes (STRING, version10,)32 database and then PPI network was visualized by Cytoscape software33. Using the CytoNCA plug-in (parameter: Without weight)34 in cytoscape software, closeness centrality (CC), degree centrality (DC) and betweenness centrality (BC) of the nodes in the PPI network were calculated to identify the hub proteins35. Using the MCODE plug-in (parameters: Degree cutoff = 2, node score cutoff = 0.2, Max. depth = 100, K-core = 2)36 in cytoscape software, module analysis was carried out for the PPI network.
Analysis of co-regulated target genes: Based on the target genes co-regulated by two miRNAs, miRNA-miRNA co-regulatory network was constructed by Cytoscape software33. For miRNA-miRNA pairs co-regulated more than 10 target genes, their targets were performed with GO functional enrichment analysis, with p<0.05 as the threshold. If the target genes co-regulated by two miRNAs were enriched in no less than one GO–BP term, the two miRNAs were thought to have synergistic effects in functional level.
RESULTS
Model validation and DE-miRNAs screening: Under the thresholds of p<0.05 and |log2 FC|>0.58, a total of 3645 DE-miRNAs in model group compared with control group were screened, including 1324 up and 2321 down-regulated miRNAs, indicating that nephropathy rat model was successfully established. The heat map of clustering analysis was shown in Fig. 1. In addition, a total of 21 DE-miRNAs were identified in drug group compared with model group, including 6 up- and 15 down-regulated miRNAs (Fig. 2). For the up-regulated and down-regulated miRNAs, 906 and 1653 miRNA→target gene pairs were predicted, respectively. The number of genes targeted by each DE-miRNA was listed in Table 1.
| Table 1: | Number of genes targeted by each differentially expressed miRNA |
 | |
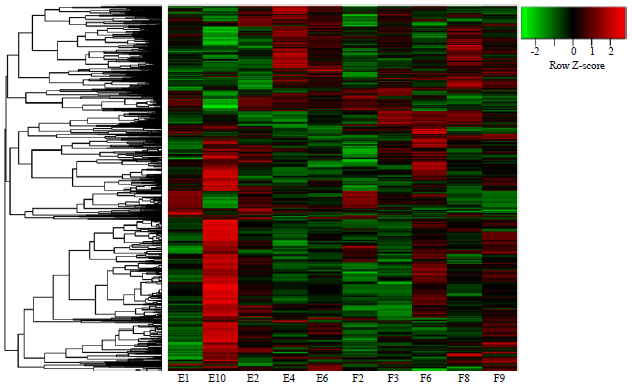 | |
| Fig. 1: | The heat map of clustering analysis for the differentially expressed miRNAs (DE-miRNAs) between model group and control group |
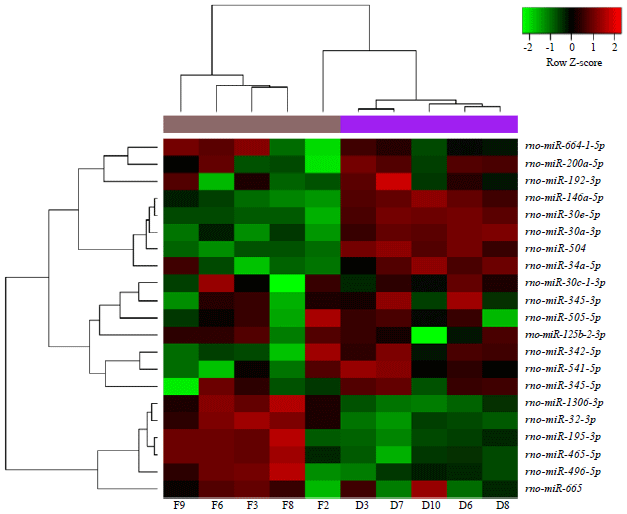 | |
| Fig. 2: | The heat map of clustering analysis for the differentially expressed miRNAs (DE-miRNAs) between drug group and model group |
Functional and pathway enrichment analysis: The GO functional and pathway enrichment analyses for target genes were performed. The target genes of up-regulated miRNAs were mainly enriched in GO–BP function associated with neuron differentiation and cell cycle pathway (KEGG) (Fig. 3a). Meanwhile, the target genes of down-regulated miRNAs were enriched in GO–BP function related to regulation of transcription and pathways in cancer (KEGG) (Fig. 3b).
PPI network and module analyses: The PPI networks for the target genes of up-regulated miRNAs and down-regulated miRNAs were constructed separately. The nodes with higher degrees in the PPI networks were listed in Table 2, including tumor protein p53 (TP53, degree = 97), v-akt murine thymoma viral oncogene homolog 2 (AKT2, degree = 51) and histone deacetylase 1 (HDAC1, degree = 48) targeted by up-regulated miRNAs, as well assignal transducer and activator of transcription 3 (STAT3, degree = 71) targeted by down-regulated miRNAs. Importantly, TP53 had interactions with both AKT2 and HDAC1. The most significant module (score = 10) identified from the PPI network for the target genes of up-regulated miRNAs was shown in Fig. 4a, which had 10 nodes and 45 edges.
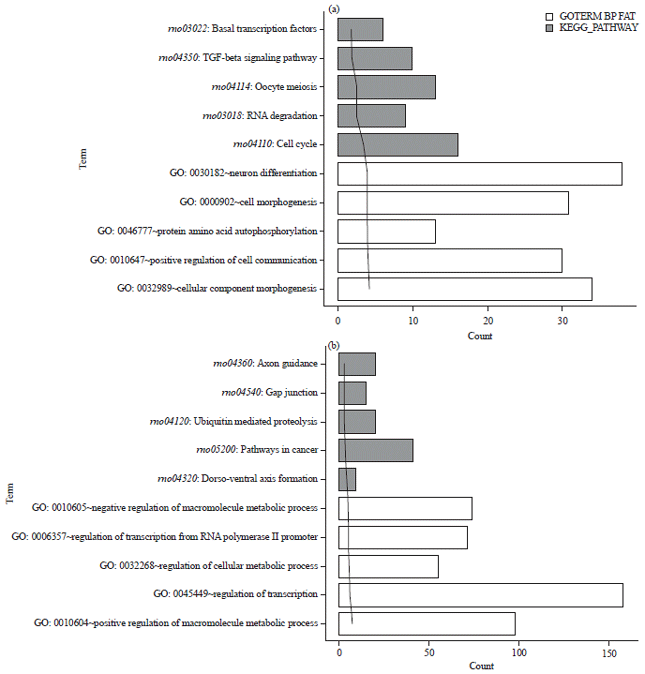 | |
| Fig. 3(a-b): | The top 5 terms enriched for the target genes of (a) Up-regulated miRNAs and (b) Down-regulated miRNAs |
GO: Gene ontology, BP: Biological process, KEGG: Kyoto Encyclopedia of Genes and Genomes | |
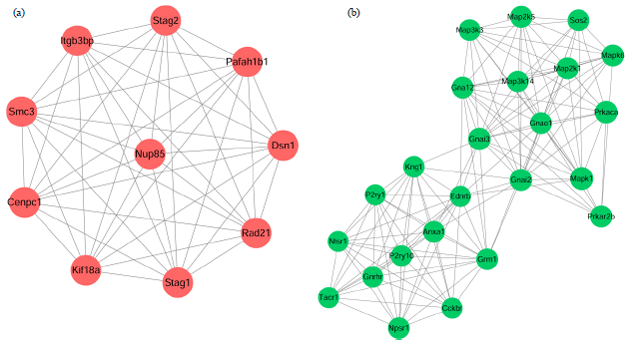 | |
| Fig. 4(a-b): | The most significant modules identified from the protein-protein interaction (PPI) network for the target genes of (a) Up-regulated miRNAs and (b) Down-regulated miRNAs |
| Table 2: | The nodes with higher degrees in the protein-protein interaction (PPI) networks for the target genes of up-regulated miRNAs and down-regulated miRNAs |
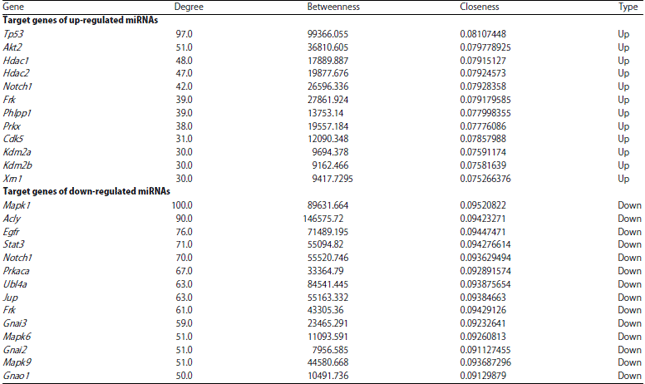 | |
Moreover, the most significant module (score = 11.739) of the PPI network constructed for the target genes of down-regulated miRNAs had 24 nodes and 135 edges (Fig. 4b).
Analysis of co-regulated target genes: The miRNA-miRNA co-regulatory network was shown in Fig. 5 and the miRNA-miRNA pairs which co-regulated more than 10 target genes were listed in Table 3.
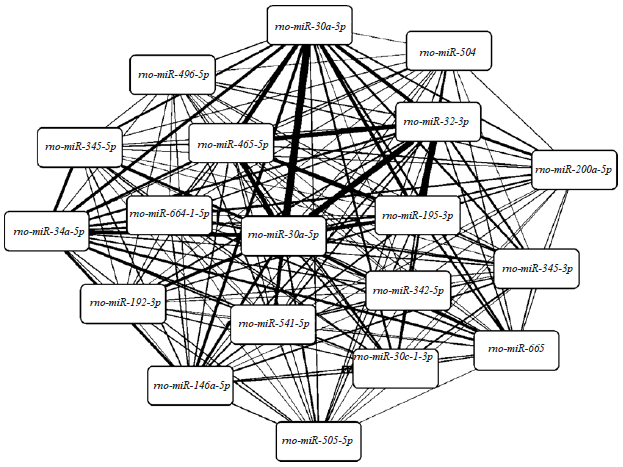 | |
| Fig. 5: | The miRNA-miRNA co-regulatory network. The thickness of edges represents the number of co-regulated genes |
| Table 3: | The miRNA-miRNA pairs co-regulated more than 10 target genes |
 | |
Especially, STAT3 was co-regulated by rno-miR-30a-3p and rno-miR-30e-5p. Functional enrichment analysis was performed for the target genes of the miRNA-miRNA pairs which co-regulated more than 10 targets and the number of enriched terms for each miRNA-miRNA pair was listed in Table 4. Notably, rno-miR-30a-3p-rno-miR-30e-5p, rno-miR-195-3p-rno-miR-32-3p, rno-miR-30e-5p-rno-miR-32-3p, rno-miR-30a-3p-rno-miR-32-3p and rno-miR-30e-5p-rno-miR-541-5p were miRNA-miRNA pairs that had synergistic effects.
DISCUSSION
In this study, a total of 21 DE-miRNAs (including 6 up and 15 down-regulated miRNAs) were identified in drug group compared with model group. Afterwards, 906 and 1653 miRNA→target gene pairs were separately predicted for the up-regulated and down-regulated miRNAs. In the PPI networks constructed for the target genes of DE-miRNAs, TP53, AKT2, HDAC1 and STAT3 had higher degrees.
| Table 4: | The number of enriched terms for each miRNA-miRNA pair. GO, Gene Ontology; BP, biological process |
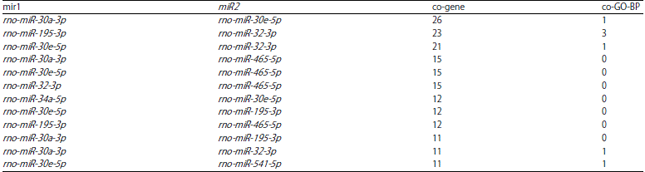 | |
HDAC1 and HDAC2 play critical roles in mediating the proliferation of renal interstitial fibroblasts, activation of STAT3 and expression of cell cycle proteins, additionally, STAT3 regulates the proliferative effects of HDACs37. Trichostatin A (TSA, an HDAC inhibitor) and other HDAC inhibitors are reported to be novel therapeutic agents for tubular epithelial-mesenchymal transition (EMT) in renal epithelial cells38. Ursolic acid (UA) has anti-oxidant activity and regulates the NF-κB and STAT3 signaling pathways, thus it can suppress carbon tetrachloride-induced inflammation in mouse kidney39. HIV-1 Nef causes the proliferation and dedifferentiation of podocytes through activating the mitogen-activated protein kinase 1, 2 (MAPK1,2) and STAT3 pathways and STAT3 functions in the progression of human immunodeficiency virus (HIV)-associated nephropathy40. These findings suggest that HDAC1 and STAT3 may be involved in the progression of nephropathy.
Through the p53 activation and STAT3 dephosphorylation, aristolochic acid (AA) leads to the death of tubular epithelial cells, suggesting that p53 contributes to renal injury in acute AA nephropathy41. The expression levels of p53, TGF-β and miR-192 are significantly up-regulated in the rats with diabetic nephropathy, which may accelerate the progression of diabetic nephropathy42. AKT2 acts in preserving podocyte viability and function, indicating that AKT2 may be used for maintaining glomerular function in chronic kidney disease43. Knockdown of AKT2 antagonizes TGF-β1-induced EMT via suppressing glycogen synthase kinase-3β (GSK3β)/Snail signaling pathway in renal tubular epithelial cells44. What’s more, AKT2/protein kinase B β (PKBβ ) is reported to function in the regulation of renal phosphate and glucose transports45,46. Therefore, TP53 and AKT2 may play roles in the pathogenesis of nephropathy. In the PPI network for the target genes of up-regulated miRNAs, TP53 had interactions with both AKT2 and HDAC1, indicating that TP53 might also function in nephropathy through interacting with AKT2 and HDAC1.
Furthermore, a previous study reveals that the levels of miR-30a-5p, miR-196a and miR-490 have correlations with the activity of focal segmental glomerulosclerosis disease47. Inhibition of miR-30a-3p and miR-30c-2-3p promotes hypoxia-inducible factor 2α (HIF2α) expression, which reduces the tumor suppressive effects of HIF1α in von-Hippel Lindau (VHL)-deficient clear cell renal cell carcinomas (ccRCC)48. The miR-30e/uncoupling protein 2 (UCP2) axis is critical for regulating EMT and kidney fibrosis induced by TGF-β1 and may be a promising therapeutic approach for fibrotic kidney disease49. The decreased expression of miR-195 protects mesangial cells against apoptosis, thus the anti-apoptosis effect of miR-195 may be important for the early stages of diabetic nephropathy50. Thus, miR-30a-5p, miR-30e-5p and miR-195-3p may also affect nephropathy. In this study, functional enrichment analysis showed that rno-miR-30a-3p and rno-miR-30e-5p had synergistic effect. Besides, STAT3 was co-regulated by miR-30a-3p and miR-30e-5p. Thus, we speculate that miR-30a-3p and miR-30e-5p may have a synergistic effect in nephropathy by regulating STAT3.
CONCLUSION
It is concluded that Amomum cardamomum may improve the renal function of nephropathy rats by regulating several key genes (including TP53, AKT2, HDAC1, STAT3, miR-30a-5p, miR-30e-5p and miR-195-3p). However, further experimental researches are still necessary for confirming these findings.
SIGNIFICANCE STATEMENT
This study discovers the synergistic effects of key genes and miRNAs regulated by Amomum cardamomum that can be beneficial for improving the renal function of nephropathy rats. This study will help the researcher to uncover the action mechanisms of Amomum cardamomum in nephropathy that many researchers are not able to explore. Thus, a new theory on these gene-miRNA interactions may be arrived at.
ACKNOWLEDGMENTS
This study was supported by Natural Science Foundation of Inner Mongolia Autonomous Region (No. 2012ZD12).
REFERENCES
- Hajivandi, A. and M. Amiri, 2014. World kidney day 2014: Kidney disease and elderly. J. Parathyroid Dis., 2: 3-4.
Direct Link - Lemley, K.V., 2016. Glomerular pathology and the progression of chronic kidney disease. Am. J. Physiol.-Renal Physiol., 310: F1385-F1388.
CrossRefDirect Link - Jha, V., G. Garcia-Garcia, K. Iseki, Z. Li and S. Naicker et al., 2013. Chronic kidney disease: Global dimension and perspectives. Lancet, 382: 260-272.
CrossRefDirect Link - Ka, S.M., Y.C. Yeh, X.R. Huang, T.K. Chao and Y.J. Hung et al., 2012. Kidney-targeting Smad7 gene transfer inhibits renal TGF-β/MAD homologue (SMAD) and Nuclear Factor κB (NF-κB) signalling pathways and improves diabetic nephropathy in mice. Diabetologia, 55: 509-519.
CrossRefPubMedDirect Link - Wang, X., Y. Zhou, R. Tan, M. Xiong and W. He et al., 2010. Mice lacking the matrix metalloproteinase-9 gene reduce renal interstitial fibrosis in obstructive nephropathy. Am. J. Physiol.-Renal Physiol., 299: F973-F982.
CrossRefDirect Link - Zhong, X., A.C.K. Chung, H.Y. Chen, Y. Dong and X.M. Meng et al., 2013. miR-21 is a key therapeutic target for renal injury in a mouse model of type 2 diabetes. Diabetologia, 56: 663-674.
CrossRefDirect Link - Wang, J., Y. Gao, M. Ma, M. Li and D. Zou et al., 2013. Effect of miR-21 on renal fibrosis by regulating MMP-9 and TIMP1 in kk-ay diabetic nephropathy mice. Cell Biochem. Biophys., 67: 537-546.
CrossRefDirect Link - He, F., F. Peng, X. Xia, C. Zhao and Q. Luo et al., 2014. MiR-135a promotes renal fibrosis in diabetic nephropathy by regulating TRPC1. Diabetologia, 57: 1726-1736.
CrossRefDirect Link - Lv, L.L., Y.H. Cao, H.F. Ni, M. Xu and D. Liu et al., 2013. MicroRNA-29c in urinary exosome/microvesicle as a biomarker of renal fibrosis. Am. J. Physiol.-Renal Physiol., 305: F1220-F1227.
CrossRefDirect Link - Fang, Y., X. Yu, Y. Liu, A.J. Kriegel and Y. Heng et al., 2013. miR-29c is downregulated in renal interstitial fibrosis in humans and rats and restored by HIF-α activation. Am. J. Physiol.-Renal Physiol., 304: F1274-F1282.
CrossRefDirect Link - Verma, D., R. Yadav and M. Kumar, 2015. Effect of seed extract of Amomum cardamomum on renal function in alloxan induced diabetic rats. World J. Clin. Pharmacol. Microbiol. Toxicol., 1: 52-54.
Direct Link - Gautier, L., L. Cope, B.M. Bolstad and R.A. Irizarry, 2004. Affy-analysis of Affymetrix GeneChip data at the probe level. Bioinformatics, 20: 307-315.
CrossRefDirect Link - Ritchie, M.E., B. Phipson, D. Wu, Y. Hu, C.W. Law, W. Shi and G.K. Smyth, 2015. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res., Vol. 43.
CrossRefDirect Link - Dweep, H., N. Gretz and C. Sticht, 2014. miRWalk database for miRNA-target interactions. Methods Mol. Biol., 1182: 289-305.
PubMed - Maragkakis, M., T. Vergoulis, P. Alexiou, M. Reczko and K. Plomaritou et al., 2011. DIANA-microT Web server upgrade supports Fly and Worm miRNA target prediction and bibliographic miRNA to disease association. Nucleic Acids Res., 39: W145-W148.
CrossRefDirect Link - Enright, A.J., B. John, U. Gaul, T. Tuschl, C. Sander and D.S. Marks, 2003. MicroRNA targets in Drosophila. Genome Biol., Vol. 5.
CrossRefDirect Link - Tsang, J.S., M.S. Ebert and A. van Oudenaarden, 2010. Genome-wide dissection of MicroRNA functions and cotargeting networks using gene set signatures. Mol. Cell, 38: 140-153.
CrossRefDirect Link - Wong, N. and X. Wang, 2014. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res., 43: D146-D152.
CrossRefDirect Link - Vejnar, C.E. and E.M. Zdobnov, 2012. MiRmap: Comprehensive prediction of microRNA target repression strength. Nucleic Acids Res., 40: 11673-11683.
CrossRefDirect Link - Hsu, S.D., C.H. Chu, A.P. Tsou, S.J. Chen and H.C. Chen et al., 2007. miRNAMap 2.0: Genomic maps of microRNAs in metazoan genomes. Nucleic Acids Res., 36: D165-D169.
CrossRefDirect Link - Blin, K., C. Dieterich, R. Wurmus, N. Rajewsky, M. Landthaler and A. Akalin, 2014. DoRiNA 2.0-upgrading the doRiNA database of RNA interactions in post-transcriptional regulation. Nucleic Acids Res., 43: D160-D167.
CrossRefDirect Link - Kertesz, M., N. Iovino, U. Unnerstall, U. Gaul and E. Segal, 2007. The role of site accessibility in microRNA target recognition. Nat. Genet., 39: 1278-1284.
CrossRefDirect Link - Miranda, K.C., T. Huynh, Y. Tay, Y.S. Ang and W.L.Tam et al., 2006. A pattern-based method for the identification of microrna binding sites and their corresponding heteroduplexes. Cell, 126: 1203-1217.
CrossRefDirect Link - Kruger, J. and M. Rehmsmeier, 2006. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res., 34: W451-W454.
CrossRefDirect Link - Shin, C., J.W. Nam, K.K.H. Farh, H.R. Chiang, A. Shkumatava and D.P. Bartel, 2010. Expanding the microRNA targeting code: Functional sites with centered pairing. Mol. Cell, 38: 789-802.
CrossRefDirect Link - Dweep, H. and N. Gretz, 2015. miRWalk2.0: A comprehensive atlas of microRNA-target interactions. Nat. Methods, 12: 697-697.
CrossRefDirect Link - Tweedie, S., M. Ashburner, K. Falls, P. Leyland and P. McQuilton et al., 2008. FlyBase: Enhancing Drosophila gene ontology annotations. Nucleic Acids Res., 37: D555-D559.
CrossRefDirect Link - Kanehisa, M. and S. Goto, 2000. KEGG: Kyoto encyclopedia of genes and genomes. Nucl. Acids Res., 28: 27-30.
CrossRefDirect Link - Dennis, Jr., G., B.T. Sherman, D.A. Hosack, J. Yang, W. Gao, H.C. Lane and R.A. Lempicki, 2003. DAVID: Database for annotation, visualization and integrated discovery. Genome Biol., Vol. 4.
CrossRefDirect Link - Szklarczyk, D., A. Franceschini, S. Wyder, K. Forslund and D. Heller et al., 2015. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res., 43: D447-D452.
CrossRefDirect Link - Saito, R., M.E. Smoot, K. Ono, J. Ruscheinski and P.L. Wang et al., 2012. A travel guide to Cytoscape plugins. Nat. Methods, 9: 1069-1076.
CrossRefDirect Link - Tang, Y., M. Li, J. Wang, Y. Pan and F.X. Wu, 2015. CytoNCA: A cytoscape plugin for centrality analysis and evaluation of protein interaction networks. Biosystems, 127: 67-72.
CrossRefDirect Link - He, X. and J. Zhang, 2006. Why do hubs tend to be essential in protein networks? PLoS Genet., Vol. 2.
CrossRefDirect Link - Bader, G.D. and C.W. Hogue, 2003. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform., Vol. 4.
CrossRefDirect Link - Pang, M., L. Ma, N. Liu, M. Ponnusamy, T.C. Zhao, H. Yan and S. Zhuang, 2011. Histone deacetylase 1/2 mediates proliferation of renal interstitial fibroblasts and expression of cell cycle proteins. J. Cell. Biochem., 112: 2138-2148.
CrossRefPubMedDirect Link - Yoshikawa, M., K. Hishikawa, T. Marumo and T. Fujita, 2007. Inhibition of histone deacetylase activity suppresses epithelial-to-mesenchymal transition induced by TGF-β1 in human renal epithelial cells. J. Am. Soc. Nephrol., 18: 58-65.
CrossRefDirect Link - Ma, J.Q., J. Ding, Z.H. Xiao and C.M. Liu, 2014. Ursolic acid ameliorates carbon tetrachloride-induced oxidative DNA damage and inflammation in mouse kidney by inhibiting the STAT3 and NF-κB activities. Int. Immunopharmacol., 21: 389-395.
CrossRefDirect Link - Feng, X., T.C. Lu, P.Y. Chuang, W. Fang and K. Ratnam et al., 2009. Reduction of STAT3 activity attenuates HIV-induced kidney injury. J. Am. Soc. Nephrol., 20: 2138-2146.
CrossRefDirect Link - Zhou, L., P. Fu, X.R. Huang, F. Liu, K.N. Lai and H.Y. Lan, 2010. Activation of p53 promotes renal injury in acute aristolochic acid nephropathy. J. Am. Soc. Nephrol., 21: 31-41.
CrossRefDirect Link - Deshpande, S.D., S. Putta, M. Wang, J.Y. Lai and M. Bitzer et al., 2013. Transforming growth factor-β-induced cross talk between p53 and a microRNA in the pathogenesis of diabetic nephropathy. Diabetes, 62: 3151-3162.
CrossRefDirect Link - Canaud, G., F. Bienaime, A. Viau, C. Treins and W. Baron et al., 2013. AKT2 is essential to maintain podocyte viability and function during chronic kidney disease. Nat. Med., 19: 1288-1296.
CrossRefDirect Link - Lan, A., Y. Qi and J. Du, 2014. Akt2 mediates TGF-β1-induced epithelial to mesenchymal transition by deactivating GSK3β/snail signaling pathway in renal tubular epithelial cells. Cell. Physiol. Biochem., 34: 368-382.
CrossRefDirect Link - Kempe, D.S., G. Siraskar, H. Frohlich, A.T. Umbach and M. Stubs et al., 2010. Regulation of renal tubular glucose reabsorption by Akt2/PKBβ. Am. J. Physiol.-Renal Physiol., 298: F1113-F1117.
CrossRefDirect Link - Kempe, D.S., T.F. Ackermann, K.M. Boini, F. Klaus and A.T. Umbach et al., 2010. Akt2/PKBβ-sensitive regulation of renal phosphate transport. Acta Physiol., 200: 75-85.
CrossRefDirect Link - Zhang, W., C. Zhang, H. Chen, L. Li and Y. Tu et al., 2014. Evaluation of microRNAs miR-196a, miR-30a-5P and miR-490 as biomarkers of disease activity among patients with FSGS. Clin. J. Am. Soc. Nephrol., 9: 1545-1552.
PubMedDirect Link - Mathew, L.K., S.S. Lee, N. Skuli, S. Rao and B. Keith et al., 2013. Restricted expression of miR-30c-2-3p and miR-30a-3p in clear cell renal carcinomas enhances HIF2α activity. Cancer Discov., 4: 53-60.
CrossRefPubMedDirect Link - Jiang, L., W. Qiu, Y. Zhou, P. Wen and L. Fang et al., 2013. A microRNA-30e/mitochondrial uncoupling protein 2 axis mediates TGF-β1-induced tubular epithelial cell extracellular matrix production and kidney fibrosis. Kidney Int., 84: 285-296.
CrossRefDirect Link - Chen, Y.Q., X.X. Wang, X.M. Yao, D.L. Zhang, X.F. Yang, S.F. Tian and N.S. Wang, 2012. Abated microRNA-195 expression protected mesangial cells from apoptosis in early diabetic renal injury in mice. J. Nephrol., 25: 566-576.
PubMed