
Farhana Aminuddin
Department of Nutrition and Dietetics, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Faisal Ali
Department of Nutrition and Dietetics, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Amin Ismail
Department of Nutrition and Dietetics, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Chong Pei Pei
Department of Biomedical Sciences, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Muhajir Hamid
Department of Microbiology, Faculty of Biotechnology and Bimolecular Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 4 | Page No.: 309-317







ABSTRACT
Cocoa, the fruit of Theobroma cacao plant is traditionally used in the folk medicine as a pharmaceutical for blood pressure reduction and cardiovascular diseases prevention. The nuclear receptor peroxisome proliferator-activated receptor gamma (PPAR-γ) is widely known to improve insulin sensitivity and is thereby being used as a major drug target for the treatment of type 2 diabetes mellitus. The present study investigated the anti-diabetic/anti-obesity effects of cocoa polyphenol-rich extract (CoPE) using obese-diabetic rats (Sprague Dawley rats). Sprague Dawley rats received either normal diet, high-fat diet or high-fat diet with additional cocoa polyphenols for 8 weeks. After the end of the treatment, body weight, plasma glucose and insulin were measured. Furthermore, mRNA and protein levels of PPARγ were measured in skeletal muscle and white adipose tissue. Compared to the high-fat diet group, increases in body weight, plasma glucose and insulin were significantly suppressed for CoPE-treated groups. Furthermore, compared to the high-fat diet group, the PPARγ mRNA level was significantly higher in both skeletal muscle and white adipose tissue for CoPE groups. Protein expression of PPARγ in CoPE groups was also significantly higher compared to the high-fat diet group. In conclusion, the anti-diabetic mechanism of actions of CoPE along with metformin hypoglycemic drug is partially attributed to increase expression of PPARγ in skeletal muscle and adipose tissue. These results suggest that CoPE could be a useful phytomedicine agent for alleviating insulin resistance.
PDF Abstract XML References Citation
Received: January 10, 2015;
Accepted: March 10, 2015;
Published: April 18, 2015
How to cite this article
Farhana Aminuddin, Faisal Ali, Amin Ismail, Chong Pei Pei and Muhajir Hamid, 2015. Cocoa Polyphenol-Rich Extract Enhances the Expression Levels of PPAR-γ
in the Skeletal Muscle and Adipose Tissue of Obese-Diabetic Rats Fed a High-Fat
Diet. International Journal of Pharmacology, 11: 309-317.
DOI: 10.3923/ijp.2015.309.317
URL: https://scialert.net/abstract/?doi=ijp.2015.309.317
DOI: 10.3923/ijp.2015.309.317
URL: https://scialert.net/abstract/?doi=ijp.2015.309.317
INTRODUCTION
Diabetes mellitus is a metabolic disorder with multiple causes and etiology that is characterized by hyperglycemia and insulin resistance. Type 2 diabetes is globally increased and comprises over 90% of diabetes cases (Gershell, 2005; Wild et al., 2004). Diabetic patients frequently demand to use natural medicines with anti-diabetic properties due to the side effects associated with the use of insulin and conventional drugs. Utilization of medicinal plants for the management and treatment of diabetes mellitus dates back to the Ebers papyrus from approximately 1550 B.C (Holman and Turner, 1991; Rao et al., 1997).
Scientific research has proven the efficacy of many of medicinal plants, some of which are remarkably effective (Grover et al., 2002). Theobroma cacao is a small evergreen tree belonging to the family Malvaceae that is widely found in the deep tropical region of the Americas. Medicinal uses of cocoa were reported in Europe and New Spain from 16th to early 20th century (Dillinger et al., 2000). Cocoa extract is rich in polyphenolic compounds, namely catechin and epicatechin, dimer, trimer, tetramer and up to tetradecamer (Maleyki and Ismail, 2010). Moreover, methylxanthines, namely caffeine, theobromine and theophylline, have also been identified in cocoa (Kelm et al., 2006). The significant amounts of bioactive compounds that have health-promoting properties have become an area of interest. Many researchers have demonstrated the health benefits of cocoa polyphenol-rich extract based on in vivo and in vitro studies (Amin et al., 2004; Kurosawa et al., 2005; Bisson et al., 2008; Maleyki and Ismail, 2010; Ruzaidi et al., 2005; Ali et al., 2014a). The mechanistic action towards metabolic syndrome is believed to be linked to PPAR-γ.
PPAR-γ is a member of the nuclear receptor superfamily of ligand-activated transcription factors, a family that includes the steroid, retinoid and thyroid hormone receptors (Keller et al., 2000). The gene for PPAR-γ was originally cloned in both homology-based screens for novel nuclear receptors and as part of a search for transcription factors that regulate adipocyte differentiation specific genes. It is also involved in fatty acid storage and is activated by natural ligands such as leukotrienes, prostaglandins and fatty acids. PPAR-γ expression can either increase or decrease the transcription of genes by for insulin sensitivity and adipogenesis, adipocyte differentiation, inflammatory process and cell proliferation (Kodera et al., 2000).
Previous studies demonstrated that cocoa beans possess hypoglycemic and hypolipidemic properties by reducing blood glucose and the lipid profiles of rats with induced diabetes (Amin et al., 2004; Ruzaidi et al., 2005; Maleyki and Ismail, 2010) indicated that cocoa supplementation could reduce short-term blood glucose as assessed by an oral glucose tolerance test and had a protective effect by reducing lipid peroxidation levels. Moreover, the health benefits of cocoa polyphenols have been shown in human clinical trials (Cooper et al., 2008). These trials clearly indicated that cocoa supplementation could reduce blood glucose as well as the lipid profiles of diabetic animals. However, limited studies have been performed on diabetic-obese rats using a combination of high-fat diet and a low dose of streptozotocin (STZ) that mimicked human diabetes in relation to PPAR-γ expression. Hence, this study was conducted to evaluate the phytophramacological activity of CoPE along with metformin hypoglycemic drug in retarding the progress of diabetic development through PPARγ activation in diet-induced Ob-db rats.
MATERIALS AND METHODS
Materials: Cocoa powder was purchased from KL Kepong (Cocoa Products) Sdn. Bhd, Selangor, Malaysia. Rats were purchased from A. Sapphire Enterprise, Sri Kembangan, Selangor, Malaysia. Raw materials for diet formulation were obtained from ICB Biolab, United States.
Chemicals and reagents: Chemicals, reagents and kits for analysis were purchased from Sigma Chemical Co. (US), Wako Diagnostic (Japan), Randox Chemical (UK), GE Healthcare (UK), BioRad Laboratory (US) and Cayman Chemical (US). Antibodies for Western blotting were purchased from Santa Cruz (CA, USA). Kits, primers and reagents for RT-PCR were purchased from Invitrogen (USA) and Promega (USA).
Methods: Preparation of cocoa polyphenol-rich extracts (CoPE). Cocoa extract was prepared according to our previous studies (Maleyki and Ismail, 2010; Ali et al., 2013). Briefly, cocoa extract was stirred with 80% (v/v) ethanol for 2 h at room temperature and then filtered using filter paper (Whatman paper No. 4). The ethanolic residue was removed from the cocoa extract using a rotary evaporator (Buchi Rotavor R-200, Flawil, Switzerland) at 55°C. The resulting extract was lyophilized using a freeze dryer (The Virtis Company Inc., Gardiner, NY, USA) at -45°C and 120 bar. The cocoa extract was analyzed for its bioactive compounds using a High Performance Liquid Chromatography (HPLC) system (Agilent 1100, Palo Alto, USA) equipped with an auto injector, degasser, quaternary pump and DAD. A reversed phase C-18 column (Alltech, Licosphere, United States) (250×4 mm, 5 μm I.D) was used for the separation of bioactive compounds with a gradient elution of (A) water-trifluoroacetic acid (99.9: 0.1, v/v) and (B) acetonitrile-trifluoroacetic acid (99.9: 0.1, v/v) (Natsume et al., 2000). A flow rate of 0.8 mL min-1 with a linear gradient elution of 0-10% (A) for 5 min, 10-25% (A) for 25 min and 25-100% (A) for 5 min was used for analysis. The amount of catechin, epicatechin, caffeine and theobromine (mg g-1 fraction) was quantified on the basis of external standards (100-1000 μg mL-1).
Animal study: Forty-eight Sprague Dawley rats (weighing 150-200 g) were purchased from A Sapphire Enterprise, Selangor. Experimental design approval was obtained from the Animal Care and Use Committee (ACUC: UPM/FPSK/PADS/BR-UUH/00344) of the Faculty of Medicine and Health Sciences, Universiti Putra Malaysia. The rats were acclimatized for 1 week at room temperature (26-28°C) under a 12 h dark: 12 h light cycle. All the rats were given normal diets and distilled water ad libitum. The rats were randomly divided into 6 groups with 8 rats each (n = 8). This study was carried out for 20 weeks and diet intake, body weight and height (anal to basal length) and plasma glucose levels were recorded. Obesity was induced in Groups 3-6 using a high-fat adjusted diet consisting of 49% fat, 32% carbohydrate and 19% protein (Maleyki and Ismail, 2010; Ali et al., 2014b) calories (kcal) administered for three months. Normal rats were given normal rat chow (14% fat, 61% carbohydrate and 25% protein calories). Body weight was recorded weekly to confirm the development of obesity. The obese model was developed after 12 weeks of the high-fat diet. Rats were injected with 35 mg kg-1 body weight streptozotocin to induce diabetes. The plasma blood glucose was checked after three days after STZ injection and later checked weekly to ensure that the rats developed diabetes. Rats with fasting plasma blood glucose levels of >120 mg dL-1 were used in this study. Six hundred milligrams CoPE per kilogram of body weight was given to the Ob-db rats in Group 5. The does was selected based on previous studies by Amin et al. (2004) and Maleyki and Ismail (2010). A conventional treatment (Metformin) at dosage of 500 mg kg-1 body weight (Adam et al., 2010) was given to the Ob-db rats in Group 6 as a positive control. Cocoa extract and metformin were given daily by gastric intubation using a force-feeding needle.
Oral glucose tolerance test: At the end of the study, rats from each group were subjected to an oral glucose tolerance test (OGTT). The OGTT was performed to determine the short term effect of cocoa extract administration on Ob-db rats (Group 5). Glucose (2 g kg-1) was intraperitoneally administered to rats fasted for 16 h. Blood samples were collected through the tail vein without anesthesia 0, 15, 30, 60, 90 and 120 min after the administration of glucose solution. The results were expressed as mmol L-1 and the curve response of each group was plotted. Then, the Area Under Curve (AUC) was determined using GraphPadPrism version 5.
Blood collection for biochemical analysis: Blood was collected through the retro-orbital plexus under general anesthesia at week 13 (after STZ injection for all groups except ND and Ob) and week 21 (after cocoa extract and metformin supplementation in Ob-db+cocoa and Ob-db+ metformin). Approximately 7 mL of blood was collected in different blood collection tubes. Blood for plasma glucose testing was collected in tubes containing potassium oxalate as an antiglycolysis agent. Blood for insulin and lipid profiles was collected in tubes containing lithium heparin as an anticoagulant. Blood samples were centrifuged (1000×g for 15 min at 4°C) and plasma was stored at -80°C until use.
Determination of lipid profiles and plasma glucose and insulin levels: Fasting plasma glucose and lipid profiles were determined using commercially available kits from ROCHE, UK. The optical density developed during this assay was measured using a chemical analyzer (Hitachi 902 Automatic Analyzer, Hitachi, Japan) and the results were expressed as mmol L-1. The fasting plasma insulin level was determined using the Rat Insulin Biotrak Assay System.
PPAR-γ protein expression (SDS-PAGE analysis and Western blot): Tissues were excised, weighed and immediately frozen in liquid nitrogen and stored at -80°C until processing. Protein was extracted using complete polytron homogenization in extraction buffer containing protease inhibitor followed by 10,000 rpm centrifugation at 4°C for 30 min. Protein concentration was assessed by the bicinchoninic acid technique (Pierce, Rockford, IL). Electrophoresis was then performed using the method described by Ikarashi et al. (2011). Basically, the prepared protein samples were resolved by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE). After electrophoresis, the resulting gel was electro blotted onto a polyvinylfluoride membrane (Millipore Corp., Bedford, MA) followed by the blocking of non-specific binding sites with 1.0% skim milk for 2 h at 4°C on a belly shaker. The membranes were then immunoblotted using an anti-human PPAR-γ rabbit polyclonal primary antibody (1:500 dilution; Santa Cruz Biotechnology, USA in blocking solution) overnight at 4°C. After that, the membranes were washed using TBS-Tween (20 mMTris-HCl, 137 mMNaCl and 0.1% Tween 20, pH 7.6). The resulting membranes were then incubated with anti-rabbit secondary antibody conjugated with horseradish peroxidase (Santa Cruz Biotechnology, CA, USA) for 2 h. The specific immune complexes corresponding to PPAR-γ were visualized by enhanced chemiluminescence (Gel Imagine CHEMI-SMART-3126, France). PPAR-γ protein levels were normalized to β-actin. The membranes were reprobed with anti-actin primary antibody (Santa Cruz Biotechnology, CA, USA) after stripping with 5 mL stripping buffer and incubation for 15 min at 50°C. The membranes were washed, re-incubated with the same secondary horseradish peroxidase antibody and detected using the same procedure as described above.
Statistical analysis: The data is expressed as the Mean±Standard deviation. Multi-group comparisons in the in vivo study were performed using ANOVA and Duncan’s test. The statistical analysis of protein expression was performed using Image J software. The mRNA expression analysis was performed using Rotorgene 600 series software. Analyses were performed using the SPSS statistical software package (SPSS Inc., Chicago, IL). A p-value <0.05 (p<0.05) was considered significant.
RESULTS
CoPE improves plasma and oral glucose tolerance in Ob-db rats: CoPE was extracted and showed a high phenolic content of 129.9 mg GAE/g extract. The high phenolic CoPE content is in agreement with the findings of Maleyki and Ismail (2010). The effects of CoPE were first demonstrated on plasma glucose by comparing the levels before and after treatment. Table 1 shows that the level of plasma glucose in Ob-db+cocoa and Ob-db+metformin significantly (p<0.05) decreased, with a value of 7.1 and 7.82 mmol L-1, respectively. However, the levels remained higher in STZ-injected groups (ND and Ob-db), with a range of 22.1-25.3 mmol L-1 and in non-injected groups (N and Ob), with a range of 7.4-8.0 mmol L-1 at the end of the study. As shown in Fig. 1, treatment for more than 10 weeks with CoPE inhibited the increase in glucose levels in Ob-db+cocoa rats 30-120 min after glucose loading while it had no effect on Ob-db control rats.
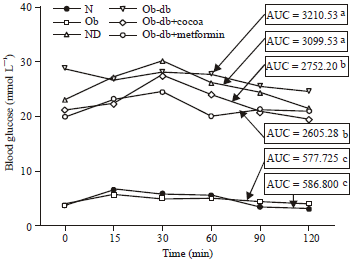 | |
| Fig. 1: | Oral Glucose Tolerance Test (OGTT) of the rats at the end of animal study. Area under curve (AUC, mmol L-1) values with different letters are significantly different at the level of p<0.05. N: Normal, Ob: Obese, ND: Normal diabetic, Ob-db, obese-diabetic; Ob-db+cocoa, Ob-db+600 mg cocoa kg-1 body weight; Ob-db+metformin; Ob-db+500 mg metformin/kg body weight, aSignificantly different from treated group bSignificantly different from untreated group cSignificantly different from both treated and untreated groups |
| Table 1: | Body weight, plasma glucose, insulin and lipid profiles of the experimental rats at the end of the animal study |
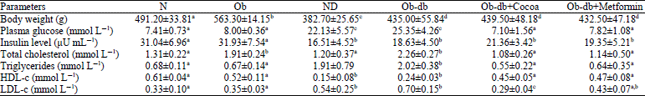 | |
| Values are expressed as the Mean ± SD. Means with different letters within a column are significantly (p< 0.05) different. Baseline values are after receiving a high-fat diet in all groups except N and ND and after STZ injection in diabetic group (ND, Ob-db, Ob-db + cocoa and Ob-db + metformin) and final values are after treatment with CoPE and metformin supplementation | |
CoPE improves insulin levels and lipid profiles of Ob-db rats: Improvement in plasma insulin sensitivity and lipid profile status resulted in a considerable change in diabetic type 2 conditions. After 8 weeks of supplementation with CoPE, the insulin level was not significantly increased compared to the Ob-db control group. However, the total cholesterol, triacylglyceride and LDL-c levels decreased significantly (p<0.05) under the influence of CoPE and metformin treatment. Moreover, CoPE and metformin treated animals also showed a significantly elevated HDL-c level (p<0.05).
PPARγ mRNA expression (Quantitative real-time PCR): RNA from adipose tissue was extracted using the RNeasy® Lipid Tissue Mini Kit according to the manufacturer’s protocol. The resulting solution was diluted 50-fold using TE buffer. Total RNA from isolated skeletal muscle was extracted using the RNeasy® Fibrous Tissue Mini Kit (Qiagen, USA) according to the manufacturer’s instructions. The RNA was then treated with RQ1 DNase (Qiagen, USA) at 37°C for 30 min followed by extraction with phenol-chloroform and ethanol precipitation. The quantity and quality of total RNA (μg mL-1) were checked by measuring the absorbance using a Biophotometer (Eppendorf). First-strand cDNA was generated using SuperScript II Quantitect® Reverse Transcription Kit (Qiagen, USA). PCR was performed on a thermocycler. cDNA samples were diluted 1:20 and 10 μL of the diluted cDNA was used in each 25 μL real-time PCR reaction using the QuantiFastTM SYBR®Green PCR Kit (Qiagen, USA) with a Rotor Gene 6000 Real Time PCR System and the Absolute Quantitation method. The genes examined were PPAR-γ (156 bp; sense: 5’-CAT TTT TCA AGG GTG CCA GT-3’ and antisense: 5’-GAG GCC AGC ATG GTG TAG AT-3’) and β-actin (348 bp; sense: 5’-CCG ACA GCA GTT GGT GGA GC-3’ and antisense: 5’- GGT CTC AAG TCA GTG TAC AG-3’). The results of quantitative PCR analysis were analyzed using the software supplied with the Rotor Gene 6000 System. A standard curve for each primer set was generated in each assay and used to derive the copy number of target sequences in unknown samples. β-actin was selected as an internal standard. All raw data was expressed as the ratio of the selected gene to β-actin.
CoPE enhances PPAR-γ mRNA expression in the adipocytes and the skeletal muscle of Ob-db rats: The SYBR Green dye assay was used to analyze PPAR-γ expression. The level of gene expression was determined by the 2–ΔΔCT method which represents the normalized target gene expression that equals the mean fold change in gene expression. The data tabulated in Table 2 showed the level of gene expression in different groups of rats in both adipose tissue and skeletal muscle. Based on the mean fold change, Ob-db+cocoa group had significantly (p<0.05) increased PPAR-γ gene expression (overexpression) in both adipose tissue and skeletal muscle compared to the other groups. Ob-db and Ob-db+metformin rats also exhibited significantly (p<0.05) increased levels of PPAR-γ gene expression in both adipose tissue and skeletal muscle. However, the N and Ob groups exhibited a lower level of PPAR-γ gene expression in both skeletal muscle and adipose tissue. The ND group had the lowest PPAR-γ gene expression in skeletal muscle but no data were obtained for ND adipose tissue because no sample was collected in the chronic diabetic condition. The gene expression levels can also be expressed using the log cDNA copy number (Fig. 2) for each sample of the different rat groups. There was no significant difference in the levels of PPAR-γ gene expression calculated by the mean fold change or log cDNA copy number.
CoPE enhances PPAR-γ protein expression in the adipocytes and skeletal muscle of Ob-db rats: PPAR-γ protein expression was determined by Western blotting as shown in Fig. 3. The effect of CoPE (600 μg mL-1) on both the adipose tissue and skeletal muscle of rats showed an increase in PPAR-γ protein expression compared to the N group. The group receiving metformin (500 μg mL-1) (positive control) showed a similar PPAR-γ expression level compared to the CoPE group in adipose tissue but was slightly lower in skeletal muscle. Ob-db rats exhibited significantly (p<0.05) enhanced PPAR-γ protein expression in both adipose tissue and skeletal muscle, whereas the control and Ob groups showed little effect.
| Table 2: | Data analysis using the 2–ΔΔCT method. The fold change in expression of the target gene (PPAR-γ) relative to the internal control gene (β-actin) was studied. The samples were analyzed using real-time quantitative PCR (Rotorgene 600 software) and the CT data were imported into Microsoft Excel. The mean fold change in the expression of the target gene was calculated (ΔΔCT = ΔCT sample-ΔCT calibrator) where 2-ΔΔCT = normalized target gene expression = mean fold change in gene expression.(A, adipose tissue; B, skeletal muscle) |
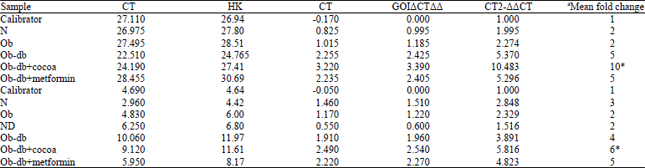 | |
| aRegulation of target gene (PPAR-γ), *Most upregulated mRNA of target gene | |
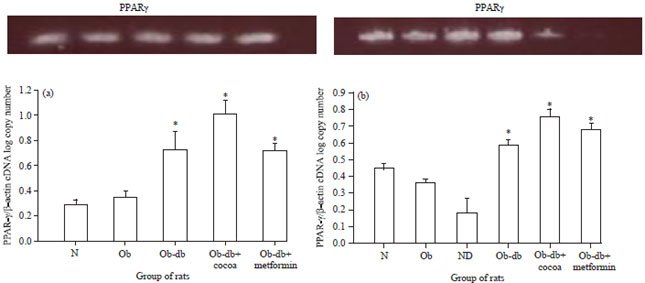 | |
| Fig. 2(a-b): | Mean mRNA levels PPAR-γ in the, (a) Adipose tissue and (b) Skeletal muscle of different group of rats. The cDNA values were calculated and the amounts shown (log copy number of cDNA) represent the mean of values determined in triplicate. The mean values of cDNA copies were corrected with respect to the reference gene (β-actin). The presence of a single RT-PCR product was verified by 2% gel electrophoresis for its specificity. *p<0.05 vs. normal (N) |
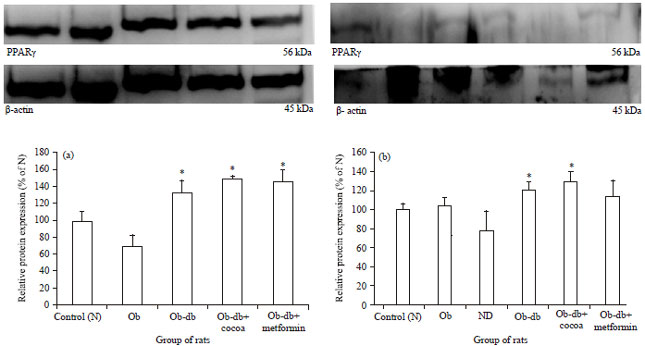 | |
| Fig. 3(a-b): | Expression of PPAR-γ protein in rat, (a) Adipose tissue and (b) Skeletal muscle. The protein extracts were subjected to immunoblotting by the PPAR-γ antibody and levels were normalized β-actin. Levels in the control were arbitrarily assigned a value of 100.0. All values are significantly different at the level of p<0.05. Protein contents were determined using the bicinchoninic acid method with BSA as a standard. *p<0.05 vs. normal (N) |
The identity of the band was confirmed by its apparent molecular weight. There was a significantly (p<0.05) lower expression of PPAR-γ protein in the skeletal muscle of the ND group compared to the N group. No comparison to the ND group can be made because no adipose tissues were collected in the diabetic normal condition. Overexpression of PPAR-γ protein in the CoPE treatment group suggested that CoPE could be a good regulator of glucose and lipid metabolism.
DISCUSSION
The partial mechanism behind the phytopharmacological and the pharmacological actions of both CoPE and metformin, respectively, is simultaneously evaluated in the present study. The expression level of PPAR-γ has been considered as a therapeutic target to improve insulin resistance (Furnsinn and Waldhausl, 2002). In the last decade, a new anti-diabetic drug, thiazolidinedione (TZDs), has been discovered as a PPAR-γ molecular target and is being used clinically; it appears to act by increasing insulin-stimulated glucose uptake in adipose tissue and skeletal muscle (Otto et al., 2002; Ali et al., 2014a). The side effects of conventional drugs have caused researchers around the world to attempt to find a natural and traditional treatment of type 2 diabetes mellitus (Bailey and Day, 1989), case reports on the use of traditional plant medicines have indicated improvements in quality of life as well as prolongation of life in diabetic patients. Medicinal plants such as Averrhoa bilimbi leaf extract possess definite hypoglycemic, hypotriglyceridemic, anti-atherogenic and anti-lipid peroxidative properties in STZ-diabetic rats (Pushparaj et al., 2000; Baba et al., 2007; Bnouham et al., 2006; Shirwaikar et al., 2004) also reported that Annona squamosa leaf extract has a protective effect via the reduction of body weight, plasma glucose and serum insulin in streptozotocin-nicotinamide type 2 diabetic rats. In addition, American berry and Gymnema yunnanense extract also exhibited anti-hyperglycemic activity in a diabetes model (Xie et al., 2002, 2003).
The present study results indicated that Ob-db rats treated with 600 mg CoPE/kg body weight can effectively control hyperglycemia and hyperinsulinemia by significantly reducing plasma glucose and improving insulin sensitivity as well as lipid profiles (Table 1). Research by Amin et al. (2004) and Ruzaidi et al. (2005) demonstrated that cocoa extract possesses hypoglycemic and hypolipidemic properties by reducing the blood glucose and lipid profiles of rats with induced diabetes. In addition, a study by Maleyki and Ismail (2010) indicated that cocoa supplementation could reduce short-term blood glucose as assessed by an oral glucose tolerance test. Moreover, dietary supplementations with cocoa liquor proanthocyanidins dose-dependently prevent the development of hyperglycemia in diabetic obese mice (Tomura et al., 2007).
The major polyphenolic compounds in CoPE can act as natural PPAR-γ ligands. This effect on the protein and mRNA expression of PPAR-γ in the adipose tissue and skeletal muscle of Ob-db rats was demonstrated here. The present study showed that CoPE and metformin possess an effective role in preventing the progress of type 2 diabetes by improving lipid metabolism and glucose homeostasis through the upregulation of PPAR-γ expression. Adipose tissues showed higher PPAR-γ expression levels compared to skeletal muscle; this reveals the importance of PPAR-γ in adipogenesis and adipocyte remodeling. According to Guo and Tabrizchi (2006), the upregulation of PPAR-γ in adipose tissue results in an increase in glucose transporter-4 (GLUT4) and c-Cbl associating protein (CAP), both of which are key in enhancing glucose uptake and translocation GLUT4 to the cell surface. The muscle is the primary organ for insulin-stimulated glucose disposal which regulates glucose homeostasis. Significant increases were observed in PPAR-γ expression levels. Cha et al. (2001) stated that increased PPAR-γ expression correlates with enhanced insulin-stimulated glucose uptake into the muscle mediated by translocation of GLUT4 to the cell membrane and increased insulin-stimulated P13K activity. The significant increase in PPAR-γ expression of muscle in CoPE-treated Ob-db rats was most likely due to augmentation of insulin-stimulated glucose uptake.
PPAR-γ is also known as a key gene in diabetes and plays an important role in maintaining homeostasis in glucose metabolism. According to Willson et al. (2000), PPAR-γ activation is associated with improved insulin sensitivity. This effect reflects the main mechanism underlying the anti-diabetic efficacy of PPAR-γ agonists. The present results showed that CoPE dose-dependently enhanced PPAR-γ mRNA and protein expression in both the adipose tissue and skeletal muscle of Ob-db rats compared with rats fed a normal diet (Fig. 2 and 3). According to Huang et al. (2005), improved glucose tolerance in ZDF rats via PPAR-γ induction was associated with improved insulin receptor sensitivity. In the same context, a link between improvement of insulin sensitivity and blood glucose reduction as well as upregulation of PPAR-γ expression has also been shown, most likely due to ligand-binding activation of PPAR-γ (Yin et al., 2009). Polyphenolic compounds in CoPE act as high-affinity ligands for PPAR-γ and as insulin sensitizers. In addition, the activation of PPAR-γ in adipose tissue and skeletal muscle by its endogenous ligand could be required to maintain the insulin sensitivity of these tissues.
Importantly, this study has revealed the phytotherapeutic effects of cocoa polyphenolic extract and pharmacological activity of metformin by elevating the expression of PPAR-γ in both adipose tissue and skeletal muscle. This result is particularly positive because cocoa is a natural product that could possess anti-diabetic activity without any long-term side effects compared to conventional drugs. The cocoa extract seems to offer potential as a plant extract that is useful for alleviating metabolic syndrome. Metformin also has the same effects as cocoa extract; however, a number of patients may experience some side effects from taking metformin. The most common side effects are gastrointestinal upset, including nausea, vomiting, diarrhea, indigestion, stomach discomfort and headache. A rare but serious side effect is lactic acidosis which can be life-threatening. Moreover, people who take metformin over the long term can develop vitamin B12 deficiency. All the side effects of metformin are why cocoa-derived polyphenols may be a good potential therapeutic agent against diabetes in the future (Staels and Fruchart, 2005; Gershell, 2005).
Overall, in this study we investigated whether changes in gene expression could be translated into changes in protein expression. However, protein expression does not always correlate with mRNA expression for many reasons such as mRNA instability, post-translational protein modifications, mRNA silencing, protein complex formation and different biological or experimental protein and mRNA degradation rates that might affect their correlation (Ali et al., 2013; Ikarashi et al., 2011; Alfonso et al., 2002).
CONCLUSION
This study have partially demonstrated the molecular mechanism underlying the Phytopharmacological action of CoPE compared with the pharmacological action of metformin in vivo, suggesting an enhanced action of the extract and drug in the hyperglycemic state. The hypoglycemic and hypolipidemic effects of both CoPE and metformin have been mediated through PPARγ upregulation in blood and tissues. Of particular important, effects on glucose levels, insulin secretion and sensitivity are insufficient to explain their hypoglycemic action, with most evidence reporting that the major effect of the drug is on glucose utilization. However, to better understand the molecular mechanisms of polyphenols from cocoa along with metformin, hepatic glucose output and the systematic insulin sensitivity in blood should be investigated as well. In addition, molecular investigations using omics technology to determine the global insulin-responsive genes and their transcription factors modulation are necessary. Finally, it is also worth to study the double pharmacological activity of natural compounds and medication drugs on insulin receptor and glucose transporting system in both diabetic individuals and animal models.
ACKNOWLEDGMENT
The authors are grateful for the financial assistance provided by the Fundamental Research Grant Scheme, Ministry of Education, Malaysia (Project No: 02-11-08-611FR) and the laboratory facilities at the Faculty of Medicine and Health Sciences, Universiti Putra Malaysia.
REFERENCES
- Ali, F., Y. Ranneh, A. Ismail and N.M. Esa, 2013. Identification of phenolic compounds in polyphenols-rich extract of Malaysian cocoa powder using the HPLC-UV-ESI-MS/MS and probing their antioxidant properties. J. Food Sci. Technol.
CrossRefDirect Link - Ali, F., A. Ismail and S. Kersten, 2014. Molecular mechanisms underlying the potential antiobesity‐related diseases effect of cocoa polyphenols. Mol. Nutr. Food Res., 58: 33-48.
CrossRefDirect Link - Ali, F., A. Ismail, N.M. Esa and C.P. Pei, 2014. Transcriptomics expression analysis to unveil the molecular mechanisms underlying the cocoa polyphenols treatment in diet-induced obesity rats. Genomics, 105: 23-30.
CrossRefDirect Link - Maleyki, M.A. and A. Ismail, 2010. Antioxidant properties of cocoa powder. J. Food Biochem., 34: 111-128.
CrossRefDirect Link - Alfonso, J., G.D. Pollevick, A. Castensson, E. Jazin and A.C. Frasch, 2002. Analysis of gene expression in the rat hippocampus using real time PCR reveals high inter‐individual variation in mRNA expression levels. J. Neurosci. Res., 67: 225-234.
CrossRefDirect Link - Amin, I., H.A. Faizul and R. Azli, 2004. Effect of cocoa powder extract on plasma glucose levels in hyperglycaemic rats. Nutr. Food Sci., 34: 116-121.
CrossRefDirect Link - Baba, S., N. Osakabe, Y. Kato, M. Natsume and A. Yasuda et al., 2007. Continuous intake of polyphenolic compounds containing cocoa powder reduces LDL oxidative susceptibility and has beneficial effects on plasma HDL-cholesterol concentrations in humans. Am. J. Clin. Nutr., 85: 709-717.
Direct Link - Bailey, C.J. and C. Day, 1989. Traditional plant medicines as treatments for diabetes. Diabetes Care, 12: 553-564.
CrossRefDirect Link - Bisson, J.F., M.A. Guardia-Llorens, S. Hidalgo, P. Rozan and M. Messaoudi, 2008. Protective effect of Acticoa powder, a cocoa polyphenolic extract, on prostate carcinogenesis in Wistar-Unilever rats. Eur. J. Cancer Prev., 17: 54-61.
CrossRefDirect Link - Cha, B.S., T.P. Ciaraldi, L. Carter, S.E. Nikoulina and S. Mudaliar et al., 2001. Peroxisome Proliferator-Activated Receptor (PPAR) γ and Retinoid X Receptor (RXR) agonists have complementary effects on glucose and lipid metabolism in human skeletal muscle. Diabetologia, 44: 444-452.
CrossRefDirect Link - Cooper, K.A., J.L. Donovan, A.L. Waterhouse and G. Williamson, 2008. Cocoa and health: A decade of research. Br. J. Nutr., 99: 1-11.
CrossRefDirect Link - Dillinger, T.L., P. Barriga, S. Escarcega, M. Jimenez, D.S. Lowe and L.E. Grivetti, 2000. Food of the gods: Cure for humanity? A cultural history of the medicinal and ritual use of chocolate. J. Nutr., 130: 2057S-2072S.
Direct Link - Furnsinn, C. and W. Waldhausl, 2002. Thiazolidinediones: Metabolic actions in vitro. Diabetologia, 45: 1211-1223.
CrossRefDirect Link - Grover, J.K., S. Yadav and V. Vats, 2002. Medicinal plants of India with anti-diabetic potential. J. Ethnopharmacol., 81: 81-100.
CrossRefDirect Link - Guo, L. and R. Tabrizchi, 2006. Peroxisome proliferator-activated receptor gamma as a drug target in the pathogenesis of insulin resistance. Pharmacol. Ther., 111: 145-173.
CrossRefDirect Link - Huang, T.H.W., G. Peng, B.P. Kota, G.Q. Li, J. Yamahara, B.D. Roufogalis and Y. Li, 2005. Anti-diabetic action of Punica granatum flower extract: Activation of PPAR-γ and identification of an active component. Toxicol. Applied Pharmacol., 207: 160-169.
CrossRefPubMedDirect Link - Keller, J.M., P. Collet, A. Bianchi, C. Huin and P. Bouillaud-Kremarik et al., 2000. Implications of Peroxisome Proliferator-Activated Receptors (PPARS) in development, cell life status and disease. Int. J. Dev. Biol., 44: 429-442.
PubMed - Kelm, M.A., J.C. Johnson, R.J. Robbins, J.F. Hammerstone and H.H. Schmitz, 2006. High-performance liquid chromatography separation and purification of cacao (Theobroma cacao L.) procyanidins according to degree of polymerization using a diol stationary phase. J. Agric. Food Chem., 54: 1571-1576.
CrossRefDirect Link - Kodera, Y., K.I. Takeyama, A. Murayama, M. Suzawa, Y. Masuhiro and S. Kato, 2000. Ligand type-specific interactions of peroxisome proliferator-activated receptor γ with transcriptional coactivators. J. Biol. Chem., 275: 33201-33204.
CrossRefDirect Link - Kurosawa, T., F. Itoh, A. Nozaki, Y. Nakano and S.I. Katsuda et al., 2005. Suppressive effects of Cacao Liquor Polyphenols (CLP) on LDL oxidation and the development of atherosclerosis in Kurosawa and Kusanagi-hypercholesterolemic rabbits. Atherosclerosis, 179: 237-246.
CrossRefDirect Link - Gershell, L., 2005. Type 2 diabetes market. Nat. Rev. Drug Discovery, 4: 367-368.
CrossRefDirect Link - Bnouham, M., A. Ziyyat, H. Mekhfi, A. Tahri and A. Legssyer, 2006. Medicinal plants with potential antidiabetic activity: A review of ten years of herbal medicine research (1990-2000). Int. J. Diabetes Metabol., 14: 1-25.
Direct Link - Natsume, M., N. Osakabe, M. Yamagishi, T. Takizawa and T. Nakamura et al., 2000. Analyses of polyphenols in cacao liquor, cocoa and chocolate by normal-phase and reversed-phase HPLC. Biosci. Biotechnol. Biochem., 64: 2581-2587.
CrossRefDirect Link - Ikarashi, N., T. Toda, T. Okaniwa, K. Ito, W. Ochiai and K. Sugiyama, 2011. Anti-obesity and anti-diabetic effects of acacia polyphenol in obese diabetic KKAy mice fed high-fat diet. Evidence-Based Complement. Altern. Med.
CrossRefDirect Link - Otto, C., M. Lehrke and B. Goke, 2002. Novel insulin sensitizers: Pharmacogenomic aspects. Pharmacogenomics, 3: 99-116.
CrossRefDirect Link - Pushparaj, P., C.H. Tan and B.K.H. Tan, 2000. Effects of Averrhoa bilimbi leaf extract on blood glucose and lipids in streptozotocin-diabetic rats. J. Ethnopharmacol., 72: 69-76.
CrossRefPubMedDirect Link - Ruzaidi, A., I. Amin, A.G. Nawalyah, M. Hamid and H.A. Faizul, 2005. The effect of Malaysian cocoa extract on glucose levels and lipid profiles in diabetic rats. J. Ethnopharmacol., 98: 55-60.
CrossRefDirect Link - Shirwaikar, A., K. Rajendran, C.D. Kumar and R. Bodla, 2004. Antidiabetic activity of aqueous leaf extract of Annona squamosa in streptozotocin-nicotinamide type 2 diabetic rats. J. Ethnopharmacol., 91: 171-175.
CrossRefPubMedDirect Link - Staels, B. and J.C. Fruchart, 2005. Therapeutic roles of peroxisome proliferator-activated receptor agonists. Diabetes, 54: 2460-2470.
CrossRefDirect Link - Tomura, M.D.D.S., H. Takano, N. Osakabe, M.S.A. Yasuda and I.M.D. Ken-Ichiro et al., 2007. Dietary supplementation with cacao liquor proanthocyanidins prevents elevation of blood glucose levels in diabetic obese mice. Nutrition, 23: 351-355.
CrossRef - Wild, S., G. Roglic, A. Green, R. Sicree and H. King, 2004. Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care, 27: 1047-1053.
CrossRefPubMedDirect Link - Willson, T.M., P.J. Brown, D.D. Sternbach and B.R. Henke, 2000. The PPARs: From orphan receptors to drug discovery. J. Med. Chem., 43: 527-550.
CrossRefDirect Link - Xie, J.T., H.H. Aung, J.A. Wu, A.S. Attele and J. Park, 2002. Effects of American ginseng berry extract on blood glucose levels in ob/ob mice. Am. J. Chin. Med., 30: 187-194.
CrossRefDirect Link - Xie, J.T., A. Wang, S. Mehendale, J. Wu and H.H. Aung et al., 2003. Anti-diabetic effects of Gymnema yunnanense extract. Pharmacol. Res., 47: 323-329.
CrossRefDirect Link - Yin, C.Y., T.S. Ha and K.A. Kadir, 2010. Effects of glycyrrhizic acid on peroxisome Proliferator-Activated Receptor Gamma (PPAR-γ), Lipoprotein Lipase (LPL), serum lipid and HOMA-IR in rats. PPAR Res.
CrossRefDirect Link - Adam, Z.A.I.N.A.H., M. Hamid, A.M.I.N. Ismail, S. Khamis and N. Marsidi, 2010. Antihyperglycemic and glucose tolerance activity of Ficus deltoidea ethanolic extract in diabetic rats. Malaysian J. Health Sci., 8: 25-30.
Direct Link