
Manouchehr Safari
Research Center of Nervous System Stem Cells, Department of Anatomy, Semnan University of Medical Sciences, Semnan, Iran
Laya Ghahari
Department of Anatomy, AJA University of Medical Sciences, Tehran, Iran
Katayoun Safari
Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran
Mahmoodreza Madadian
School of Pharmacy, Pharmaceutical Sciences Branch, Islamic Azad University,Iran
Mohammad Reza Aldaghi
Research Center of Nervous System Stem Cells, Department of Anatomy, Semnan University of Medical Sciences, Semnan, Iran
Sam Zarbakhsh
Research Center of Nervous System Stem Cells, Department of Anatomy, Semnan University of Medical Sciences, Semnan, Iran
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 4 | Page No.: 359-365







ABSTRACT
Regenerative medicine using bone marrow cells is noteworthy therapy for the improvement of patients with severe liver disease. Bone marrow stem cells can participate to liver regeneration. However, conflicting results have been reported. Twenty five, 12 week old albino-wistar male rats were randomly assigned into five groups. In treatment groups CD133+ cells in both differentiated and non-differentiated form were transplanted in rat models of CCl4-induced chronic liver dysfunction. All rats were killed 4 weeks after transplantation. Transplantation of the differentiated and non differentiated hematopoietic stem cells into the blood of rats significantly restored their serum albumin level. These effects were not seen in CCl4 rats. However, fibrotic liver lesions with severe inflammation were decreased in CCl4-treated following hematopoietic stem cells transplantation. Effective migration of transplanted hematopoietic stem cells was limited to persistently injured liver in CCl4-treated rat, where they may be effective in resolving inflammatory fibrotic lesions. These results suggest that differentiated Hematopoietic stem cells are an effective cell source for liver regeneration.
PDF Abstract XML References Citation
Received: January 05, 2015;
Accepted: February 26, 2015;
Published: April 18, 2015
How to cite this article
Manouchehr Safari, Laya Ghahari, Katayoun Safari, Mahmoodreza Madadian, Mohammad Reza Aldaghi and Sam Zarbakhsh, 2015. Therapeutic Effects of CD133+ in the Carbon Tetrachloride (CCl4) Induced Chronic Liver Dysfunction in Rat Model. International Journal of Pharmacology, 11: 359-365.
DOI: 10.3923/ijp.2015.359.365
URL: https://scialert.net/abstract/?doi=ijp.2015.359.365
DOI: 10.3923/ijp.2015.359.365
URL: https://scialert.net/abstract/?doi=ijp.2015.359.365
INTRODUCTION
Cirrhosis demonstrates the definitive common pathological outcome for the majority of chronic liver diseases. Most patients with cirrhosis die from one or more of clinical complications such as ascites, hepatic encephalopathy and hemorrhage (Bataller et al., 2005). The majority of chronic liver diseases are initiated by the infection of Hepatitis C Virus (HCV) and Hepatitis B Virus (HBV) (Lai et al., 2003; Poynard et al., 2003). Chronic hepatitis can develop into hepatic cirrhosis (Poldervaart et al., 2009).
As limited treatments for liver disease, liver transplantation is the most effective therapy for the patients with advanced liver diseases. However, transplantation has specific problems, such as a lack of donors, operative damage, post-transplant rejection, high costs and a requirement for immunosuppressive therapy. Over 10% of patients die while waiting for liver transplantation. Among the patients who received liver transplants, the survival rate has been 94% at 3 months, 88% at 1 year and 79% at 3 years (Friedman, 2008). Thus, it is of great interest to search for an effective alternate to treat this type of life-threatening disease. Recently, regenerative medicine using stem cells-based cytotherapy has shown as a benefit therapy with minimally invasive procedures and with few complications for treating patients with severe liver disease. Stem cells are defined as undifferentiated cells from an embryo, fetus or an adult which have the potential to generate, in vitro and in vivo, various differentiated tissue cells (Caplan, 1991; Dominici et al., 2006; Pittenger et al., 1999). Several studies have shown that Bone Marrow (BM) derived Hematopoietic Stem Cells (HSCs) such as CD133+ have a high degree of plasticity are an attractive cell source for liver regeneration, due to the ease of obtaining bone marrow stem cells compared to other tissue-specific stem cells. Many reports have shown the capacity of bone marrow stromal cells to differentiate into hepatocytes (Alison et al., 2000; Korbling et al., 2002; Okamoto et al., 2002; Theise et al., 2000).
The number of circulating CD133+ HSCs expressing early markers for hepatic differentiation increase subsequent treatment with mobilizing factors such as Granulocyte-Colony Stimulating Factor (G-CSF). This effect has led to speculation about the existence of BM-derived hepatic-committed Stem Cells (SCs) which could migrate from the peripheral blood into the liver and contribute to its regeneration and repair (Piscaglia et al., 2003a, b; Ratajczak et al., 2004).
To investigate the efficacy of HSCs treatment in improving liver conditions CCl4-induced chronic liver dysfunction rat were employed.
MATERIALS AND METHODS
Animals and experimental protocol: Adult male albino-wistar rat weighing 200-250 g were obtained from the animal center of Iran University in Tehran, Iran. All rats were housed with food and water available ad libitum. The storage room was maintained at constant temperature and humidity on a 12 h dark and light. All procedures were carried out in accordance with the National Institutes of Health Guide for Care and Use of Laboratory Animals that authorized by Ethics Committee of Semnan Medical University Semnan, Iran. Rats were randomly divided into five groups (n = 5). Each group were housed in a separate cages, the first group received olive oil (vehicle of CCl4) as a control. The second group received 1 cc kg-1 (CCl4) repeatedly administered into the peritoneal cavity, twice weekly, for 2 weeks, as (cirrhosis). The third group received 1 cc kg-1 (CCl4) repeatedly administered into the peritoneal cavity, twice weekly, for 2 weeks, with Dulbecco’s Modified Eagle Medium (DMEM) (culture medium). The forth group received 1 cc kg-1 (CCl4) repeatedly administered into the peritoneal cavity, twice weekly, for 2 weeks, received CD133+ HSCs that treated (differentiated) with 20 μg mL-1 Hepatocytes Growth Factor (HGF). The fifth group received 1 cc kg-1 (CCl4) repeatedly administered into the peritoneal cavity, twice weekly, for 2 weeks, received CD133+ HSCs without HGF treated (undifferentiated). All rats were sacrificed 4 weeks after transplantation.
CD133+ BMSCs isolation and culture conditions: The BMSCs were collected from tibias and femurs from rats under anesthesia. The bone marrow cells were seeded onto a Hanks Buffer Salt Solution (HBSS). The cell suspension was filtered through a cell strainer (100 μm) to remove debris, Isolation of CD133+ BMSC using the CD133+ Cell Isolation Kit (Miltenyi Biotec). The positively and negatively selected cell fractions were assessed by Immunocytochemistry, using PE-conjugated anti-CD133+ antibody, following the previously described protocol (Zocco et al., 2011) and then CD133+ cells cultured in Dulbecco’s modified Eagle Medium (DMEM, Invitrogen), supplemented with Fetal Bovine Serum (FBS, 10%), streptomycin (50 μg mL-1) and amphotericin B (2.5 μg mL-1) in a 5% CO2 incubator at 37°C. After 2 days, non-adherent cells were removed with the medium and adherent cells were used as the BMSCs. The culture medium was changed twice a week and cells in the second passage were used for injection.
Twenty micrograms per milliliter HGF (Sigma-Aldrich) and 10 nM dexamethasone (Sigma-Aldrich) were added to the Medium in the priming culture to induce differentiation into hepatocyte. As a control, bone marrow cells were also cultured without HGF. During the cell culture with or without HGF, each medium change eliminated the floating cells and allowed the adherent cells to proliferate. The HSCs cultured for 2 weeks in the presence or absence of HGF were detached by trypsin treatment from the plate and 2×106 cells were injected via caudal vein in experimental groups. After 4 weeks the rats were killed, blood and liver tissue samples were collected.
Histological analysis and immunohistochemistry: At 28 days after injection, rats were sacrificed and the right lobe of liver tissues was removed for histological evaluation. The liver samples were fixed with 4% buffered formalin, embedded in paraffin and sectioned with a microtome (5-7 μm thick). Finally, the sections were subjected to Hematoxylin and Eosin (HE) and Masson Trichrome (MT) staining to determine the extent of liver inflammation and fibrosis development. The area of liver fibrosis was quantified with MT staining. Briefly the fibrotic area was assessed at a magnification of 40 using computer-assisted image analysis with Image J software. Ten fields were randomly selected for each group. Sections were subjected to immunoperoxidase biotin-avidin reaction in the Labelled Streptavidin Biotin method (LSAB). Sections were incubated at room temperature for 30 min with the following antibodies: The CD133+ rabbit polyclonal (dilution 1:100, Bio care Medical, CA), alpha-fetoprotein (AFP) rabbit polyclonal (dilution 1:200, Dako). Slides were counter stained with hematoxylin. Negative controls were performed using non-immunized rabbit or mouse serum and omitting the primary antibodies.
Immunocytochemical staining differentiated cells to hepatocyte: Cells were cultured in collagen-coated 6-well plates (Nippon Becton Dickinson, Tokyo, Japan) and fixed with 10% formaldehyde for 20 min. Cells were treated with 0.5% Triton X-100 for several minutes and then with blocking solution for 1 h. The cells were incubated with primary antibody, i.e., rabbit IgG anti-rat albumin (Cappel, Durham, NC, USA) diluted 1:200 with the blocking solution, for 1 h. The cells were mixed with the secondary antibody, goat F (ab) antirabbit IgG (H and L)-Fluorescein (Leinco Technologies Inc., St Louis, MO, USA) diluted 1:100 with the blocking solution and incubated for 2 h between each step, the cells were washed three times for 5 min each with Phosphate-buffered Saline (PBS). All steps were carried out at room temperature.
Serum albumin level analysis: Under anesthesia, blood specimens were collected by cardiac puncture. Serum albumin levels after 2 weeks of cell infusion were analyzed by using the Spotchem EZ SP-4430 dry chemical system (Arkray, Kyoto, Japan) and assessed using routine laboratory methods (Fuji drychem, Fuji Photo film, Co., Ltd., Kanagawa, Japan).
Statistical analysis: All statistical analyses was performed using the SPSS program 11.5 for Windows (SPSS, Chicago, IL, USA). Serum parameter data and fibrotic areas are presented as Mean±SD. Significant differences were determined using the Student’s t-test with p<0.05 considered statistically significant.
RESULTS
The present study aims to demonstrate the differentiation potential of HSCs CD133+ into hepatocytes after in vitro culture and in vivo engraftment into cirrhotic liver organ.
Characterization of CD133+ BMSCs: The adherent cells exhibited polymorphic and fibroblast-like morphology. The cells were positive for Hematopoietic Stem Cells (HSCs) specific markers CD133+ (Fig. 1).
Morphological observations of the HSCs: Isolated CD133+ cells were cultured in the presence or absence of HGF for 2 weeks.
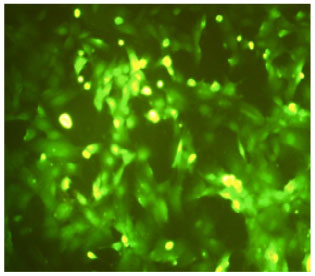 | |
| Fig. 1: | Immunofluorescent staining of rat HSCs. HSCs were positive for the Bone Marrow Stem Cell (BMSC) specific markers CD133+ |
Adherent cells started to proliferate on the 3rd day of culture and spindle-shaped cells with large and small clone then extended to form projections, regardless of the presence of HGF, at least for 8 days of culture (Fig. 2a). Cells were confluent in 2-3 week. When these cells were placed in a medium containing HGF medium for 2 week, spindle-shaped cells gradually transformed into hepatocytes-like round or polygonal cells and the ratio of nucleoplasm decreased. Adhesion ability weakened and was associated with a reduced number of cells in the HGF group medium at the time of medium change. In contrast, cells in the other groups maintained the spindle phenotype of BMSCs (Fig. 2b). A significant difference in cell growth between the HGF group medium with the other groups was observed.
Immunofluorescent staining: Differentiated hepatocytes-like round and polygonal cells were tested following HGF induction by immunofluorescence staining. The fluorescence was observed only in HGF-treated cells. Therefore, HSCs differentiated into hepatocyte-like cells in response to the culture being primed with HGF.
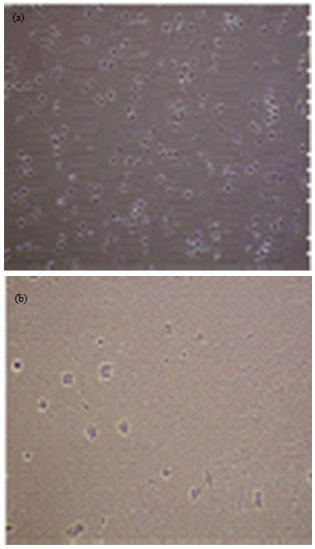 | |
| Fig. 2(a-b): | Rat BMSCs (CD133+) sub cultured and induced 2 week (a) Without different conditions and (b) Under HGF differentiate condition |
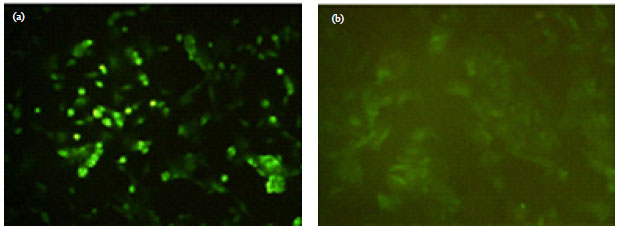 | |
| Fig. 3(a-b): | Immunofluorescent staining. Expression of FITC labeled albumin in hepatocyte-like cells, (a) HGF group and (b) Negative expression of FITC-labeled albumin (original magnification, 340) |
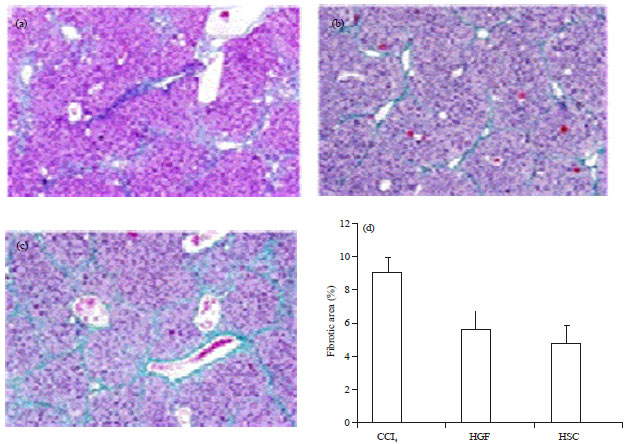 | |
| Fig. 4(a-d): | PhotomicrographsoflivertissueinCCl4 rat stained Masson-trichrome (a) Fibrotic (blue) area induced by CCl4 (b) Decreased afterHSCs (CD133+)treatedwith HGF transplantation, (c)UntreatedHGF and(d) Magnification; 20× quantifiedanalysisoffibrosis in different groups and (d) All values are expressed as ±SEM (*p<0.05) |
In the HGF group, we found FITC-labeled albumin green fluorescence (Fig. 3a). The non-induced cells in the untreated group displayed a negative reaction (Fig. 3b).
Improvement in liver fibrosis after HSC (CD133+) infusion: Liver fibrosis was evaluated by Masson Trichrome (MT). After 4 weeks, HSC (CD133+) transplanted liver exactly showed reduction of liver fibrosis compared to the group that treated with CCl4 alone and vehicle group. Quantitative analysis of liver fibrosis by image J software indicated that the percent of fibrotic area of liver in HSC which treated with HGF was significantly decreased as compared to other groups (Fig. 4 a, d). Immunohistochemical analysis showed, CD133+ positive cells were mainly observed in the portal region (Fig. 5).
Improvement of albumin production by HSCs transplantation: The normal level serum albumin in rat blood was 4.8±0.8 g dL-1.
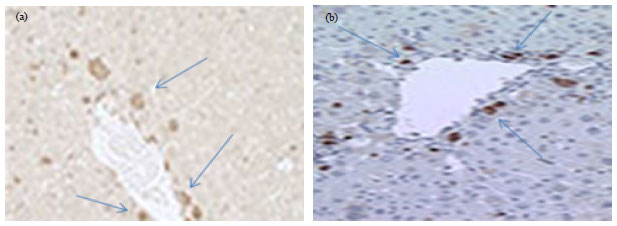 | |
| Fig. 5(a-b): | Immunohistochemical staining of HSCs injected in rat. (a) Magnification; 20×CD133+ cells are shown by arrows in portal region and (b) Liver after cell injection and the position of these cells |
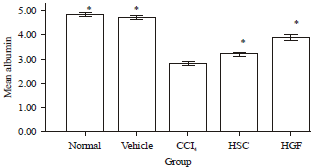 | |
| Fig. 6: | Recovery of serum albumin concentration in CCl4-injured rats caused by the transplantation of HSCs. Non-HGF-treated and HGF-treated HSCs were transplanted. Values are Means±SEM of five different animals. Asterisk indicates a significant difference from the values of rats that received a transplant with CCl4 (*p<0.05) |
As shown in Fig. 6, the albumin level decreased to 2.8±0.8 g dL-1 following the CCl4 treatment. Liver-injured rats did not completely recover normal serum albumin levels following the transplantation of non-HGF-treated HSCs (3.2±0.07 g dL-1). In contrast, the transplantation of HGF-treated HSCs into liver damaged rats further improved the serum albumin compared to non-HGF-treated HSCs (3.9±0.12 g dL-1). Although, it is not clear whether the transplanted HSCs differentiated into hepatocytes that produced albumin, or whether CCl4-damaged liver regenerated in response to the HGF-treated HSCs.
DISCUSSION
In this study CCl4 induced liver fibrosis that was ameliorated after transplantation of HSCs. It was believed that adult BMSCs might contain cells capable of differentiating into hepatocyte-like cells. Previous study (Wang et al., 2004) has suggested the expression of hepatocyte like cells in BMSCs after in vitro HGF induction. Research show that transplanted BMSCs could decrease collagen fibers and distinctly reduce liver fibrosis through high expression of matrix metalloproteinase (MMPs) and MMP-9 in particular. The reason for the strong expression of MMP-9 is still unknown (Sakaida et al., 2004). However, MMP-9 induction in BMSCs caused the release of soluble Kit-ligand which might be related to the transition of stem cells in the BMSC population to the proliferative niche (Heissig et al., 2005). The BMSCs improve liver fibrosis by expressing some factors such as MMP-9 (Lee et al., 2004) or releasing differentiation factors that differentiate and proliferate BMSCs in liver inflammation induced by injection of CCl4 (Sakaida et al., 2004). The BMSCs secret some of growth factors such as HGF, Nerve Growth Factor (NGF) and some cytokines have antiapoptotic activity in the liver and have the important role in the regeneration of the liver (Li et al., 2002; Baram et al., 2001). Activation of hepatic stellate cells cause more expression of collagen 1α1 that leads to liver fibrosis (Fang et al., 2004), BMSCs reduce the activity of stellate cells in liver injury (Trim et al., 2000). In this study, treated and untreated HSCs with HGF were used. These cells can release or activate these factors.
Effective migration of transplanted HSCs was limited to the injured liver in CCl4 group and reduction of liver fibrosis was observed only in CCl4 treated with HSCs transplantation. Therefore, HSCs might be particularly effective in resolving inflammatory fibrotic lesions, as previously reported (Kollet et al., 2003; Rabani et al., 2010; Zheng and Liang, 2008). It should be noted that differentiation of bone marrow cells into hepatocytes has also been reported (Neo et al., 2009; Petersen et al., 1999). Moreover, transplanted bone marrow cells have been shown to restore biochemical function in recipients with progressive liver failure and hepatic cells differentiated from bone marrow cells protect against or can ameliorate and CCl4-induced fibrosis in recipient rats. In this study, two kinds of HSCs differentiated into hepatocyte and not differentiated into hepatocyte prior to transplantation were used. Differentiated cells were more effective in reducing liver fibrous than undifferentiated cells. Therefore, HSCs differentiation might be necessary prior to transplantation. In conclusion, this study demonstrates that transplanted HSCs migrate into liver regions exhibiting severe inflammation and fibrosis. Liver fibrosis was significantly decreased in CCl4 rat that received transplantation with HSCs. These results suggest that HSCs have the potential to be an effective source of inducers that support liver regeneration.
Transplanted HSCs have been observed to differentiate into hepatocyte and subsequently into albumin-producing hepatocytes, many reports have demonstrated improvements in serum albumin following BMSCs transplantation therapy (Terai et al., 2006).
Serum albumin was lower in the CCl4 injury group but was significantly higher in the HGF treatment group and the HSCs without treatment group. The HGF group was significantly higher than in HSCs Group. These results showed that Transplanted HSCs have been observed to differentiate into hepatoblasts and subsequently into functional albumin-producing hepatocytes that cause improvement of liver function. Many reports have demonstrated improvements in serum albumin following BMSCs transplantation therapy (Aziz et al., 2007; Terai et al., 2006).
CONCLUSION
This study demonstrates that transplanted HSCs migrate into liver regions, substantially decrease in the inflammation and fibrosis. Liver fibrosis was significantly decreased in CCl4 rat that received transplantation with HSCs. These results suggest that HSCs have the potential to be an effective source of inducers that support liver regeneration.
ACKNOWLEDGMENTS
This study was supported by Grant of the Semnan University of Medical Sciences (302).
REFERENCES
- Abdel Aziz, M.T., H.M. Atta, S. Mahfouz, H.H. Fouad and N.K. Roshdy et al., 2007. Therapeutic potential of bone marrow-derived mesenchymal stem cells on experimental liver fibrosis. Clin. Biochem., 40: 893-899.
CrossRefDirect Link - Alison, M.R., R. Poulsom, R. Jeffery, A.P. Dhillon and A. Quaglia et al., 2000. Cell differentiation: Hepatocytes from non-hepatic adult stem cells. Nature, 406: 257-257.
Direct Link - Baram, D., G.G. Vaday, P. Salamon, I. Drucker, R. Hershkoviz and Y.A. Mekori, 2001. Human mast cells release metalloproteinase-9 on contact with activated T cells: Juxtacrine regulation by TNF-α. J. Immunol., 167: 4008-4016.
Direct Link - Bataller, R. and D.A. Brenner, 2005. Liver fibrosis. J. Clin. Invent., 115: 209-218.
CrossRefPubMedDirect Link - Dominici, M., K. le Blanc, I. Mueller, I. Slaper-Cortenbach and F. Marini et al., 2006. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy, 8: 315-317.
CrossRefPubMedDirect Link - Fang, B., M. Shi, L. Liao, S. Yang, Y. Liu and R.C. Zhao, 2004. Systemic infusion of FLK1+ mesenchymal stem cells ameliorate carbon tetrachloride-induced liver fibrosis in mice. Transplantation, 15: 83-88.
PubMedDirect Link - Friedman, S.L., 2008. Hepatic stellate cells: Protean, multifunctional and enigmatic cells of the liver. Physiol. Rev., 88: 125-172.
CrossRefDirect Link - Heissig, B., Y. Ohki, Y. Sato, S. Rafii, Z. Werb and K. Hattori, 2005. A role for niches in hematopoietic cell development. Hematology, 10: 247-253.
CrossRefDirect Link - Kollet, O., S. Shivtiel, Y.Q. Chen, J. Suriawinata and S.N. Thung et al., 2003. HGF, SDF-1 and MMP-9 are involved in stress-induced human CD34+ stem cell recruitment to the liver. J. Clin. Invest., 112: 160-169.
CrossRefDirect Link - Korbling, M., R.L. Katz, A. Khanna, A.C. Ruifrok and G. Rondon et al., 2002. Hepatocytes and epithelial cells of donor origin in recipients of peripheral-blood stem cells. New Engl. J. Med., 346: 738-746.
CrossRefDirect Link - Lai, C.L., V. Ratziu, M.F. Yuen and T. Poynard, 2003. Viral hepatitis B. Lancet, 362: 2089-2094.
CrossRefDirect Link - Lee, K.D., T.K.C. Kuo, J. Whang-Peng, Y.F. Chung and C.T. Lin et al., 2004. In vitro hepatic differentiation of human mesenchymal stem cells. Hepatology, 40: 1275-1284.
CrossRefPubMedDirect Link - Li, Y., J. Chen, X.G. Chen, L. Wang and S.C. Gautam et al., 2002. Human marrow stromal cell therapy for stroke in rat: Neurotrophins and functional recovery. Neurology, 59: 514-523.
PubMed - Neo, S., T. Ishikawa, K. Ogiwara, N. Kansaku and M. Nakamura et al., 2009. Canine bone marrow cells differentiate into hepatocyte-like cells and placental hydrolysate is a potential inducer. Res. Vet. Sci., 87: 1-6.
CrossRefDirect Link - Okamoto, R., T. Yajima, M. Yamazaki, T. Kanai and M. Mukai et al., 2002. Damaged epithelia regenerated by bone marrow-derived cells in the human gastrointestinal tract. Nat. Med., 8: 1011-1017.
CrossRefDirect Link - Petersen, B.E., W.C. Bowen, K.D. Patrene, W.M. Mars and A.K. Sullivan et al., 1999. Bone marrow as a potential source of hepatic oval cells. Science, 284: 1168-1170.
CrossRefDirect Link - Piscaglia, A.C., C. di Campli, G. Gasbarrini and A. Gasbarrini, 2003. Stem cells: New tools in gastroenterology and hepatology. Dig. Liver Dis., 35: 507-514.
CrossRefDirect Link - Piscaglia, A.C., T. Shupe and A. Gasbarrini, 2003. Microarray RNA/DNA in different stem cell lines. Curr. Pharmaceut. Biotechnol., 8: 167-175.
CrossRefDirect Link - Pittenger, M.F., A.M. Mackay, S.C. Beck, R.K. Jaiswal and R. Douglas et al., 1999. Multilineage potential of adult human mesenchymal stem cells. Science, 284: 143-147.
CrossRefPubMedDirect Link - Poldervaart, J.H., R.P. Favier, L.C. Penning, T.S.G.A.M. van den Ingh and J. Rothuizen, 2009. Primary hepatitis in dogs: A retrospective review (2002-2006). J. Vet. Internal Med., 23: 72-80.
CrossRefDirect Link - Poynard, T., M.F. Yuen, V. Ratziu and C.L. Lai, 2003. Viral hepatitis C. Lancet, 362: 2095-2100.
PubMed - Ratajczak, M.Z., M. Kucia, R. Reca, M. Majka, A. Janowska-Wieczorek and J. Ratajczak, 2004. Stem cell plasticity revisited: CXCR4-positive cells expressing mRNA for early muscle, liver and neural cells hide out in the bone marrow. Leukemia, 18: 29-40.
CrossRefDirect Link - Sakaida, I., S. Terai, N. Yamamoto, K. Aoyama, T. Ishikawa, H. Nishina and K. Okita, 2004. Transplantation of bone marrow cells reduces CCl4-induced liver fibrosis in mice. Hepatology, 40: 1304-1311.
CrossRefDirect Link - Terai, S., T. Ishikawa, K. Omori, K. Aoyama and Y. Marumoto et al., 2006. Improved liver function in patients with liver cirrhosis after autologous bone marrow cell infusion therapy. Stem Cells, 24: 2292-2298.
CrossRefDirect Link - Theise, N.D., M. Nimmakayalu, R. Gardner, P.B. Illei and G. Morgan et al., 2000. Liver from bone marrow in humans. Hepatology, 32: 11-16.
CrossRefDirect Link - Trim, N., S. Morgan, M. Evans, R. Issa and D. Fine et al., 2000. Hepatic stellate cells express the low affinity nerve growth factor receptor p75 and undergo apoptosis in response to nerve growth factor stimulation. Am. J. Pathol., 156: 1235-1243.
CrossRefDirect Link - Wang, P.P., J.H. Wang, Z.P. Yan, M.Y. Hu and G.K. Lau et al., 2004. Expression of hepatocyte-like phenotypes in bone marrow stromal cells after HGF induction. Biochem. Biophys. Res. Commun., 320: 712-716.
CrossRefDirect Link - Zocco, M.A., A.C. Piscaglia, F. Giuliante, V. Arena and M. Novi et al., 2011. CD133+ stem cell mobilization after partial hepatectomy depends on resection extent and underlying disease. Dig. Liver Dis., 43: 147-154.
CrossRefDirect Link